

Биология внутренних вод, 2022, № 4, стр. 395-403
Вертикальная структура вод и планктон водохранилища в весенний период
А. В. Гончаров a, *, С. Э. Болотов b, c, В. В. Пуклаков a, Д. В. Малашенков a, О. Н. Ерина a, В. А. Ломов a
a Московский государственный университет им. М.В. Ломоносова
Москва, Россия
b Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл., Россия
c Тюменский государственный университет
Тюмень, Россия
* E-mail: mama15333@mail.ru
Поступила в редакцию 24.05.2021
После доработки 30.01.2022
Принята к публикации 11.02.2022
- EDN: QMNECC
- DOI: 10.31857/S0320965222040106
Аннотация
Рассмотрены изменения вертикальной структуры вод и связанная с ней сукцессия планктона Можайского водохранилища в переходный от зимы к лету период 2019 г. Показано, что характерные черты этих изменений остались такими же, как и несколько десятилетий назад. Однако, появились и различия. Происходящее в настоящее время потепление климата приводит в Можайском водохранилище к активизации фотосинтеза подо льдом весной, раннему наступлению весенней гомотермии и более быстрому установлению стратификации. Значительно раньше совершается ежегодный весенний цикл развития циклопов (выходящих из диапаузы); раньше наблюдается следующий за этим максимум дафний. Вместе с тем степень развития зоопланктона весной 2019 г. была меньше, чем 30–40 лет назад. Возможно, причина этого – усиление гипоксии в гиполимнионе. Возрастание гипоксии связано с ранним наступлением и с увеличением продолжительности стратификации водной массы, наблюдаемыми в последние десятилетия, а также – с эвтрофированием водоема.
ВВЕДЕНИЕ
Переходный от зимы к лету период в жизни водоемов насыщен событиями, многие из которых достаточно хорошо изучены. Весной, с прогревом поверхности воды, происходит конвективное перемешивание водной массы и биогенные элементы поднимаются из придонных слоев к поверхности. Возникают благоприятные условия для развития фитопланктона, который в это время представлен мелкими формами, способными к быстрому размножению (Hutchinson, 1967; Sommer et al., 1986; Сахарова, Корнева, 2018).
Затем водную толщу заселяют циклопы: большую часть года они провели на дне в состоянии диапаузы и теперь размножаются, имея достаточное питание для себя и своего потомства (Сахарова, 1970). Позже из покоящихся яиц выводятся дафнии. Они способны к быстрому партеногенетическому размножению и, соответственно, быстрому увеличению численности популяции (Lampert et al., 1986; Elser, Goldman, 1991; Sommer et al., 2007). Дафнии – эффективные фильтраторы, с которыми молодь циклопов не может конкурировать. Поэтому, достигнув IV копеподитной стадии, представители рода Cyclops опускаются на дно водоема, где в малоподвижном состоянии находятся до следующей весны (Naess, Nilssen, 1991).
Изменяется вертикальная структура вод, т.е. характер распределения по глубине водоема основных изучаемых характеристик – устанавливается прямая температурная стратификация водной толщи и формируются эпи-, мета- и гиполимнион.
Дафнии достаточно быстро потребляют фитопланктон, прозрачность воды увеличивается, наступает фаза “чистой воды” (Lampert, Sommer, 2007). Численность дафний, подорвавших свою кормовую базу, через некоторое время также резко сокращается.
Сходный характер развития планктона отмечен и в Можайском водохранилище, служащем объектом нашего рассмотрения. Проводившиеся здесь в 1981–1991 гг. исследования выявили характерную последовательность изменений фито- и зоопланктона (Сахарова, Левшина, 1992; Сахарова и др., 1993). Однако связь этих изменений с перестройками вертикальной структуры водоема не была изучена. С тех пор прошло >30 лет, в течение которых в Можайском водохранилище произошли изменения, связанные с потеплением климата (Пуклаков и др., 2019).
Цель работы – изучить особенности планктона в период весенней трансформации водной массы, а также оценить возможные изменения в водной экосистеме при потеплении климата.
Актуальность таких исследований подтверждается многими современными работами, в которых показано, что потепление климата приводит к изменению вертикальной структуры водоемов (Richardson, 2008; Jane et al., 2021), что, в свою очередь оказывает воздействие на планктон (Carter et al., 2017; Razlutskij et al., 2018; Сухаревич, Поляк, 2020).
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследования проводили в средней части Можайского водохранилищa у дер. Красновидово в районе затопленного русла р. Москвы (55°34.958′ с.ш., 35°51.536′ в.д.). Можайское водохранилище – типичный для средней полосы России водоем долинного типа, созданный в 1960 г. в верхнем течении р. Москвы и осуществляющий многолетнее регулирование ее стока. Объем водохранилища при НПУ 0.23 км3, площадь водной поверхности 30.7 км2, длина 28 км, ширина ≤2.6 км, максимальная глубина до 22 м у плотины гидроузла. Водоем слабопроточный (коэффициент водообмена ~1.8 год–1), стратифицирован летом и зимой, относится к эвтрофному типу (Пуклаков и др., 2015).
Наблюдения проводили с 10 марта 2019 г., когда водоем еще был покрыт льдом, до 17 июня 2019 г., когда в нем установилась вертикальная стратификация вод. Глубина в точке исследования была ~12 м.
Исследование зоопланктона, фитопланктона и фосфора проводили с 21 апреля 1 раз в 7–10 сут. Фосфаты определяли методом Морфи–Райли в соответствии с РД 52.10.738-2010 (Массовая концентрация фосфатов в морских водах. Методика измерений фотометрическим методом); пробы отбирали с горизонтов 0.5 и 11 м.
Пробы зоопланктона отбирали с помощью замыкающей сети Джеди (размер ячеи 82 мкм), диаметр входного отверстия 20 см. В период с 21 апреля по 26 мая зоопланктон облавливали в слоях воды 0–4 и 4–11 м; позже, с усилением вертикальной стратификации вод, пробы отбирали в слоях 0–4, 4–7, 7–11 м, которые примерно соответствовали эпи-, мета- и гиполимниону соответственно. Пробы фиксировали 4%-ным формалином, подсчет и определение организмов проводили под микроскопом в камере Богорова, биомассу рассчитывали по средним размерам организмов каждого вида (Методика…, 1975; Балушкина, Винберг, 1979). Для идентификации зоопланктона использовали определители (Кутикова, 1970; Определитель…, 2010).
Фитопланктон отбирали батометром Руттнера из поверхностного и придонного горизонтов (0.5 и 11 м), с усилением стратификации (после 26 мая) добавляли промежуточный горизонт (6 м). Пробы фиксировали формалином, концентрировали фильтрованием через мембранные фильтры (фирма “Владипор”) с диаметром пор 0.8 мкм. Водоросли подсчитывали в камере Нажотта, биомассу определяли по средним размерам клеток каждого вида в пробе (Методика…, 1975; Кузьмин, 1984).
С помощью зонда YSI 6600-2V определяли распределение температуры воды и содержания растворенного кислорода по глубине водоема. С 10 марта по 25 апреля водную толщу зондировали вручную (через 1 м), с 4 мая – с помощью автоматической лебедки, установленной на плоту в точке исследования. Опускание прибора до дна и подъем к поверхности проводили круглосуточно каждые 3 ч. Параметры регистрировали 1 раз в минуту, что примерно соответствовало прохождению зондом одного метра. Имелось несколько перерывов в работе лебедки, связанные с техническими проблемами. Настройку (калибровку) зонда проводили в соответствии с инструкцией (https://www.ysi.com).
Сведения о температуре воздуха и скорости ветрa получены с метеостанции г. Можайска, расположенной в 12 км от пункта наблюдения (https://rp5.ru), данные по уровню воды – из АО “Мосводоканал”. Многолетние данные о температуре воды в Можайском водохранилище у дер. Красновидово (на глубине 0.5 м) получены на гидрологическом посту Красновидовской учебно-научной базы Географического факультета МГУ им. М.В. Ломоносова. Использованы также данные о зоопланктоне за 1981–1991 гг., предоставленные М.И. Сахаровой.
Расчеты и построение диаграмм проводили с использованием статистических программ. Связи между изучаемыми параметрами выявляли с помощью коэффициента корреляции Пирсона, достоверность корреляции оценивали с помощью t-критерия Стьюдента (Христофоров, Юмина, 2017).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Вертикальная структура вод. В конце зимнего периода (10 марта) при толщине льда 47 см (лед без снега, малопрозрачный) в водохранилище еще сохранялась обратная температурная стратификация (рис. 1): на глубине 0.5 м температура была 1.2°С, на 6 м – 2.4°С, на 10.5 м – 1.9°С (понижение температуры у дна связано с проникновением по русловой ложбине более холодных вод р. Москвы) (Пуклаков и др., 2015). Содержание кислорода у поверхности достигало 73% насыщения, у дна – ~40%.
Рис. 1.
Вертикальное распределение температуры воды (а) и растворенного кислорода (% насыщения) (б) в Можайском водохранилище в марте–июне 2019 г.

С повышением температуры воздуха лед становился тоньше (3 апреля – 38 см), его верхний малопрозрачный водно-снеговой слой разрушался. Проникающие в воду солнечные лучи нагревали ее и активизировали фотосинтез фитопланктона. К 3 апреля температура воды на глубине 1 м увеличилась до 3.3°С, содержание растворенного кислорода – до 122% насыщения (рис. 1). Столь высокое значение обусловлено тем, что лед препятствует выходу в атмосферу кислорода, выделяющегося при фотосинтезе (Ерина и др., 2019).
К 21 апреля водоем полностью освободился ото льда. Происходило конвективное перемешивание водной массы, при котором содержание кислорода в придонных слоях увеличилось до 60%. Температура воды у поверхности достигла 5.3°С, к 25 апреля – 9–10°С. Начала устанавливаться прямая температурная стратификация, которая сохранялась в течение всего последующего периода наблюдений (рис. 1).
На глубинах 4–8 м формировался металимнион, градиент температуры в нем постепенно увеличивался с 1°С/м в середине мая до 2°С/м в середине июня. Это привело к усилению стратификации и ослаблению вертикального перемешивания.
Для рассматриваемого периода характерно пересыщение поверхностного слоя воды кислородом и постепенное истощение его в придонных слоях – к 10 июня глубже 8 м сформировалась устойчивая зона с содержанием кислорода <1 мг/л (<10% насыщения) (рис. 1). Ее стабильное существование определялось вертикальной стратификацией, препятствующей поступлению кислорода из поверхностных слоев воды на глубину.
Установление стратификации тесно связано с гидрометеорологическими условиями (Эдельштейн, 2014). Это подтверждают коэффициенты корреляции (табл. 1): вертикальные градиенты температуры и кислорода (разность между их значениями у поверхности и у дна) закономерно возрастают с увеличением температуры воды, температуры воздуха, интенсивности солнечной радиации и снижаются при усилении ветра.
Таблица 1.
Коэффициенты корреляции (Пирсона) между параметрами, характеризующими вертикальную стратификацию водной толщи Можайского водохранилища, и метеорологическими данными в период 4 мая–10 июня 2019 г.
| Показатель | Тводы | Градиент О2 | Градиент Т |
|---|---|---|---|
| Твоздуха | 0.84 | 0.85 | 0.85 |
| Тводы | 1.00 | 0.98 | 0.99 |
| Градиент О2 | 0.98 | 1.00 | 0.98 |
| Градиент Т | 0.99 | 0.98 | 1.00 |
| P | 0.70 | 0.74 | 0.66 |
| V | –0.63 | –0.60 | –0.60 |
| ФАР | 0.69 | 0.63 | 0.70 |
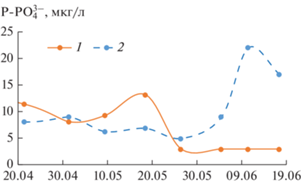
Вертикальная структура вод отражается и на распределении фосфора. Во время перемешивания водной массы концентрации фосфатов у поверхности и у дна различаются не сильно – 6–13 мкг Р-Р${\text{O}}_{4}^{{3 - }}$/л. После установления вертикальной стратификации количество фосфатов, потребляемых фитопланктоном, в поверхностном горизонте снижается до 2.5 мкг Р-Р${\text{O}}_{4}^{{3 - }}$/л, а у дна возрастает до 17–23 мкг Р-Р${\text{O}}_{4}^{{3 - }}$/л (рис. 2). Это связано с тем, что у дна, в условиях наступившей гипоксии, происходит активное выделение фосфатов из иловых отложений (Мартынова, 1984), однако в условиях стратификации подъема их к поверхности не происходит.
Рис. 2.
Изменение концентрации фосфатов у поверхности (1) и в придонных слоях (2) воды Красновидовского плеса Можайского водохранилища в 2019 г.
Зоопланктон, фитопланктон. До 9 мая, когда температура воды достигала 5–13оС, степень развития зоопланктона была невелика – 0.3–0.6 г/м3 (рис. 3а). От поверхности до дна зоопланктон распределялся почти равномерно и представлен, главным образом, веслоногим рачком Cyclops strenuus Fischer, 1851 (80% биомассы зоопланктона) (рис. 3б).
Рис. 3.
Изменение биомассы зоопланктона (а), его основных представителей (Cyclops strenuus и Daphnia) (б) и фитопланктона (в) в Можайском водохранилище в 2019 г. 1 – Rotifera, 2 – Cladocera, 3 – Copepoda, 4 – Cyclops strenuus в слое 4–11 м, 5 – Cyclops strenuus в слое 0–4 м, 6 – Daphnia, 7 – диатомовые водоросли, 8 – зеленые, 9 – прочие.

С 21 апреля по 18 мая наблюдали слабое развитие фитопланктона (0.2–0.9 мг/л) с преобладанием по биомассе (>90%) диатомовых водорослей (в основном, Synedra sp.). От поверхности до дна водоросли были распределены почти равномерно (рис. 3в). К 18 мая вода у поверхности прогревалась до 16°С, количество Cyclops strenuus существенно возросло (рис. 3а, 3б) – >90% общей биомассы Copepoda (биомассу Cyclops strenuus рассчитывали без учета науплиальных и младших копеподитных стадий). Увеличилась численность коловратки Conochilus unicornis Rousselet, 1892 (до 200 тыс. экз./м3), служащей пищей для взрослых циклопов и их молоди (Монаков, 1998). В это время начинают появляться дафнии (Daphnia galeata Sars, 1864).
В конце мая–начале июня вода прогрелась до 20–25°С. При такой температуре дафнии стали половозрелыми, у них быстро созрели яйца и появилась молодь. В это же время возросла кормовая база фильтраторов – биомасса фитопланктона, представленного, в основном, диатомовыми Cyclostephanos dubius (Fricke) Round (>90% общей биомассы фитопланктона), увелчилавсь до 3.6 и 2.7 мг/л у поверхности 26 мая и 3 июня соответственно (рис. 3в). Это способствовало увеличению плодовитости дафний (Сахарова, Левшина, 1992). При таких благоприятных условиях численность фильтраторов у поверхности водоема резко возросла и достигла 3 июня 100 тыс. экз./м3 (при биомассе ~2.3 г/м3). Преобладали Daphnia cucullata Sars, 1862 (49% биомассы ветвистоусых), Daphnia cristata Sars, 1862 (30%) и Daphnia galeata (13%). В то же время количество Cyclops strenuus резко снизилось (рис. 3б).
К 10 июня биомасса фитопланктона, выедаемого дафниями, в поверхностном горизонте уменьшилась до 0.64 мг/л, а к 17 июня уменьшилась и биомасса зоопланктона – до 0.8 г/м3 у поверхности (рис. 3).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
По данным проведенного исследования можно выделить три периода в развитии планктона Можайского водохранилища. В первый период, с 21 апреля до 18 мая, при температуре воды 5–16°С, наблюдали слабое развитие организмов с преобладанием по биомассе диатомовых водорослей Synedra sp. и веслоногих раков Cyclops strenuus. По данным (Сахарова, 1973; Naess, Nilssen, 1991), последние переживают зиму на дне в состоянии диапаузы и ранней весной поднимаются к поверхности для размножения. Максимальная биомасса циклопов отмечена в конце первого периода – 1.8 г/м3 у поверхности.
Во второй период (26 мая–10 июня) произошли существенные изменения в планктоне: Synedra sp. заместилась на Cyclostephanos dubius с резким увеличением биомассы фитопланктона, а веслоногие ракообразные – уступили место ветвистоусым (дафниям).
Дафнии выводятся из покоящихся яиц – эфипиумов, проводящих зиму на дне водоема. Нарождающейся молодью питаются циклопы (Монаков, 1998). Возможно, поэтому высокой биомассы дафнии достигают после 18 мая, когда количество Cyclops strenuus существенно уменьшается (рис. 3а, 3б). По данным (Lampert, 1987), в Боденском озере дафнии достигают максимальной численности весной только после того, как циклопы перейдут в диапаузу. Аналогичная ситуация наблюдалась в Можайском водохранилище: с 18 мая по 10 июня концентрация циклопов у поверхности снижалась быстрее по сравнению с нижележащими слоями, отражая процесс опускания циклопов на дно (рис. 3б).
Cyclops strenuus в процессе эволюции выработал способность к диапаузе, из которой выходит ранней весной, когда хищников мало и имеется достаточное количество пищи. В это время он быстро производит потомство, которое, подрастая, снова впадает в диапаузу на дне и таким образом избегает выедания. Cyclops strenuus можно назвать видом-эксплерентом, который в условиях резких колебаний ресурса, максимально использует его в период высоких значений и уходит в диапаузу до следующего подъема ресурса (Романовский, 1989; Сахарова, Левшина, 1992).
Сходное с изученным в 2019 г. поведение циклопов описано по наблюдениям, проведенным на Можайском водохранилище в 1981–1984 гг. (Сахарова, Левшина, 1992). Различие состоит в видовом составе: преобладающим видом в те годы был Cyclops vicinus Uljanin, 1875, достигавший ~60% биомассы зоопланктона в апреле–мае), в настоящее время – это Cyclops strenuus (~90%). Проводившиеся ранее исследования показали также, что опустившиеся на дно циклопы в огромном количестве скапливаются в русловой ложбине водохранилища, где их численность зимой была несколько миллионов экз./м2, биомасса – до 100 г/м2 (Сахарова, 1978).
Видовой состав дафний в 2019 г. сходен с наблюдавшимся в начале летних периодов 1981–1991 гг. (Сахарова, Левшина, 1992). Различие заключается в том, что прежде доминантом была Daphnia galeata (~90% биомассы зоопланктона в июне 1981–1991 гг.), а в 2019 г. преобладала Daphnia cucullata (~60%).
Исследование 2019 г. показало, что начиная с 18 мая биомасса дафний с глубиной снижается, несмотря на наличие в средних слоях водоема в начале июня наибольшего количества пищи – диатомовых водорослей (рис. 3а, 3в). Причиной уменьшения биомассы дафний может быть снижение с глубиной содержания кислорода, концентрация которого в придонных слоях в начале июня падает до 1 мг/л, в металимнионе – до 4–5 мг/л (рис. 1). К сходному выводу пришли исследователи трех стратифицированных озер в Польше (Razlutskij et al., 2018): уменьшение содержания растворенного кислорода ниже эпилимниона препятствовало вертикальной миграции зоопланктона, и он скапливался в эпилимнионе.
Обращает на себя внимание необычное распределение фитопланктона по глубине в первой половине июня: больше всего водорослей (Cyclostephanos dubius) сосредоточено не у поверхности, а в средних слоях водоема (до 8 мг/л на глубине 6 м 3 июня). Причиной этого может быть выедание водорослей дафниями (Sommer, 1989), которых больше всего в верхнем горизонте. Кроме того, клетки Cyclostephanos dubius погружаются вниз, поскольку имеют массивные кремнеземные створки (Reynolds, 1990). Их опусканию способствует установление в начале июня вертикальной стратификации водной массы (рис. 1), при которой перемешивание воды почти отсутствует.
Третий период (после 10 июня) характеризуется уменьшением биомассы и фитопланктона, и зоопланктона: дафнии подрывают свою кормовую базу; кроме того, сами они становятся жертвой хищных беспозвоночных и мальков рыб, которые обычно появляются в водоеме в это время. Развитие фитопланктона сдерживается также тем, что в условиях стратификации водной толщи, биогенные элементы не поступают из придонных слоев к поверхности в зону фотосинтеза.
Многолетние изменения. Данные метеостанции г. Можайск показывают, что средняя годовая температура воздуха в рассматриваемом районе устойчиво повышалась с 1987 г. по 2018 г., достигая 0.17°С/10 лет. Причина – глобальное потепление климата, которое в данном регионе проявилось особенно ярко (Курганова и др., 2017).
Многолетние исследования Можайского водохранилища (Пуклаков и др., 2019) показали, что в настоящее время наступление летней вертикальной стратификации водной толщи происходит на 9 сут раньше, чем в 80-е годы, а весенней гомотермии – на 5 сут. При этом средняя длительность летней стратификации увеличилась на 18 сут, длительность весенней гомотермии сократилась на 4 сут. Поэтому можно ожидать, что в настоящее время раньше наступают не только гидрометеорологические процессы, но и биологические, и гидрохимические, которые с ними связаны.
Максимумы развития циклопов и дафний в 2019 г. наблюдали значительно раньше, чем в прежние годы (в среднем за 1981–1984, 1987, 1990–1991 гг.), разница была около полумесяца (рис. 4). В то же время, по абсолютным значениям биомассы 2019 г. существенно уступает средним значениям 30–40-летней давности, несмотря на больший прогрев воды в настоящее время.
Рис. 4.
Сравнение биомассы зоопланктона в 2019 г. с таковой в тот же период 30–40-летней давности. 1 – Cladocera среднее многолетнее, 2 – Cladocera, 2019 г., 3 – Cyclopoida, среднее многолетнее, 4 – Cyclopoida, 2019 г.

Смещение сроков развития зоопланктона демонстрирует также связь между временем наступления максимумов численности рачков и средней температурой воды у поверхности (0.5 м) за предшествующий месяц (коэффициенты корреляции ~ –0.9) (рис. 5). Для Cladocera предшествующим месяцем считали весь май, для Cyclopoida – период с 15 апреля по 14 мая, т.е., чем теплее весна, тем раньше достигается максимум развития зоопланктона.
Рис. 5.
Связь между временем наступления максимума численности рачков и средней температурой воды у поверхности (0.5 м) за предшествующий месяц: а – для Cyclopoida, б – для Cladocera.
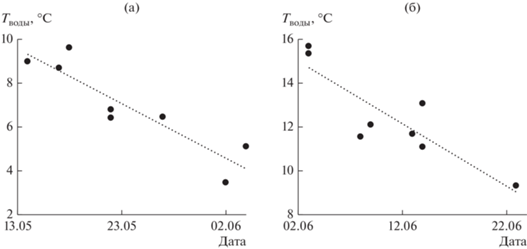
Что касается влияния температуры на количественное развитие зоопланктона весной, то для прошлых лет (1981–1984, 1987, 1990–1991 гг.) оно не обнаруживается: коэффициенты корреляции между максимумами Cyclopoida и Cladocera и соответствующими весенними температурами воды близки к нулю.
Низкие показатели количественного развития зоопланктона в 2019 г., по сравнению с прежними годами, предположительно можно объяснить ухудшением кислородного режима в водоеме в течение последнего 30–40-летнего периода. Как показано выше, потепление климата приводит к раннему установлению вертикальной стратификации вод, что влечет за собой формирование более продолжительного периода гипоксии в нижних слоях водоема летом (Даценко, Пуклаков, 2021). Кроме того, происходит эвтрофирование водохранилища вследствие постоянного поступления в него органических веществ и биогенных элементов с речным стоком (Даценко, Пуклаков, 2019). Содержание кислорода в гиполимнионе особенно важно для циклопов, проводящих значительную часть жизни на дне в состоянии диапаузы. Снижение концентрации растворенного кислорода при потеплении климата подтверждается результатами исследования многих озер умеренной зоны. Так, Джейн с соавторами (Jane et al., 2021), обобщив данные о 393 озерах за 1941–2017 гг., показала, что уменьшение кислорода происходит не просто в результате снижения растворимости газа при повышенной температуре, но и в результате усиления вертикальной стратификации, препятствующей аэрации нижележащих слоев водоема.
Выводы. Общий ход сукцессии планктона весной 2019 г. был сходен с наблюдаемым несколько десятилетий назад. Вместе с тем, отмечены различия. Изменился состав преобладающих видов зоопланктона. Если раньше доминировала Daphnia galeata, то в 2019 г. ей на смену пришла Daphnia cucullata, вместо Cyclops vicinus появился Cyclops strenuus. Максимум развития этих видов в 2019 г. наблюдался намного раньше, чем в прежние годы из-за более раннего весеннего прогрева воды. Однако уровень количественного развития зоопланктона снизился. Возможно, причина этого – ухудшение кислородных условий в водоеме, вызванное ранним наступлением стратификации в результате потепления климата, а также эвтрофированием водохранилища.
Список литературы
Балушкина Е.Б., Винберг Г.Г. 1979. Зависимость между массой и длиной тела у планктонных животных. Общие основы изучения водных экосистем. Ленинград: Наука. С. 169.
Даценко Ю.С., Пуклаков В.В. 2019. Анализ многолетних изменений экологического состояния Можайского водохранилища по расчетам его кислородного режима. Водохранилища Российской Федерации: Сб. матер. Всерос. науч.-практ. конф. Новочеркасск: Лик. С. 457.
Даценко Ю.С., Пуклаков В.В. 2021. Анализ многолетних изменений зон аноксии в Можайском водохранилище по результатам модельных расчетов // Водные ресурсы. Т. 48. № 1. С. 34. https://doi.org/10.31857/S0321059621010132
Ерина О.Н., Пуклаков В.В., Соколов Д.И., Гончаров А.В. 2019. Подледное цветение фитопланктона в Можайском водохранилище // Вестн. биотех. и физ.-хим. биологии им. Ю.А. Овчинникова. Т. 15. № 2. С. 49.
Кузьмин Г.В. 1984. Таблицы для вычисления биомассы водорослей. Магадан: Дальневосточный науч. центр АН СССР.
Курганова И.Н., Лопес де Гереню В.О., Аблеева В.А., Быховец С.С. 2017. Климат южного Подмосковья: Современные тренды и оценка аномальности. Фундаментальная и прикладная метеорология. Т. 4. С. 66.
Кутикова Л.А. 1970. Коловратки фауны СССР. Ленинград: Наука.
Мартынова М.В. 1984. Азот и фосфор в донных отложениях озер и водохранилищ. Москва: Наука.
Методика изучения биогеоценозов внутренних водоемов. 1975. Москва: Наука.
Монаков А.В. 1998. Питание пресноводных беспозвоночных. Москва: Ин-т проблем эвол. и экол. РАН им. А.Н. Северцова.
Определитель зоопланктона и зообентоса пресных вод Европейской России. 2010. Т. 1. Зоопланктон. Москва: Тов-во науч. изд. КМК.
Пуклаков В.В., Даценко Ю.С., Гончаров А.В. и др. 2015. Гидроэкологический режим водохранилищ Подмосковья. Москва: Перо.
Пуклаков В.В., Ерина О.Н., Соколов Д.И., Эдельштейн К.К. 2019. Влияние современных климатических изменений на гидрологический режим Можайского водохранилища. Глобальные климатические изменения: региональные эффекты, модели, прогнозы. Междунар. науч.-практ. конф. Воронеж. С. 484.
Романовский Ю.Э. 1989. Конкуренция за флуктуирующий ресурс: эволюционные и экологические последствия // Журн. общ. биол. Т. 50. № 3. С. 304.
Сахарова Е.Г., Корнева Л.Г. 2018. Фитопланктон литорали и пелагиали Рыбинского водохранилища в годы с разными температурным и уровенным режимами // Биология внутр. вод. № 1. С. 11. https://doi.org/10.7868/S0320965218010023
Сахарова М.И. 1970. Сезонная динамика микробентоса Учинского водохранилища // Зоол. журн. Т. 49. № 12. С. 1767.
Сахарова М.И. 1973. Донная фаза в жизненном цикле массовых видов пелагических циклопов Учинского водохранилища // Комплексные исследования водохранилищ. № 2. Москва: Изд-во МГУ. С. 122.
Сахарова М.И. 1978. Микробентос Можайского водохранилища. Комплексные исследования водохранилищ. № 4. Москва: Изд-во МГУ. С. 192.
Сахарова М.И., Левшина Н.А. 1992. Многолетние исследования сезонной динамики планктонного сообщества в пелагиали Можайского водохранилища // Экологические исследования в г. Москве и Московской области. Состояние водных систем. Москва: ИНИОН РАН. С. 145.
Сахарова М.И., Соколова Н.Ю., Эдельштейн К.К. 1993. Многолетние изменение структуры сообществ и некоторые мероприятия по снижению темпов евтрофирования // Биол. науки. № 1. С. 66.
Сухаревич В.И., Поляк Ю.М. 2020. Глобальное распространение цианобактерий: причины и последствия (обзор) // Биология внутр. вод. № 6. С. 562.https://doi.org/10.31857/S0320965220060170
Христофоров А.В., Юмина Н.М. 2017. Теория вероятностей и математическая статистика. Учебное пособие. Московский гос. ун-т им. М.В. Ломоносова, Геогр. ф-т. Москва: АПР.
Эдельштейн К.К. 2014. Гидрология озер и водохранилищ. Учебник для вузов. Москва: “Перо”.
Carter J., Schindler D., Francis T. 2017. Effects of climate change on zooplankton community interactions in an Alaskan lake // Clim Chang Responses. V. 4. № 3. https://doi.org/10.1186/s40665-017-0031-x
Elser J.J., Goldman C.R. 1991. Zooplankton effects on phytoplankton in lakes of contrasting trophic status // Limnol., Oceanogr. V. 36. № 1. P. 64. https://doi.org/10.4319/lo.1991.36.1.0064
Hutchinson G.E. 1967. Introduction to Lake Biology and Limnoplankton. A Treatise on Limnology // BioScience. V. 2. New York: John & Sons.
Jane S.F., Hansen Gretchen J.A., Kraemer B.M. et al. 2021. Widespread deoxygenation of temperate lakes // Nature. V. 594. P. 66. https://doi.org/10.1038/s41586-021-03550-y
Lampert W. 1987. Predictability in lake ecosystems: the role of biotic interactions // Ecological studies. V. 61. P. 333. https://doi.org/10.1007/978-3-642-71630-0_16
Lampert W., Sommer U. 2007. Limnoecology. New York: Oxford University Press Inc.
Lampert W., Fleckner W., Rai H., Taylor B.E. 1986. Phytoplankton control by grazing zooplankton: A study on the spring clear-water phase // Limnol., Oceanogr. V. 31. № 3. P. 478. https://doi.org/10.4319/lo.1986.31.3.0478
Naess T., Nilssen J.P. 1991. Diapausing fertilized adults // Oecologia. V. 86. P. 368. https://doi.org/10.1007/BF00317602
Razlutskij V., Feniova I., Ejsmont-Karabin J. et al. 2018. Impact of enhanced summer temperatures on the distribution and structure of zooplankton communities in the heated stratified lakes: Implications for climate change // Limnologica. V. 73. P. 1. https://doi.org/10.1016/j.limno.2018.08.004
Reynolds C.S. 1990. Temporal scales of variability in pelagic environments and response of phytoplankton // Freshwater Biol. V. 23. P. 25. https://doi.org/10.1111/j.1365-2427.1990.tb00252.x
Richardson A. 2008. In hot water: zooplankton and climate change // ICES Journal of Marine Science. V. 65. P. 279. https://doi.org/10.1093/icesjms/fsn028
Sommer U. 1989. Plankton ecology. Succession in plankton communities. Berlin; New York: Springer.
Sommer U., Gliwicz Z.M., Lampert W., Duncan A. 1986. The PEG-model of seasonal succession of planktonic events in fresh waters // Arch. Hydrobiol. V. 106. P. 433.
Sommer U., Aberle N., Engel A. et al. 2007. Anindoor mesocosm system to study the effect of climate change on the late winter and spring succession of Baltic Sea phyto- and zooplankton // Oecologia. V. 150. P. 655. https://doi.org/10.1007/s00442-006-0539-4
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод