

Биология внутренних вод, 2022, № 4, стр. 381-394
Оценка состояния растительного покрова трансграничного водотока северной Фенноскандии (российская часть р. Паз)
Н. В. Зуева a, *, О. Г. Гришуткин b, Ю. А. Зуев c, Д. Ю. Ефимов b, Е. В. Чемерис b, А. А. Бобров b, **
a Российский государственный гидрометеорологический университет
Санкт-Петербург, Россия
b Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл, Россия
c Санкт-Петербургский филиал ВНИРО, ГосНИОРХ им. Л.С. Берга
Санкт-Петербург, Россия
* E-mail: nady.zuyeva@ya.ru
** E-mail: bobrov@ibiw.ru
Поступила в редакцию 30.09.2021
После доработки 19.01.2022
Принята к публикации 29.01.2022
- EDN: NQRCBQ
- DOI: 10.31857/S0320965222040325
Аннотация
По материалам 2019 г. дана оценка растительного покрова пограничной р. Паз. Обнаружено высокое разнообразие флоры, нехарактерное для других рек северной Фенноскандии. Основу растительного покрова в русле реки составляют Sparganium angustifolium, Myriophyllum alterniflorum, Ranunculus schmalhausenii, Potamogeton gramineus, P. perfoliatus, на мелководьях – Subularia aquatica, Isoёtes echinospora, I. lacustris, вдоль берега – Equisetum fluviatile, Carex rostrata, Comarum palustre – преимущественно виды олиго- и мезотрофных вод. Состав макрофитов почти не изменился за >25 лет с момента первого исследования. Зарастание реки по сравнению с водотоками региона повышенное. Наибольший вклад в продукцию органического вещества вносят доминирующие в русле реки Potamogeton perfoliatus и P. gramineus. Основная причина этого – благоприятный для развития растений гидрологический режим, формируемый на реке каскадом гидроэлектростанций. Состояние исследованного участка водотока по характеристикам растительного покрова признано стабильным и благополучным.
ВВЕДЕНИЕ
Водные экосистемы высоких широт затронуты глобальными антропогенными и климатическими изменениями (Prowse et al., 2006; Wrona et al., 2006; AMAP, 2017). Это определяет угрозы и для пресноводных объектов, распространенных в Арктике и Субарктике. Особое внимание привлечено к проблеме изменения биологического разнообразия (Wrona et al., 2006; Heino et al., 2009; Culp et al., 2012). Так, в северной Фенноскандии расположено значительное число рек, в разнообразии биоты которых могут произойти изменения (Heino et al., 2003, 2009; Tolonen et al., 2016; Mustonen et al., 2018; Brittain et al., 2020).
Река Паз (Пасвик, Патсойоки, Патсо-йоки) – трансграничная река cеверной Фенноскандии, протекающая по территории Финляндии, России и Норвегии. Ее бассейн находится севернее Полярного круга. Это субарктическая система, характеризующаяся значительным разнообразием гидробионтов (Ylikörkkö et al., 2015).
В настоящее время экосистема реки значительно антропогенно трансформирована. (Moiseenko et al., 1994). Она находится под влиянием измененного гидрологического режима, промышленного загрязнения, биологических инвазий (Amundsen et al., 2019) и климатических флуктуаций (Демин, 2012; Ylikörkkö et al., 2015). Оценка состояния водной растительности в таких условиях весьма актуальна.
Флористические исследования в бассейне р. Паз были начаты в конце XIX–начале XX в. Их история подробно описана в работах (Кожин и др., 2020; Кравченко, 2020). Сведения о водной флоре и растительности р. Паз содержатся в ряде общефлористических и специализированных работ (Летопись…, 1998; Костина, 2003; Канева, 2008; Канева, Ламзов, 2009; Кириллова, 2012, 2013; Кравченко и др., 2017; Боровичев, Бойчук, 2018; Кравченко, 2020). Определение экологического состояния озерно-речной системы р. Паз на основе анализа разнообразия водных макрофитов, индикаторных свойств видов, их обилия и частоты встречаемости проведено в двух исследования (Moiseenko et al., 1994; Riihimäki el al., 2013). В них отмечен хороший и высокий экологический статус водотока, несмотря на его антропогенное преобразование. Однако оценку проводили на основе материалов, полученных преимущественно на участках водотока, расположенных в Финляндии и Норвегии.
Попыток комплексного изучения водной флоры и растительности (видовой состав, разнообразие фитоценозов, степень и характер зарастания, продукция) для оценки состояния озерно-речной системы р. Паз до сих пор не предпринимали.
Цель работы – оценить состояние растительного покрова р. Паз.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Территория исследования. Длина р. Паз 147 км, водосборная площадь 18 325 км2, средний многолетний расход воды (Борисоглебская ГЭС) 186.8 м3/с (Водный…, 2019; Качество…, 2020). Исток – оз. Инари и первые 2 км реки находятся в Финляндии. Далее до пос. Раякоски р. Паз протекает по территории России, а ниже по руслу реки проходит государственная граница с Норвегией. В нижнем течении в районе пос. Борисоглебский небольшая часть водотока вновь находится в России, а устьевая зона, где река впадает в Варангер-фьорд Баренцева моря, – в Норвегии (рис. 1). Водоток представляет собой озерно-речную систему, зарегулированную семью гидроэлектростанциями.
Рис. 1.
Изученный участок р. Паз и расположение исследованных станций. 1 – станции исследования, границы участков реки: 2 – гидроэлектростанции, 3 – естественные сужения, 4 – государственная граница.

Данный район подвергается заметному антропогенному воздействию. Во-первых, это влияние комбината Печенганикель – выбросы тяжелых металлов и диоксида серы. Кроме того, в нижнем течении река принимает промышленные и коммунальные стоки, а на части водосборной территории ведется лесное и сельское хозяйство (Moiseenko et al., 1994; Сандимиров, 2012).
Участок р. Паз, длиной 98 км, исследован в июне–сентябре 2019 г., он начинается от предплотинной зоны водохранилища ГЭС Кайтакоски и простирается вниз по течению до оз. Сальмиярви (рис. 1). Здесь расположено шесть ГЭС (четыре российских и две норвежских): Кайтакоски, Янискоски, Раякоски, Хевоскоски, Скугфосс, Мелькефосс. Участок от ст. р11 до ст. р26 входит в государственный заповедник “Пасвик”, ст. p27 расположена на его границе.
Плотинами и естественными сужениями исследованная часть реки делится на девять участков, их характеристика дана в табл. 1. Описание растительного покрова восьми из них проведено на 27 станциях, на каждом участке располагалось от одной до шести станций (табл. 1).
Таблица 1.
Некоторые характеристики участков реки
| Участок реки (станции) | S, км2 | SРФ, км2 (%) | W, м | L, км | TDS, мг/л |
|---|---|---|---|---|---|
| Виртаниеми–Кайтакоски (p1) | 1.8 | 1.8 (100) | 305 | 6.0 | 15–16 |
| Кайтакоски–Янискоски (p2–p5) | 4.4 | 4.4 (100) | 528 | 8.4 | 14 |
| Янискоски–Раякоски (p6–p10) | 7.2 | 7.2 (100) | 646 | 11.1 | 14–18 |
| Раякоски–Хевоскоски (–) | 14.5 | 7.0 (48) | 883 | 16.4 | 16 |
| Хевоскоски–Йорданфосс (p11–p13) | 9.0 | 5.1 (56) | 780 | 11.6 | 15–18 |
| Йорданфосс–проход в Лангватнет (p14–p16) | 34.8 | 8.0 (23) | 1417 | 13.4 | 15 |
| Проход в Лангватнет–Скугфосс (p17–p22) | 23.9 | 12.8 (54) | 1352 | 17.7 | 15 |
| Скугфосс–Мелькефосс (p23) | 2.0 | 0.8 (42) | 332 | 5.1 | 16–17 |
| Мелькефосс–Сальмиярви (p24–p27) | 16.7 | 8.3 (50) | 1131 | 14.8 | 17–35 |
В исследованиях использованы традиционные методические подходы (Катанская, 1981; Бобров, Чемерис, 2006). Описание растительного покрова на станциях проводили с лодки и берега, протяженность станций ~500 м, небольшие заливы осматривали полностью. Портативным анализатором (Hanna HI 98129) измеряли минерализацию воды. В доминирующих растительных сообществах проводили укосы с площади 0.25 м2 для дальнейшего определения фитомассы и продуктивности. Видовое разнообразие оценивали с использованием индекса Шеннона (Shannon, 1948).
Степень зарастания всего русла оценивали с помощью анализа спутниковых снимков за 2017 г. в программе MapInfo 11.5. Использованные снимки находятся в свободном доступе интернет-сервисов Яндекс (https://yandex.ru/maps) и Bing (https://www. bing.com/maps).
Валовая годовая продукция растительных сообществ рассчитана по формуле, предложенной И.М. Распоповым (1985). Оценка продукции макрофитов для всего исследованного участка р. Паз дана по следующему алгоритму: определены площади зарослей каждого вида по полевым описаниям сообществ на станциях; подсчитана площадь, занимаемая макрофитами на всем участке р. Паз, по космическим снимкам; составлены уравнения соответствия площадей зарослей видов на станциях и всего исследованного участка р. Паз (экстраполяция данных); подсчитана продукция каждого вида.
Для сравнения флористических различий станций использовали “тепловую” карту. Для ее построения применяли пакет gplots в среде RStudio (Warnes et al., 2012; R Development…, 2013). Иерархическую кластеризацию проводили с помощью функции hclust в пакете RStudio (R Deve-lopment…, 2013). Для группировки использовали Евклидово расстояние и метод Варда.
Значимость различий проверялась процедурой ANOSIM (Analysis of Similarities test) (Anderson, Walsh, 2013). Для выделения видов, внесших наибольший вклад в различие, использовали процедуру SIMPER (Similarity Percentage Analysis).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Флористическое разнообразие. Водные растения на изученных станциях р. Паз (рис. 1) представлены 42 таксонами (40 видов и два гибрида) из 26 родов, 22 семейств (табл. S1). Наибольшее число таксонов – 33 (31 вид и два гибрида) из 20 родов, 17 семейств – зарегистрировано у сосудистых растений, на их долю приходится 79% общего состава. Криптогамные растения включают три вида харовых водорослей из двух родов, двух семейств и шесть видов мохообразных из четырех родов, трех семейств, на их долю приходится 21%. Гибриды представлены Potamogeton × nitens и P. × vepsicus.
В экологическом спектре среди сосудистых растений наиболее многочисленны гидрофиты, или истинно-водные растения, – 22 таксона (67% их состава); гигрогидрофиты (гелофиты), или воздушно-водные растения, включают 2 таксона (6%); гидрогигрофиты (гигрогелофиты), или растения уреза воды, – 8 таксонов (24%); гигрофиты, или растения сырых местообитаний, – 1 таксон (3%). Среди криптогамных растений харовые водоросли представлены исключительно гидрофитами, мохообразные – двумя гигрогидрофитами, двумя гидрогигрофитами и одним гигрофитом.
Гидрофиты присутствовали на всех обследованных станциях, в отличие от остальных экогрупп (рис. 2). Гигрогидрофиты (гелофиты) встречены на 67% из них, гидрогигрофиты (гигрогелофиты) – на 63%, а гигрофиты – на 26%. Гигрофиты представлены в составе растительного покрова на станциях с невысоким богатством истинно-водных растений.
Рис. 2.
Спектр экологических групп макрофитов на исследованных станциях. D – доля растений различных экологических групп: I – гидрофиты, II – гигрогидрофиты (гелофиты), III – гидрогигрофиты (гигрогелофиты), IV – гигрофиты.

Самые распространенные виды исследованной речной системы – Sparganium angustifolium и Myriophyllum alterniflorum (табл. S2). Они встречаются почти повсеместно. Sparganium angustifolium, кроме того, отличается значительными площадями зарослей, Equisetum fluviatile – второй по этому показателю.
Флористическое разнообразие станций реки изменяется в широком диапазоне (рис. 3). Число видов, встреченных на станциях, варьирует от 1 до 17, на более чем трети из них насчитывается ≥10 видов.

Видовое разнообразие оценено с помощью индекса Шеннона (рис. 3). Он колеблется от 0 (в единственном случае, когда на ст. р6 обнаружен лишь один вид) до 3.5 (ст. р12). Для семи станций он превышает 3.0, в среднем для изученной части реки равен 2.4.
Степень зарастания. Оценка степени зарастания всего русла р. Паз, включая зарубежную часть, по спутниковым данным, показала, что оно неравномерно (табл. S3). Участки с низкой степенью зарастания сменяются вниз по течению на более заросшие (рис. 4).
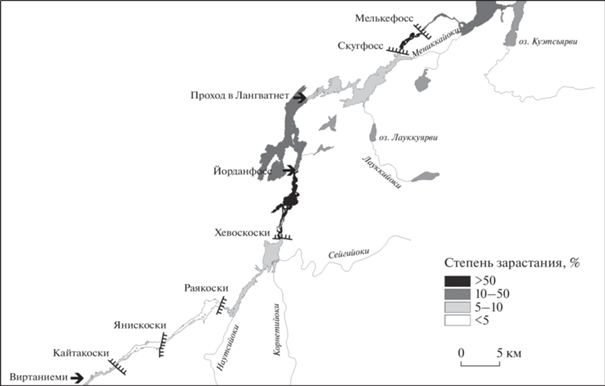
Менее всего зарастают участки реки от Виртаниеми до Хевоскоски (от <5 до 10%) и проход в Лангватнет–Скугфосс (<10%). Здесь невелики заросли и в воде, и в прибрежье (рис. 5). Наибольшие значения доли акватории, занятой зарослями макрофитов (>50%), характерны для участков с большой долей мелководий – Хевоскоски–Йорданфосс (74%) и Скугфосс–Мелькефосс (66%). В целом весь обследованный участок р. Паз зарастает почти на 30%. Несколько отличается на отдельных участках реки соотношение площадей, занятых водной и прибрежно-водной растительностью. Так, для зоны Хевоскоски–Йорданфосс отмечено наибольшее (>10%) участие в зарастания прибрежно-водных растений (рис. 5).
Рис. 5.
Доля участия водных и прибрежно-водных растений в формировании зарастания участков реки по спутниковым данным.

Характер зарастания. Три участка реки от Виртаниеми до Раякоски (станции р1–р10) характеризуются прибрежно-фрагментарным зарастанием, которое наиболее выражено в заливах и других зонах замедленного течения, например, в “тени” островов. Вдоль берегов здесь развиваются разреженные сообщества Carex spp., сменяемые с увеличением глубины на заросли Equisetum fluviatile. Сообщества гелофитов могут окаймлять пятна Sparganium angustifolium или Potamogeton gramineus и P. perfoliatus. Также представлены разреженные заросли Isoëtes spp., обычно встречающиеся совместно со Sparganium angustifolium. На участках русла с быстрым течением, рядом с островами, изредка располагаются заросли Ranunculus schmalhausenii и Sparganium angustifolium с Potamogeton gramineus. Область ниже водосброса ГЭС Янискоски отличается очень быстрым течением (>1 м/с) и твердым грунтом из валунов и скальных обломков, в таких экстремальных условиях развиваются лишь очень разреженные сообщества Myriophyllum alterniflorum.
Участок Хевоскоски–Йорданфосс (р11–р13) имеет преимущественно прибрежное зарастание. Как и на предыдущих участках, полоса Carex spp. и Equisetum fluviatile тянется вдоль берега, однако заросли в русле здесь более обильны, чем выше по течению. Они могут быть представлены мозаичными пятнами Potamogeton gramineus, P. × vepsicus, P. × nitens, P. perfoliatus, Sparganium angustifolium и Ranunculus schmalhausenii, как на ст. р11, либо располагаться по градиенту глубины, когда полосы Sparganium angustifolium сменяются подводными лугами Isoëtes lacustris.
На следующем участке Йорданфосс–проход в Лангватнет (p14–p16) характер растительного покрова в целом сходный с предыдущим – прибрежные полосы Equisetum fluviatile и Sparganium angustifolium. Однако обширные русловые заросли Potamogeton spp. и Ranunculus schmalhausenii встречаются реже, и на ст. р14 они представлены только сообществами Potamogeton perfoliatus.
Участок проход в Лангватнет–Скугфосс (p17–p22) можно охарактеризовать слабым развитием растительного покрова, по сравнению с вышележащими участками от Хевоскоски до прохода в Лангватнет. Прибрежно-фрагментарное зарастание развивается здесь лишь в заливах. Следует отметить единичную находку Nuphar pumila в составе зарослей, маленькое пятно которой обнаружено на ст. р19.
На участке Скугфосс–Мелькефосс (р23) заросли протянулись широкой полосой вдоль берега и представлены поясами Carex spp., Equisetum fluviatile и Sparganium angustifolium с Potamogeton gramineus.
Площадь зарастания русла увеличивается вниз по течению от плотины на участке Мелькефосс–Сальмиярви (p24–p27). Вначале идет прибрежное зарастание узкой полосой Sparganium angustifolium, Ranunculus schmalhausenii и Myriophyllum alterniflorum. Далее оно увеличивается, и к концу участка растительность занимает бóльшую часть русла. Здесь вдоль берега тянутся пояса Equisetum fluviatile или Carex spp., появляются сообщества Potamogeton gramineus, а затем P. perfoliatus. В некоторых местах макрофиты формируют сплошное зарастание.
На всех участках развивается комплекс мелких придонных трав с преобладанием Subularia aquatica и участием Isoёtes spp., Eleocharis acicularis и Ranunculus reptans.
Продукция. Получены средние значения фитомассы и продукции доминирующих на реке растений (рис. 6, табл. S4). Лидирующее положение по этим показателям занимает Ranunculus schmalhausenii (ст. р11), сообщества которого можно охарактеризовать как среднепродуктивные (до 1 кг/м2 в год). Однако при пересчете на весь исследованный участок реки (рис. 7) видно, что наибольший вклад в годовую продукцию органического вещества водотока вносят Potamogeton perfoliatus (3039 т/год) и P. gramineus (1908 т/год), сообщества которых формируют значительные по площади заросли. Остальные виды менее значимы, их вклад убывает в ряду: Ranunculus schmalhausenii (480 т/год) – Sparganium angustifolium (231 т/год) – Equisetum fluviatile (167 т/год) – Myriophyllum alterniflorum (74 т/год) (табл. S5).


ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Флористическое разнообразие. В сложении растительного покрова исследованного участка в среднем течении р. Паз участвовало 42 вида и гибрида водных макрофитов, из них 25 – гидрофиты, или истинно водные. В исследованиях зарубежной части водотока изначально в сумме было найдено 35 видов (Moiseenko et al., 1994), затем (Riihimäki et al., 2013) – 47 видов, причем 37 отмечены по финской полевой методике и 34 – по норвежской. По финской и российской методикам, в отличие от норвежской, фиксировались все экологические группы макрофитов водотока, и во флористический список включались мохообразные. Также следует отметить, что несколько отличались и изученные участки реки. Исследованный на территории Норвегии участок вдоль левого берега реки (Riihimäki et al., 2013) продолжается ниже впадения притока, загрязненного сточными водами комбината Печенганикель, наше исследование охватывает значительный участок русла выше по течению – от Виртаниеми до Раякоски (рис. 8).
Рис. 8.
Число видов, обнаруженных на исследованных станциях реки по материалам собственных исследований (1) (станции р1–р27), финских (2) и норвежских (3) данных (станции New, 1–9, 15), остальные обозначения, как на рис. 1.

Флористическое разнообразие р. Паз можно сопоставить с другими реками Фенноскандии. В работе Комулайнен (1990) для девяти средних и малых рек Мурманской обл. и Карелии суммарно приводится 54 вида, из них 39 – гидрофиты. Также автором показано заметное обеднение флоры к северу. Особо отмечена бедность макрофитами рек севера Мурманской обл. Комплексное исследование трех притоков оз. Онежское (Синькевич и др., 2021) продемонстрировало крайне слабое развитие водной растительности. В этих реках зарегистрировано лишь 11 видов водных сосудистых растений, встречавшихся спорадически или единично. В работах Moiseenko et al. (1994) и Riihimäki et al. (2013) указано, что по сравнению с водотоками Норвегии, р. Паз несколько уступает по разнообразию лишь самой длинной (>600 км) в стране р. Гломма, причем этот водоток также зарегулирован ГЭС. Таким образом, разнообразие водных макрофитов р. Паз можно признать весьма высоким.
Число макрофитов на исследованных станциях водотока сопоставимо с таковым на станциях зарубежной части реки (рис. 8), лишь на ряде последних станций отмечено чуть большее разнообразие макрофитов (Riihimäki et al., 2013).
В целом флора самого верхнего участка Виртаниеми–Хевоскоски, по данным настоящего исследования, относительно бедна. В более ранних работах (Moiseenko et al., 1994; Riihimäki et al., 2013) этот район не рассматривали, за исключением единственной станции (“New”), на которой тоже отмечено минимальное число таксонов водных растений по сравнению с другими участками реки (рис. 8). Русло ниже ГЭС Хевоскоски, напротив, выделяется как участок реки, характеризующейся весьма высоким разнообразием макрофитов. Здесь зарегистрированы одни из наибольших значений числа видов растений. На следующем участке русла число видов макрофитов (в среднем) несколько ниже. По нашим материалам, выделяется станция р20 в устьевой зоне р. Лауккуйоки, на которой зафиксирован второй максимум. Ниже ГЭС Скугфосс богатство остается на среднем уровне, несколько повышаясь к концу рассмотренного отрезка русла. Результаты исследований ниже всего изученного нами участка реки (Riihimäki et al., 2013) подтверждают увеличение разнообразия от оз. Сальмиярви и ниже по течению.
На исследованной акватории выделен комплекс видов с высокой встречаемостью – это Sparganium angustifolium, Myriophyllum alterniflorum, Ranunculus schmalhausenii, Equisetum fluviatile и Subularia aquatica. Данные водные растения обнаружены на более чем половине обследованных станций. Несколько менее распространены Potamogeton gramineus, P. perfoliatus, Carex rostrata и Isoëtes lacustris. Однако и они зафиксированы на более трети станций. Sparganium angustifolium, Po-tamogeton perfoliatus, Myriophyllum alterniflorum и Subularia aquatica относятся к наиболее обычным растениям и в зарубежной части р. Паз (Moiseenko et al., 1994). По обилию также можно выделить набор доминирующих видов. Наиболее обильным на изученном отрезке русла был Sparganium angustifolium. Несколько ниже показатели у Equisetum fluviatile, Myriophyllum alterniflorum и Potamogeton gramineus. Все наиболее обычные и обильные виды р. Паз характерны и для водных объектов Норвегии (Rørslett et al., 1989).
Необходимо прокомментировать ряд указанных таксонов. Из представителей рода Nuphar нами найдена только Nuphar pumila, в зарубежной части водотока обнаружены и N. lutea (L.) Smith, и N. pumila (Riihimäki et al., 2013). Среди гибридных рдестов идентифицирован (в том числе и по молекулярным данным) Potamogeton × vepsicus (P. natans L. × P. praelongus), ранее указанный для системы р. Паз как другой морфологически сходный P. × sparganiifolius Laest. ex Fries (P. gramineus × P. natans). Ranunculus schmalhausenii в работах коллег присутствует под устаревшим названием R. peltatus Schrank (Riihimäki et al., 2013). Нами обнаружен только Sagittaria natans, однако в других исследованиях (Кравченко, 2020) отмечали и его гибрид – S. × lunata C. D. Preston et Uotila (S. natans × S. sagittifolia L.).
На исследованных участках зарегистрированы четыре вида, включенные в Красную книгу Мурманской обл. (2014): Isoëtes echinospora, I. lacustris, Elatine hydropiper, Sagittaria natans. Два из них (Isoëtes echinospora и I. lacustris) имеют федеральный уровень охраны (Красная книга…, 2008).
Наиболее обычные и обильные виды на всех отрезках реки – индикаторы олиго- и олигомезотрофных условий: Sparganium angustifolium, Myriophyllum alterniflorum, Subularia aquatica, Ranunculus schmalhausenii, Isoëtes echinospora и I. lacustris, что связано с низким уровнем содержания органических и биогенных соединений и высокой прозрачностью вод реки (Сандимиров, 2012; Новиков, Харламова, 2016).
Интересен набор видов, встреченных лишь на ст. p27 – при впадении р. Мениккайоки: Callitriche hermaphroditica, Elatine hydropiper, Lemna trisulca и Myriophyllum sibiricum. Все они указаны и для зарубежной части водотока (Moiseenko et al., 1994; Riihimäki et al., 2013). С большой вероятностью отличие в химическом составе воды на ст. p27 определяет наличие данных видов. Именно здесь зафиксирована самая высокая минерализация воды, в ~2 раза превышающая характерную для р. Паз (табл. 1). Причина повышенной минерализации в геологическом строении истоковой области р. Мениккайоки – района Глухой плотины, что показано и для наземных растений-кальцефилов, сконцентрированных в этом месте (Кравченко, 2020).
На основе флористического состава проведена кластеризация станций (рис. 9). По встречаемости макрофитов на “тепловой” карте выделяются два кластера. Первый из них сложен наиболее часто встречающимися видами (Ranunculus schmalhausenii, Myriophyllum alterniflorum, Potamogeton gramineus, Sparganium angustifolium, P. perfoliatus, Subularia aquatica, Isoëtes lacustris и Equisetum fluviatile), которые на этих станциях и наиболее обильны. Все остальные формируют вторую группу.
Рис. 9.
Тепловая карта, построенная по обилию макрофитов в баллах на станциях р. Паз. Цветом обозначено относительное обилие вида на станции от отсутствия таксона (белый цвет) до максимального обилия (темно-серый). Виды: Call cor – Calliergon cordifolium, Call cus – Calliergonella cuspidata, Calli her – Callitriche hermaphroditica, Calli pal – Callitriche palustris, Calt pal – Caltha palustris, Care acu – Carex acuta, Care aqu – Carex aquatilis, Care ros – Carex rostrata, Care ves – Carex vesicaria, Char vir – Chara virgata, Coma pal – Comarum palustre, Elat hyd – Elatine hydropiper, Eleo aci – Eleocharis acicularis, Equi flu – Equisetum fluviatile, Hipp vul – Hippuris vulgaris, Isoё ech – Isoёtes echinospora, Isoё lac – Isoёtes lacustris, Lemn tri – Lemna trisulca, Meny tri – Menyanthes trifoliata, Myri alt – Myriophyllum alterniflorum, Myri sib – Myriophyllum sibiricum, Nite fle/opa – Nitella flexilis vel opaca, Nite opa – Nitella opaca, Nuph pum – Nuphar pumila, Persi amp – Persicaria amphibia, Pota х nit – Potamogeton × nitens (P. gramineus × P. perfoliatus), Pota х vep – Potamogeton × vepsicus (P. natans × P. praelongus), Pota alp – Potamogeton alpinus, Pota ber – Potamogeton berchtoldii, Pota gra – Potamogeton gramineus, Pota per – Potamogeton perfoliatus, Ranu rep – Ranunculus reptans, Ranu sch – Ranunculus schmalhausenii, Sagi nat – Sagittaria natans, Scor rev – Scorpidium revolvens, Scor sco – Scorpidium scorpioides, Spar ang – Sparganium angustifolium, Subu aqu – Subularia aquatica, Utri vul – Utricularia vulgaris, Utri int – Utricularia intermedia, Warn exa – Warnstorfia exannulata, Warn flu – Warnstorfia fluitans.

Станции также поделены на две группы по составу таксонов. В первую группу вошли станции р11–р13, р15, р16, р23, р26, р27, где с наибольшим обилием представлен комплекс обычных для реки видов, во вторую – станции p1–p10, р14, р17–р22, р24 и р25, на которых обычные для реки виды в основном имеют низкое обилие. На станциях первой группы значительна доля истинно-водных растений (гидрофитов) – ≥6 видов. Кроме того, эти участки отличаются высоким зарастанием, причем непосредственно в русле реки (до глубин >2 м): Хевoскоски–Йорданфосс, Мелькефосс–Сальмиярви. На последнем участке встречаются зоны сплошного зарастания.
Станции второй группы отличаются неблагоприятными условиями для развития макрофитов – затопленные участки вырубленного леса и болот, с пнями, корягами, торфяными отложениями (р4, р17–р20), с обломочно-скальным грунтом и экстремальной скоростью течения (>1 м/с) ниже водослива ГЭС (р6).
Различия между выделенными группами достоверны (ANOSIM: R = 0.72, p = 0.0001). Процедура SIMPER показывает, что наибольший вклад (31%) в различия между кластерами станций вносит наличие и обилие Isoëtes lacustris, Equisetum fluviatile и Potamogeton gramineus (табл. S6).
Зарастание и продукция. Прослеживается повышение степени зарастания вниз по течению реки. Максимально зарастают области средней и нижней частей исследованного участка. Это небольшой по длине и площади отрезок Скугфосс–Мелькефосс и участок Хевоскоски–Йорданфосс со средними морфометрическими характеристиками (табл. 1) и относительно большой долей мелководий (Поликарпова, Хлебосолов, 2009), что благоприятно для формирования зарослей.
Степень зарастания всего исследованного участка реки достигает ~30%, максимальная – 74%. Таким образом, зарастание водотока можно считать умеренным (Бобров, Чемерис, 2006). Столь высокое по степени зарастание вместе с его прибрежно-фрагментарным типом с участками сплошного зарастания нехарактерно для водотоков данной природной зоны. Для северных рек, особенно районов с молодыми формами рельефа как в Мурманской обл., обычно слабое (до 15–20%), фрагментарное зарастание и отсутствие сплошного зарастания (Бобров, Чемерис, 2008).
Зарастанием речных участков р. Паз напоминает расположенные заметно южнее реки возвышенностей Русской равнины со сглаженным рельефом (Бобров, Чемерис, 2008). Причина такой несколько повышенной степени зарастания, по-видимому, кроется в формировании плотинами гидроэлектростанций благоприятного гидрологического режима “озерного” типа бо́льшей части р. Паз. Происходит снижение скорости течения реки в зоне водохранилищ, формирование в таких местах подходящих для растений донных отложений (песка и ила). Стабильный уровень воды, как предполагается в работе (Moiseenko et al., 1994), вместе с ее высокой прозрачностью, по-видимому, обеспечивают высокое разнообразие макрофитов и обилие их зарослей. Отметим, что уровенный режим, обусловленный работой ГЭС, – один из основных факторов в зарастании антропогенно трансформированных водных объектов (Поддубный и др., 2018, 2022). Кроме того, подобная картина увеличения степени зарастания зарегулированного водотока описана для других норвежских рек (Ylikörkkö et al., 2015). Другой причиной такого развития растительного покрова выступает отепляющий эффект достаточно крупной реки, текущей с юга (Фащевский, 2007).
Наши данные по биомассе водных растений, полученные летом 2019 г., были сопоставлены с таковыми 2003–2006 гг. в работе Кирилловой (2012). Средняя сырая биомасса доминирующих водных растений реки в материалах предшествующей работы изменялась в пределах 18.5–131 г/м2, по нашим данным, она больше на порядок – 196–1528 г/м2.
Так, наибольшие значения сырой фитомассы в 2003–2006 гг. (Кириллова, 2012) зарегистрированы у Equisetum fluviatile (131 г/м2) и группы видов рода Carex (117 г/м2). Меньшие значения отмечены у Ranunculus schmalhausenii (“Вatrachium peltatum”) (104 г/м2), группы видов рода Sparganium (51) и видов рода Potamogeton (85), самое низкое – у Comarum palustre (18.5 г/м2). Нами в 2019 г. наиболее высокие значения сырой фитомассы получены у Ranunculus schmalhausenii (1528 г/м2) и Potamogeton perfoliatus (464–936), наименьшее – у Equisetum fluviatile (196–228). Воздушно-сухая фитомасса была наибольшая также у Ranunculus schmalhausenii (120 г/м2) и Potamogeton perfoliatus (44–96), наименьшая – у Equisetum fluviatile (26–30 г/м2). Эти показатели соответствуют таковым для северных популяций Ranunculus schmalhausenii – 16–80 г/м2 воздушно-сухой фитомассы при проективном покрытии 50–80% (Bobrov et al., 2015) и Potamogeton perfoliatus – 600–6000 г/м2 сырой фитомассы и 92–672 г/м2 воздушно-сухой фитомассы (Чемерис, Бобров, 2020).
Значительные различия в полученных данных могут быть следствием разных методических задач. Мы для оценки продуктивности доминирующих растений проводили укосы в наиболее развитых и обширных зарослях растений, чтобы зафиксировать максимальные значения биомассы. Исследования Н. Р. Кирилловой (2012) были направлены на определение значимых для водоплавающих птиц кормовых водных растений заповедника “Пасвик”, и укосы проводили в сообществах с различной степенью проективного покрытия – от слабого до высокого.
Выводы. Озерно-речная система р. Паз отличается более богатым и хорошо развитым растительным покровом по сравнению с другими реками северной Фенноскандии. Этому способствует ряд факторов. Во-первых, это благоприятный гидрологический режим, создаваемый плотинами гидроэлектростанций. Снижаются скорости течения реки в зоне водохранилищ, происходит аккумуляция мелкодисперсного субстрата. При этом поддерживается постоянный уровень воды, а водная толща прогревается. Во-вторых, оптимальный для водных растений световой режим: малый процент затененных участков русла и высокая прозрачность воды. В-третьих, умеренное содержание биогенных соединений, поступающих с водосбора, и локальный приток минерализованных вод достаточны для развития растений в течение всего вегетационного периода. В-четвертых, отепляющий эффект долины достаточно крупной реки. Состояние водотока по характеристикам растительного покрова на исследованном участке можно признать стабильным и благополучным.
Список литературы
Бобров А.А., Чемерис Е.В. 2006. Изучение растительного покрова ручьев и рек: методика, приемы, сложности // Гидроботаника-2005: Матер. VI Всерос. шк.-конф. (пос. Борок, 11–16 окт. 2005 г.). Рыбинск: Рыбинский Дом печати. С. 181.
Бобров А.А., Чемерис Е.В. 2008. Речная растительность на севере европейской России: предварительные результаты // Экосистемы малых рек: биоразнообразие, экология, охрана: Лекции и матер. докл. Всерос. школы-конф. (Борок, 18–21 ноября 2008 г.). Борок: Принтхаус. С. 76.
Боровичев Е.А., Бойчук М.А. 2018. Мохообразные заповедника “Пасвик”. Петрозаводск: Карельск. науч. центр РАН.
Водный кадастр Российской Федерации. Реки и озера Российской Федерации (ресурсы, режим и качество воды). Ежегодный справочник 2018 г. 2019. Санкт-Петербург: ФГБУ “ГГИ”. 96 с. URL: http://www.hydrology.ru/sites/default/files/Books/spra-vochnik_200220.pdf
Демин В.И. 2012. Основные климатические тенденции на Кольском полуострове за период инструментальных метеорологических измерений // Труды Кольского научного центра РАН. № 3(10). С. 98.
Канева Н.Р. 2008. Флористические находки в заповеднике “Пасвик” (Мурманская область) // Бюллетень Московского общества испытателей природы. Отдел биол. Т. 113. Вып. 3. С. 63.
Канева Н.Р., Ламзов Д.С. 2009. Новые виды и новые места обитания видов // Летопись природы заповедника “Пасвик”. Книга 11 (2004). Апатиты. С. 83.
Катанская В.М. 1981. Высшая водная растительность континентальных водоемов СССР. Методы изучения. Ленинград: Наука.
Качество поверхностных вод Российской Федерации. Ежегодник 2019. 2020. Ростов-на-Дону. URL: http://gidrohim.com/sites/default/files/Ежегодник%-202019_0.pdf
Кириллова Н.Р. 2012. Ресурсная оценка кормовых угодий растительноядных водоплавающих птиц заповедника “Пасвик” (Мурманская область) по биомассе водных растений // Водное хозяйство России: проблемы, технологии, управление. № 1. С. 92.
Кириллова Н.Р. 2013. Исследования водной растительности в Мурманской области // Вестник Кольского научного центра РАН. № 1(12). С. 66.
Кожин М.Н., Королeва Н.Е., Кравченко А.В. и др. 2020. История и основные итоги изучения флоры сосудистых растений и растительности Зеленого Пояса Фенноскандии в пределах Мурманской области // Труды Карельского научного центра РАН. № 8. С. 26. https://doi.org/10.17076/bg1084
Комулайнен С.Ф. 1990. Макрофиты в малых реках Карелии и Кольского полуострова // Деп. в ВИНИТИ 05.01.1990, № 75-890. Киев.
Костина В.А. 2003. Сосудистые растения заповедника “Пасвик” (аннотированный список видов) // Флора и фауна заповедников. Москва: Комиссия РАН по сохранению биоразнообразия (секция заповедного дела); Институт проблем экологии и эволюции им. А.Н. Северцова РАН. Вып. 103. 44 с.
Кравченко А.В. 2020. Сосудистые растения заповедника “Пасвик” и смежной территории Мурманской области. Петрозаводск: Карельский науч. центр РАН.
Кравченко А.В., Боровичев Е.А., Химич Ю.Р. и др. 2017. Значимые находки растений, лишайников и грибов на территории Мурманской области // Труды Карельского научного центра РАН. № 7. С. 34.
Красная книга Мурманской области. 2014. Кемерово: Изд-во “Азия-принт”.
Красная книга Российской Федерации (растения и грибы). 2008. Москва: Товарищество науч. изданий КМК.
Летопись природы заповедника “Пасвик”. Книга 2: 1995 г. 1998. Мурманск: Изд-во НИЦ “Пазори”.
Новиков М.А., Харламова М.Н. 2016. Исследование спектральных характеристик флуоресценции природных вод Кольского полуострова // Труды ВНИРО. Вып. 161. С. 181.
Поддубный С.А., Чемерис Е.В., Бобров А.А. 2018. Влияние режима уровня воды на зарастание мелководий Рыбинского водохранилища (обзор) // Биология внутр. вод. № 4. С. 38.https://doi.org/10.1134/S0320965218030142
Поддубный С.А., Чемерис Е.В., Кутузов А.В. и др. 2022. Динамика высшей водной растительности защищенного мелководья в связи с уровнем воды в Волжском плесе Рыбинского водохранилища // Биология внутр. вод. № 2. С. 136. https://doi.org/10.31857/S0320965222020085
Поликарпова Н.В., Хлебосолов Е.И. 2009. Ключевые участки для мониторинга редких видов // Летопись природы заповедника “Пасвик”. Книга 11 (2004). Апатиты: Изд-во Кольского науч. центра РАН. С. 14.
Распопов И.М. 1985. Высшая водная растительность больших озер Северо-Запада СССР. Ленинград: Наука.
Сандимиров С.С. 2012. Современное гидрохимическое состояние озерно-речной системы реки Пасвик (Кольский полуостров) // Труды Кольского научного центра РАН. № 3(10). С. 88.
Синькевич С.М., Ефремов Д.А., Тимофеева В.В. и др. 2021. Экологическая оценка состояния малых рек среднетаежной подзоны Карелии // Труды Карельск. науч. центра РАН. № 7. С. 46. https://doi.org/10.17076/them1417
Фащевский Б.В. 2007. Экологическое значение поймы в речных экосистемах // Ученые записки РГГМУ. № 5. С. 118.
Чемерис Е.В., Бобров А.А. 2020. Продуктивность рдестов (Potamogeton, Stuckenia, Potamogetonaceae) в реках севера европейской России // Водные ресурсы. Т. 47. № 1. С. 1.https://doi.org/10.31857/S0321059620010198
AMAP. 2017. Adaptation Actions for a Changing Arctic: Perspectives from the Barents Area. Arctic Monitoring and Assessment Programme (AMAP), Oslo, Norway. xiv + 267 pp. https://www.amap.no/documents/ doc/adaptation-actions-for-a-changing-arctic-perspectives-from-the-barents-area/1604
Amundsen P.-A., Primicerio R., Smalås A. et al. 2019. Long-term ecological studies in northern lakes–challenges, experiences, and accomplishments // Limnol., Oceanogr. V. 64. P. S11. https://doi.org/10.1002/lno.10951
Anderson M.J., Walsh D.C. 2013. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? // Ecological Monographs. V. 83. P. 557. https://doi.org/10.1890/12-2010.1
Bobrov A.A., Zalewska-Gałosz J., Jopek M., Movergoz E.A. 2015. Ranunculus schmalhausenii (section Batrachium, Ranunculaceae), a neglected water crowfoot endemic to Fennoscandia – a case of rapid hybrid speciation in postglacial environment of North Europe // Phytotaxa. V. 233. № 2. P. 101. https://doi.org/10.11646/phytotaxa.233.2.1
Brittain J.E., Heino J., Friberg N. et al. 2020. Ecological correlates of riverine diatom and macroinvertebrate alpha and beta diversity across Arctic Fennoscandia // Freshwater Biol. Spec. Issue. P. 1. https://doi.org/10.1111/fwb.13616
Culp J.M., Lento J., Goedkoop W. et al. 2012. Developing a circumpolar monitoring framework for Arctic freshwater biodiversity // Biodiversity. V. 13. P. 215.
Heino J., Muotka T., Mykrä H. et al. 2003. Defining macroinvertebrate assemblage types of headwater streams: Implications for bioassessment and conservations // Ecological Applications. V. 13. P. 842. https://doi.org/10.1890/1051-0761(2003)013[0842:D-MATOH]2.0.CO;2
Heino J., Virkkala R., Toivonen H. 2009. Climate change and freshwater biodiversity: Detected patterns, future trends and adaptations in northern regions // Biological Reviews. V. 84. P. 39. https://doi.org/10.1111/j.1469-185X.2008.00060.x
Moiseenko T., Mjelde M., Brandrud T. et al. 1994. Pasvik River watercourse, Barents Region: Pollution impacts and ecological responses. Investigations in 1993. Oslo: NIVA-report OR-3118.
Mustonen K.-R., Mykrä H., Marttila H. et al. 2018. Thermal and hydrologic responses to climate change predict marked alterations in boreal stream invertebrate assemblages // Global Change Biology. V. 24. P. 2434. https://doi.org/10.1111/gcb.14053
Preston C.D., Uotila P. 2009. Sagittaria × lunata, a binomial for the widespread North European hybrid between S. natans and S. sagittifolia (Alismataceae) // Ann. Bot. Fenn. V. 46. № 3. P. 215. https://doi.org/10.5735/085.046.0304
Prowse T.D., Wrona F.J., Reist J.D. et al. 2006. Climate change effects on hydroecology of Arctic freshwater ecosystems // Ambio. V. 35. № 7. P. 347. https://doi.org/10.1579/0044-7447(2006)35[347:CC-EOHO]2.0.CO;2
R Development Core Team. R: A language and environment for statistical computing. 2013. http://www.r-project.org.
Riihimäki J., Mjelde M., Hellsten S. 2013. Ecological status assessment of Lake Inari and River Pasvik and recommendations for biological monitoring using aquatic macrophytes // EU ENPI – project Trilateral cooperation on environmental challenges in the joint border area: Kolarctic ENPI cross-border cooperation report. https://www.pasvikmonitoring.org/pdf/raportteja/En-pi%20macrophyte%20report_Final.pdf
Rørslett B., Mjelde M., Johansen S.W. 1989. Effects of hydropower development on aquatic macrophytes in Norwegian rivers: Present state of knowledge and some case studies // Regulated Rivers Research & Management. V. 3. P. 19. https://doi.org/10.1002/rrr.3450030104
Shannon C.E. 1948. A mathematical theory of communication // The Bell System Technical Journal. V. 27. P. 379.
Tolonen K.E., Tokola L., Grönroos M. et al. 2016. Hierarchical decomposition of trait patterns of macroinvertebrate communities in subarctic streams // Freshwater Science. V. 35. P. 1032. https://doi.org/10.1086/687966
Warnes G.R., Bolker B., Bonebakker L. et al. 2012. GPLOTS: Various R programming tools for plotting data (comprehensive R archive network). R package version 2.8.0. http://cran.r-project.org/web/packages/gplots/index.html
Wrona F.J., Prowse T.D., Reist J.D. et al. 2006. Climate change effects on aquatic biota, ecosystem structure and function // Ambio. V. 35. № 7. P. 359. https://doi.org/10.1579/0044-7447(2006)35[359:CC-EOAB]2.0.CO;2
Ylikörkkö J., Christensen G., Kashilin N. et al. 2015. Environmental challenges in the joint border area of Norway, Finland and Russia. Reports 41/edit by Jelkänen E. Centre for Economic Development, Transport and the Environment for Lapland. Kokkola: Juvenes Print. https://www.doria.fi/handle/10024/104779
Дополнительные материалы
- скачать ESM.doc
- Приложение 1.
Инструменты
Биология внутренних вод