

Биология внутренних вод, 2022, № 4, стр. 372-380
Таксономический состав и экологическая характеристика диатомовых водорослей альгоценозов бентали устья крупной эвтрофно-гипертрофной реки (р. Ока, Россия)
Т. Хедаириа a, *, А. Г. Охапкин a, С. И. Генкал b, А. М. Глущенко c, М. С. Куликовский c
a Институт биологии и биомедицины, Нижегородский государственный университет им. Н.И. Лобачевского
Нижний Новгород, Россия
b Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл, Россия
c Институт физиологии растений Российской академии наук
Москва, Россия
* E-mail: khedairia.ta@gmail.com
Поступила в редакцию 03.12.2020
После доработки 03.02.2022
Принята к публикации 03.03.2022
- EDN: ZJXQCX
- DOI: 10.31857/S032096522204012X
Аннотация
Определены видовой состав, таксономическая структура и экологические показатели донных диатомовых водорослей в устье крупной эвтрофно-гипертрофной реки. Выявлены сходство и различие таксономической структуры, набора массовых и редких видов диатомовых водорослей в разных типах сообществ бентали (эпипилоне, эпилитоне и эпифитоне). Продемонстрировано удовлетворительное соответствие оценки условий обитания Bacillariophyta по их составу физико-химическим параметрам воды.
ВВЕДЕНИЕ
Бентосные водоросли играют значительную роль в формировании биоразнообразия водоемов, их продуктивности и качества воды. Европейская Рамочная водная директива ЕС провозгласила фитобентос одним из элементов оценки биологического качества для определения экологического состояния рек и озер (Семенченко, 2004). Диатомовые, наиболее разнообразная и многочисленная составляющая фитобентоса (Round et al., 1990), хорошо изучены в экологическом отношении (Coste et al., 2009; Kelly et al., 2009), считаются чувствительными показателями изменения окружающей среды и активно применяются для биологического мониторинга лотических экосистем (Bere, Tundisi, 2010). Водоемы и водотоки бассейна р. Волга в отношение альгоценозов бентали изучены фрагментарно (Волга…, 1978; Метелева, Девяткин, 2005; Метелева, 2013; Насибулина, Тлекова, 2015). Для крупных рек России обобщенные сведения известны для р. Днепр (Владимирова, 1978) и р. Енисей (Левадная, 1986). Подробно охарактеризованы закономерности формирования состава и структуры фитоперифитона для водных объектов Карелии (Комулайнен, 2004). Водоросли, населяющие различные биотопы р. Ока, изучают ~100 лет, но как компоненты фитопланктона, специальные исследования фитобентоса реки ранее не проводили.
Цель работы – определить видовой состав, охарактеризовать таксономическую структуру и оценить экологическое состояние диатомовых водорослей альгоценозов бентали устья р. Ока.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Пробы фитобентоса отбирали на трех станциях, расположенных в рипали правобережья р. Ока в пределах г. Нижний Новгород, в первых декадах июля и ноября 2018 г. (рис. 1). Станция 1 находилась в 400 м выше нового Окского моста (56.294045° с.ш., 43.966690° в.д.), ст. 2 – 400 м ниже моста (56.301329° с.ш., 43.966726° в.д.) и ст. 3 – 900 м ниже моста (56.305490° с.ш., 43.967413° в.д.) на глубине 1.0–1.5 м. Температура воды, достигавшая в июле 24–27°С, в ноябре сильно снизилась (4.9°С) и была почти одинакова на всех станциях. Прозрачность и летом, и осенью была стабильно низкой (0.5–0.8 м), электропроводность в ноябре колебалась в пределах 634–663.1 μS/cm. Активная реакция среды достигала в июле 8.3–8.7, осенью немного снизилась (7.8–8.1). Для характеристики абиотических условий обитания бентосных альгоценозов анализировали данные по гидрохимическому составу вод р. Ока в пределах города, предоставленные Верхне-Волжским управлением по гидрометеорологии и контролю окружающей среды (табл. 1).
Рис. 1.
Карта-схема правобережья устья р. Оки в пределах г. Нижний Новгород. 1–3 – станции отбора проб.

Таблица 1.
Основные гидрофизические и гидрохимические показатели вод устья р. Ока (M ± m)
| Показатель | Безледный период 2011 г. |
Июнь–июль 2018 г. |
Октябрь–ноябрь 2018 г. |
|---|---|---|---|
| Цветность, град | 26.7 ± 8.2 | – | – |
| pH | 8.12 ± 0.14 | 8.23 ± 0.16 | 8.49 ± 0.04 |
| Взвешенные вещества, мг/л | 9.6 ± 2.2 | 23.3 ± 3.53 | 12.4 ± 2.03 |
| Растворенный кислород, мг О2/Л | 8.90 ± 0.74 | 8.60 ± 0.09 | 10.2 ± 0.42 |
| Углекислый газ, СО2 мг/л | 8.66 ± 0.64 | – | – |
| Сумма ионов, мг/л | 545 ± 29 | 349* | 596* |
| Ионы Са2+, мг/л | 91.3 ± 0.4 | 60.2* | 85.6* |
| Ионы Mg2+, мг/л | 31.5 ± 7.2 | 26.7* | 25.5* |
| Сумма ионов, Na+ + K+, мг/л | 41.6 ± 6.8 | 9.6* | 46.0* |
| Гидрокарбонаты, HC${\text{O}}_{3}^{ - }$, мг/л | 156.9 ± 39.0 | 128.0* | 229.0* |
| Сульфаты, S${\text{O}}_{4}^{{2 - }}$, мг/л | 163.8 ± 26.2 | 106.0* | 182.0* |
| Хлориды, Cl–, мг/л | 24.9 ± 3.3 | 27.1* | 27.6* |
| БПК5, мг О2/л | 2.73 ± 0.35 | 3.0 ± 0.56 | 2.19 ± 0.31 |
| ХПК, мг О/л | 28.3 ± 0.98 | 24.4 4.6 | 28.8 ± 1.7 |
| Азот минеральный, мг N/Л | 2.76 ± 0.78 | 1.67 ± 0.21 | 1.12 ± 0.06 |
| Фосфаты, P${\text{O}}_{4}^{{2 - }}$, мг P/Л | 129 ± 44 | 30.0* | 91 ± 46 |
| Фосфор общий, Pобщ, мг P/Л | 149 ± 26 | 99 ± 34 | 95 ± 26 |
| Кремний, Si, мг/л | 5.43 ± 1.28 | 0.50* | 1.80* |
| Железо общее, Fe, мг/л | – | 0.08 ± 0.01 | 0.02* |
Река Ока, второй по величине русла и объему стока крупный приток р. Волга (длина 1500 км, площадь водосбора 24 500 км2), имеет на протяжении последних десятилетий стабильно низкое качество вод, оцениваемое четвертым классом. Воды ее устьевого участка, как и прежде (Охапкин и др., 2015; Джамалов и др., 2017), отличаются высокими мутностью, концентрациями минеральных форм азота, фосфора, органического вещества, тяжелых металлов и других компонентов загрязнения за счет поступления этих компонентов с организованым и рассеянным стоком.
Грунты сформированы илами и сильно заиленными песками с вкраплением каменистого субстрата (в основном известняка). Высшая водная и прибрежно-водная растительность развиты слабо, образованы несомкнутыми зарослями стрелолиста, кубышки и рдеста. Пробы отбирали с поверхности мягких грунтов (эпипелон), камней (эпилитон) и высших водных и прибрежно-водных растений (эпифитон) с использованием ряда методик (Водоросли…, 1989; Комулайнен, 2003; Неврова и др., 2015). Всего отобрано 25 количественных и качественных проб.
Диатомовые обрабатывали путем горячего окисления перекисью водорода, постоянные препараты готовили с применением среды Naphrax (показатель преломления = 1.74, Brunel Microscopes Ltd.), идентификацию проводили по определителям (Krammer, H. Lange-Bertalot, 1986, 1988, 1991а, 1991б; Diatomeen…, 2013; Куликовский и др., 2016).
Количественную представленность видов оценивали по модифицированной глазомерной балльной шкале Стармаха (Водоросли…, 1989): 1 – единично, 3 – немного, 5 – умеренно, 7 – много, 9 – массово. Анализировались частоты встречаемости видов (% проб, в которых вид обнаружен) и доминирования (% проб, в которых обилие вида было семь или девять баллов). Экологическую характеристику диатомовых приводили по (Баринова и др., 2019). Для построения дендрограмм сходства структуры сообществ в зависимости от субстрата, сезона и станции отбора проб с использованием кластерного анализа, применяли метод средней связи, индекс Ренконена и открытую программу среды R (Якимов и др., 2016).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Выявлено 133 вида и внутривидовых таксона Bacillariophyta, относящихся к трем классам, 13 порядкам, 23 семействам и 47 родам. Список видов с указанием экологических группировок (1 – эпипелон, 2 – эпилитон, 3 – эпифитон) приведен ниже:
Класс Coscinodiscophyceae
Порядок Coscinodiscales: Actinocyclus normanii (Greg.) Hust. (1, 2, 3).
Порядок Aulacosirales: Aulacoseira ambigua (Grun.) Sim. (1, 2, 3), A. granulata (Ehr.) Sim. (1, 2, 3), A. subаrctica (O. Müll.) Haw. (1, 2, 3).
Порядок Melosirales: Melosira varians Ag. (1, 2, 3).
Порядок Thalassiosirales: Cyclotella meneghiniana Kütz. (1, 2, 3), Handmannia radiosa (Grun.) Kociolek et Khurs. (2), Stephanodiscus hantzschii Grun. (1, 2, 3), S. invisitatus Hohn et Hellerm. (1, 2, 3), S. neoastraea Håk. et Hick. (1, 2, 3).
Класс Fragilariophyceae
Порядок Fragilariales: Asterionella formosa Hass. (2), Diatoma moniliformis (Kütz.) Williams (2), D. tenuis Ag. (1, 2, 3), D. vulgaris Bory (1, 2, 3), Fragilaria capucina Desm. (2, 3), F. vaucheriae (Kütz.) Peters. (1, 2, 3), Fragilariforma virescens var. subsalina (Grun.) Bukht. (2), Meridion circulare (Grev.) Ag.(1, 2), M. constrictum (Ralfs) Kütz. (2), Odontidium mesodon (Kütz.) Kütz. (1), Pseudostaurosira brevistriata (Grun.) Williams et Round (1, 2, 3), P. parasitica (W. Sm.) Morales (2), Punctastriata discoidea Flower (2), P. glubokoensis Williams, Chudaev et Gololobova (2), P. lancettula (Schum.) Hamilt. et Siver (1, 2), Staurosira construens (Ehr.) Cl. et Möller (2), S. grigorszkyi Ács, Morales et Ector (2), S. mutabilis (W. Sm.) Pfitz. (1, 2, 3), S. oldenburgiana (Hust.) Lange-Bert. (3), S. venter (Ehr.) Cl. et Möller (1, 2), Staurosirella martyi (Hérib.) Morales et Manoylov (1, 2, 3), S. pinnata (Ehr.) Williams et Round (1, 2, 3), Ulnaria acus (Kütz.) Ab. (1, 2, 3), U. ulna (Nitzsch) Comp. (1, 2, 3).
Класс Bacillariophyceae
Порядок Achnanthales: Achnanthidium affine (Grun.) Czarn. (1, 2, 3), A. anastasiae (Kacz.) Chudaev et Gololobova (2, 3), A. minutissimum (Kütz.) Czarn. (1, 2), Cocconeis lineata Ehr. (1, 3), C. pediculus Ehr. (1, 2, 3), C. placentula Ehr. (1, 2, 3), Planothidium dubium (Grun.) Round et Bukht. (1), P. frequentissimum (Lange-Bert.) Lange-Bert. (1, 2, 3), P. lanceolatum (Bréb. ex Kütz.) Lange-Bert. (1, 2, 3), P. reichardtii Lange-Bert. et Werum (1, 2), P. rostratum (Østr.) Lange-Bert. (1, 2, 3).
Порядок Naviculales: Amphipleura pellucida (Kütz.) Kütz. (1, 2, 3), Calonies amphisbaena (Bory) Cl. (1), Craticula cuspidata (Kutz.) Mann (2), Frustulia vulgaris (Thw.) De Toni (1), Gyrosigma acuminatum (Kütz.) Rab. (1, 2, 3), G. attenuatum (Kütz.) Rab. (2), G. exilis (Grun.) Reimer (1, 2), G. obtusatum (Sull. et Wormley) Boyer, Hippodonta capitata (Ehr.) Lange-Bert., Metz. et Witkowski (1, 2, 3), H. linearis (Østr.) Lange-Bert., Metz. et Witkowski (2), Luticola goeppertiana (Bleisch) Mann (1, 2), L. permuticopsis Kopalova et van de Vijver (2), Navicula antonii Lange-Bert. (1, 2, 3), N. capitatoradiata Germ. (1,2,3), N. cryptocephala Kütz. (1, 2, 3), N. menisculus Schum (2, 3), N. radiosa Kütz. (1, 2, 3), N. rostellata Grun. (1), N. tripunctata (O. Müll.) Bory (1, 2, 3), N. veneta Kütz. (2, 3), N. viridula (Kütz.) Ehr. (2), Neidium dubium (Ehr.) Cl. (1), Pinnularia neohalophila Kulikovskiy, Genkal et Mikheeva (2, 3), Sellaphora bacillum (Ehr.) Mann (2), S. pupula (Kütz.) Mereschk. (1, 2, 3), S. seminulum (Grun.) Mann (2), Stauroneis anceps Ehr. (2).
Порядок Cymbellales: Brebissonia lanceolata (Ag.) Mahoney et Reimer (2), Cymbella compacta Østr. (1, 2, 3), C. cymbiformis Ag. (2), C. neocistula Krammer (2, 3), C. tumida (Bréb.) van Heurck (1, 2), Encyonema caespitosum Kütz. (1, 2, 3), E. leibleinii (Ag.) Silva, Jahn, Ludwig et Menez. (1, 2, 3), E. minutum (Hilse) Mann (2, 3), E. reichardtii (Kramm.) Mann (2), E. silesiacum (Bleisch) Mann (2, 3), E. ventricosum (Ag.) Grun. (1, 2, 3), Gomphonella olivacea (Hornemann) Rabenhorst (1, 2, 3), Gomphonema acuminatum Ehr. (2), G. augur Ehr. (2), G. italicum Kütz. (2, 3), G. geisslerae Reich. et Lange-Bert. (1, 2, 3), G. parvulum Kütz. (1, 2, 3), G. pumilum (Grun.) Reich. et Lange-Bert. (1, 2, 3), G. truncatum Ehr. (3), G. supertergestinum Reich. (3), G. utae Lange-Bert. et Reich. (1), Navigeia decussis (Østr.) Bukht. (1, 2, 3), Placoneis clementis (Grun.) Cox (2), P. gastrum (Ehr.) Mereschk. (2, 3), P. pseudanglica Cox, nom. Illeg. (3), Rhoicosphenia abbreviata (Ag.) Lange-Bert. (1, 2, 3).
Порядок Thalassiophysales: Amphora copulata (Kütz.) Schoem. et Archib. (1, 2, 3), A. ovalis (Kütz.) Kütz. (1, 2, 3), A. pediculus (Kütz.) Grun. ex Schmidt (1, 2, 3), Halamphora obscura (Krasske) Levkov (2), H. veneta (Kütz.) Levkov (2).
Порядок Rhopalodiales: Epithemia sorex Kütz. (3).
Порядок Eunotiales: Eunotia bilunaris (Ehr.) Schaar. (3), E. incisa W. Sm. ex W. Greg. (3), E. pectinalis (Kütz.) Rab. (3).
Порядок Bacillariales: Hantzschia amphioxys (Ehr.) Grun. (2), Nitzschia acicularis (Kütz.) W. Sm. (1, 2, 3), N. angustata (W. Sm.) Grun. (1, 2, 3), N. clausii Hantzsch (2), N. constricta (Kützing) Ralfs in Pritchard (1, 2), N. dealpina Lange-Bert. et Hofm. (2), N. dissipata (Kütz.) Rabenh. (1, 2, 3), N. frustulum (Kütz.) Grun, (1, 2, 3), N. intermedia Hantzsch (2), N. linearis W. Sm. (1, 2, 3), N. palea (Kütz.) W. Sm. (1, 2, 3), N. recta Hantzsch ex Rabenh. (2), N. sigmoidea (Nitzsch) W. Sm. (2), N. sublinearis Hust. (2, 3), N. vermicularis (Kütz.) Hantzsch (1, 2, 3), T. calida (Grun.) Mann (2), T. hungarica Grun. (1, 2, 3), T. levidensis W. Sm. (1, 2), T. salinarum (Grun.) Pellet. (2).
Порядок Surirellales: Surirella angusta Kütz. (1, 2, 3), S. brebissonii Krammer et Lange-Bert. (2), S. librile (Ehr.) Ehr. (1, 2, 3), S. minuta Bréb. ex Kütz., nom. Illeg (1, 2, 3), S. neglecta Reich. (2), S. ovalis Bréb. (2, 3), S. suecica Grun. (1, 2, 3).
Класс Coscinodiscophyceae представлен лишь десятью видами (доля во флоре 7.5%), из четырех порядков. Несколько богаче других порядки Thalassiosirales (3.7%) и Aulacosirales (2.2%), участие во флоре остальных порядков незначительно.
Основу видового богатства диатомовых бентоса создавали классы Fragilariophyceae и Bacillariophyceae (123 видовых и внутривидовых таксона, 92.5%), относящиеся к девяти порядкам, 19 семействам и 41 роду. Первый сформирован порядком Fragilariales с одноименным семейством (11 родов, 24 вида), богаче других роды Staurosira (пять видов, 3.8%) и Punctastriata (три вида, 2.3%).
Класс Bacillariophyceae (99 таксонов, 74.4%) включал 18 семейств. Состав родов порядка Naviculales заметно разнообразнее других (12), скромнее – у Cymbellales (7) и Bacillariales (3). Большинство семейств этого класса содержало по 1–2, реже 3–4 вида, максимальным богатством отличались Bacillariaceae (19), Cymbellaceae (15) и Naviculaceae (11).
Таким образом, в пятерку доминирующих порядков в составе диатомовых входили Naviculales (27 видовых и внутривидовых таксонов), Cymbellales (26), Fragilariales (24), Bacillariales (19) и Achnanthales (11) (80.4% списка). Среди доминирующих семейств выделялись: Fragilariaceae (24), Bacillariaceae (19) Cymbellaceae (15), Naviculaceae (11), Gomphonemataceae (10) и Surirellaceae (7). Головная часть флоры диатомовых образована родами Nitzschia (13 видов), Gomphonema (10), Navicula (9), Surirella (7), Encyonema (6), Tryblionella, Staurosira, Planothidium (по 5), Gyrosigma и Cymbella (по 4).
Список диатомовых водорослей эпилитона оказался разнообразнее, чем эпифитона и эпипелона, а вклад основных таксономических групп на уровне классов был почти одинаковым. Сезонные изменения богатства бентосных диатомовые невелики, хотя число видов в пробе немного возрастало от июля (32.6 ± 2.6) к ноябрю (37.5 ± 2.0).
Комплекс диатомовых с частотой встречаемости во всех альгоценозах бентали ≥50% формировали 27 видов водорослей. В ≥70% проб отмечены Stephanodiscus hantzschii, S. neoastraea (100%), Cyclotella meneghiniana, Cocconeis placentula (96%), Navicula tripunctata, N. cryptocephala, Ulnaria ulna (88%), Gomphonema pumilum (80%), G. parvulum (76%), Tryblionella hungarica (72%) и Aulacosiera subractica (72%), встречаемость остальных заметно ниже.
Число видов, обнаруженных единично (например, Craticula cuspidаtа, Luticula permuticopsis, L. goeppertiana, Gomphonema supertergestinum и др.), достигало 40% общего числа определенных видов, причeм в эпилитоне оно было почти в два (43%), а их доля (37%) в 1.6 раза больше, чем в эпипелоне и эпифитоне.
Несмотря на высокую частоту встречаемости массовых видов (обилие от пяти до девяти баллов), частота их доминирования, для всех сообществ диатомовых оказалась ≤24–40%. Максимальные показатели отмечены для типичных планктеров – доминантов фитопланктона устья р. Окa Stephanodiscus neoastraea, S. hantzschii (Okhapkin et al., 2014). Большинство донных форм были массовыми в 4–12% проб и только четыре вида (Nitzschia dissipata, Navicula tripunctata, Gomphonema pumilum, Diatoma vulgaris) доминировали чаще (20–28%).
Встречаемость диатомовых незначительно, а частота доминирования достоверно возрастали от эпипелона к эпифитону (в 5 раз) и эпилитону (в 6 раз) (табл. 2). Таким образом, комплекс диатомовых водорослей эпилитона оказался флористически более насыщенным по максимальному общему (116) и удельному (38.5 ± 3.0) видовому богатству, частоте встречаемости и доминированию.
Таблица 2.
Основные характеристики видового богатства диатомовых водорослей
| Показатель | Эпипелон | Эпифитон | Эпилитон | Все альгоценозы |
|---|---|---|---|---|
| Общее видовое богатство | 95 | 94 | 116 | 135 |
| Число массовых видов | 26 | 24 | 27 | 27 |
| Удельное видовое богатство* | $\frac{{13 - 37}}{{29.6 \pm 2.9}}$ | $\frac{{30 - 39}}{{{\text{34}}{\text{.6}} \pm {\text{1}}{\text{.2}}}}$ | $\frac{{18 - 53}}{{38.5 \pm 3.0}}$ | $\frac{{13 - 53}}{{33.0 \pm 1.7}}$ |
| Частота встречаемости, % | 53.7 ± 5.9 | 56.1 ± 6.7 | 64.1 ± 4.8 | 59.3 ± 5.2 |
| Частота доминирования, % | 2.6 ± 1.5 | 13.1 ± 3.5 | 15.7 ± 2.7 | 10.8 ± 1.9 |
Ценозообразующий комплекс всех группировок диатомовых бентали формировали центрические диатомовые из планктонных сообществ р. Окa в результате их седиментации из толщи воды. Только Fragillaria vaucheriae в 14% проб эпипелона выступала в качестве доминанта. В эпифитоне преобладали Nitzschia palea, Cocconeis placentula, Ulnaria ulna, Gomphonema pumilum и G. parvulum (рис. 2), значительно реже – Navicula tripunctata, Diatoma vulgaris, Cocconeis pediculus, Nitzschia dissipata и Nitzschia sp. Кроме центрических диатомовых эпилитон формировали Navicula tripunctata, Nitzschia dissipata, Nitzschia sp. sp., Diatoma vulgaris и Achnanthidium affine, менее часто – Gomphonema parvulum, G. pumilum, Navicula capitatoradiata и Fragillaria vaucheriae.
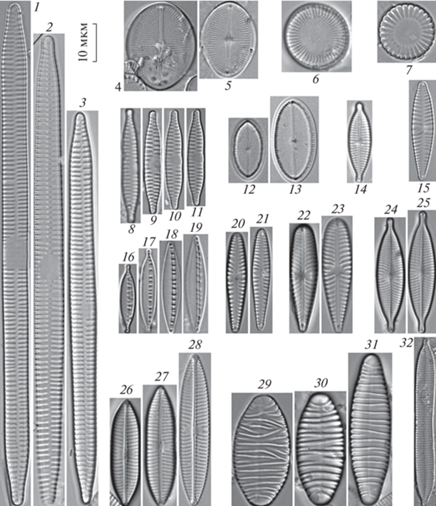
Рис. 2.
Доминирующие виды диатомовых водорослей. 1–3 – Ulnaria ulna, 4, 5 – Cocconeis pediculus, 6 – Stephanodiscus neoastraea, 7 – Cyclotella meneghiniana, 8–11 – Fragilaria vaucheriae, 12, 13 – Cocconeis placentula, 14 – Gomphonema parvulum, 15 – Nitzschia palea, 16–19 – Nitzschia dissipata, 20, 21 – Gomphonema pumilum, 22, 23 – Gomphonella olivacea, 24, 25 – Navicula capitatoradiata, 26–28 – Navicula tripunctata, 29–31 – Diatoma vulgaris Bory, 32 – Nitzschia constricta.
Анализ пространственно-временного распределения видовой структуры сообществ диатомовых водорослей с учетом их обилия методом кластерного анализа показал, что наиболее значимые факторы, определяющие особенности видового состава диатомовых бентоса, – сезон (июль, ноябрь) и приуроченность к определенному типу субстрата (рис. 3).
Рис. 3.
Дендрограмма иерархической кластеризации видового состава альгоценозов бентоса. По оси ординат – расстояние объединения. По оси абсцисс цифрами даны экологические группировки на станциях в разные сезоны. Июль: 1-я станция – 1 – эл (эпилитон), 2 – эп (эпипелон), 3–5 – эф (эпифитон); 2-я станция – 6, 9 – эл, 7 – эф, 8 – эп; 3-я станция – 10, 13 – эл, 11 – эп, 12, 14 – эф. Ноябрь: 1-я станция – 15, 18 – эл, 16, 17– эп, 19 – эф; 2-я станция – 20, 21 – эл, 22 – эп; 3-я станция – 23, 24 – эл, 25 – эп. Пунктирной линией выделены кластеры, показывающие сезонные различия и приуроченность к определенному типу субстрата: I – ноябрь, эл; II – июль, эл; III – ноябрь, эп; IV – июль, эф; V – июль, эп.

Среди диатомовых водорослей бентали реки поровну представлены планктонно-бентосные (46%) и бентосные (45%) виды, обитатели умеренно теплых (81% видов с известными из литературы температурными предпочтениями) нейтрально-олигощелочных вод с замедленным течением и умеренно насыщенных кислородом. Среди определенных диатомовых водорослей – главным образом типичные представители пресных вод, индифференты, устойчивые к органическому загрязнению, индикаторы незначительной и повышенной сапробности. Насыщенность вод р. Ока биогенными элементами и органическими веществами отразилась на преимущественном развитии мезо-эутрафентов и эутрафентов – видов, требующих для своей жизнедеятельности повышенного и высокого содержания органических форм азота, в том числе и миксотрофов.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В период наблюдения характеристики компонентного состава и уровень загрязнения окских вод, по сравнению с данными 2011 г., почти не изменились (табл. 1). По-прежнему низовья реки можно рассматривать как эвтрофно-гипертрофный водный объект с высокми уровнем антропогенного эвтрофирования и загрязнения (Охапкин и др., 2015; Джамалов и др., 2017) на фоне роста минерализации и содержания сульфатов и щелочных металлов, отмечаемого со второй половины XX в. к началу XXI в.
Несмотря на низкий класс качества вод, локальность изученного участка бентали и непродолжительность периода исследования, видовой состав диатомовых водорослей оказался весьма богатым (133 таксона рангом ниже рода) и сопоставимым с таковым донных и эпифитных представителей Bacillariophyta (137 таксонов), зарегистрированных ранее в фитопланктоне реки (Охапкин, Хедаириа, 2019). Этот показатель незначительно отичался от отмеченного в нижнем течении р. Днепр (Владимирова, 1978), но был в 1.7 раза ниже такового в устье р. Дунай (Кoziychuk, 2019) и в 1.3 раза – выявленного в нижнем течении р. Енисей (Левадная, 1986) при несопоставимо большем объеме изученного материала.
Заметная роль семейств Naviculaceae и Nitzschiaceae и представителей родов Nitzschia, Navicula, Gomphonema, Achnanthidium в альгоценозах бентоса р. Ока характерна была ранее и для водоемов и водотоков Верхней Волги, рек Днепр, Дунай и других крупных эвтрофированных водных объектов умеренной зоны (Владимирова, 1978; Волга…, 1978; Potapova, Chrles, 2002; Кoziychuk, 2019). Часть отмеченных диатомей (Achnanthes, Cocconeis, Gomphonema, Ulnaria и др.) имеет широкий географический ареал, развиваясь в РФ от водоемов субарктической тундры до водотоков Дальнего Востока (Медведева, Семенченко, 2019; Копырина и др., 2021). Отсутствие в перечне ведущих родов Pinnularia и Eunotia, характерных для горных рек с водами незначительной минерализации, богатых железом, и рек с заболоченным и залесенным водосбором (Diatomeen…, 2013; Krizmaniĉ et al., 2015), отражает нейтрально-олигощелочной характер окских вод с повышенным содержанием солей, богатых доступными формами азота и фосфора.
Соотношение часто встречающихся и редких (обнаруженных в одной–двух пробах видов) донных диатомей (первых – 20%, вторых >70% общего состава) аналогично таковому для альгофлоры планктона рек Ока и Волга (Охапкин, 1994). Примерно такое же соотношение, как в р. Окa (21.6%), установлено и для р. Монйолинка в Бразилии (Bere, 2010).
Состав и обилие доминирующих диатомовых в бентосе связаны с развитием фитопланктона, водоросли из которого, постоянно оседая на илистом грунте, определяют специфику микрофитобентоса. Это явление для рипали р. Волга известно еще из работы Н.В. Кордэ, 1950 (цит. по: Волга…, 1978) и отмечалось неоднократно другими авторами (Potapova, Charles, 2002; Комулайнен, 2004; Метелева, 2013). Седиментация фитопланктона активно происходила и на поверхности водных растений, но состав бентосно-перифитонных видов и их встречаемость в сообществах эпифитона заметнее, чем в ценозах эпипелона. Более чем в 2 раза разница между минимальными и максимальными значениями удельного видового богатства водорослей эпифитона по сравнению с аналогичным показателем неживых субстратов, возможно, свидетельствует о средообразующем влиянии растения-форофита, наличии конкурентных отношений и формировании более стабильной среды обитания для диатомовых водорослей. Богатый видовой состав эпилитона с достаточно высоким или заметным доминированием, по-видимому, определяется большей пространственной неоднородностью абиотических условий. Приуроченность ряда видов диатомовых к одному типу субстрата, возможно, является следствием значительно большей их стенобионтности, однако для выявления характера и механизмов этой приуроченности нужны специальные исследования.
Оценка условий среды обитания по показательным свойствам бентосных водорослей в основном соответствовала химизму окских вод, кроме индикации загрязнения органическими веществами. Преобладание обитателей чистых вод (60% числа индикаторов сапробности) и незначительная доля высокосапробных компонентов может быть связано с заниженными значениями индивидуальных характеристик многих видов, приведенных в работе (Баринова и др., 2019). Это еще раз подтверждает необходимость исследований по региональной адаптации показательных значений отдельных видов. Возможно, такой результат является следствием химического состава органического вещества грунтов, что требует специальных исследований.
Выводы. Видовой состав диатомовых водорослей альгоценозов бентоса р. Ока довольно богат (133 вида и внутривидовых таксона) и достаточно типичен для крупных эвтрофированных и загрязненных рек умеренной зоны. Преобладание пеннатных шовных диатомовых, ранговое распределение ведущих семейств и родов свойственно составу диатомовых бентоса крупных речных экосистем с заметным антропогенным влиянием. Сообщества диатомовых эпилитона по всем изученным флористическим характеристикам оказались сложнее организованными, чем таковые эпифитона и эпипелона. На структуру видового богатства прибрежных бентосных альгоценозов большое влияние оказывает фитопланктон, доминирующие компоненты которого встречены на всех субстратах. Массовые виды, отмеченные в >50% проб, составляли лишь 20% видового состава, редкие, найденные единично – 40%. Состав доминирующих видов (встречаемость ≥50%, доминирование 20–40%) небогат (шесть видов). Низкая частота доминирования большинства обычных компонентов бентосных альгоценозов косвенно свидетельствует о пространственно-временном разнообразии условий обитания диатомовых в бентали р. Ока с преобладанием субстратной и сезонной приуроченности. Оценка экологического состояния рипали устьевого района р. Ока по составу диатомовых водорослей подтвердила их высокие индикационные свойства. Отмечено ее соответствие химизму вод, кроме характеристики органического загрязнения по числу индикаторов сапробности.
Список литературы
Баринова С.С., Бeлоус Е.П., Царенко П.М. 2019. Альгоиндикация водных объектов Украины: методы и перспективы. Хайфа. Киев: Изд-во ун-та Хайфы.
Владимирова К.С. 1978. Фитомикробентос Днепра, его водохранилищ и Днепровско-Бугского лимана. Киев: Наук. думка.
Водоросли. Справочник. 1989. Киев: Наук. думка.
Волга и ее жизнь. 1978. Ленинград: Наука.
Джамалов Р.Г., Никаноров А.М., Решетняк О.С., Сафронова Т.И. 2017. Воды бассейна Оки: химический состав и источники загрязнения // Вода и экология. № 3. С. 114.
Комулайнен С.Ф. 2003. Методические рекомендации по изучению фитоперифитона в малых реках. Петрозаводск: Карельск. науч. центр РАН.
Комулайнен С.Ф. 2004. Фитоперифитон рек Республики Карелия // Ботан. журн. Т. 89. № 3. С. 18.
Копырина Л.И., Генкал С.И., Ремигайло П.А. 2021. Диатомовые водоросли водоемов субарктической тундры // Биология внутр. вод. № 2. С. 107. https://doi.org/10.31857/S0320965221020091
Куликовский М.С., Глущенко А.М., Генкал С.И., Кузнецова И.В. 2016. Определитель диатомовых водорослей России. Ярославль: Филигрань.
Левадная Г.Д. 1986. Микрофитобентос реки Енисей. Новосибирск: Наука.
Медведева Л.А., Семенченко А.А. 2019. Структурные и количественные особенности сообществ фитоперифитона в водотоках бассейна реки Зея (Амурская область) // Биология внутренних вод. № 1. С. 23. https://doi.org/10.1134/S0320965219010145
Метелева Н.Ю. 2013. Структура и продуктивность фитоперифитона водоемов бассейна Верхней Волги: Автореф. дис. … канд. биол. наук. Борок.
Метелева Н.Ю., Девяткин В.Г. 2005. Формирование и продуктивность перифитона Рыбинского водохранилища: состав и обилие // Биология внутр. вод. № 2. С. 56.
Насибулина Б.М., Тлекова Р.С. 2015. Динамика речных биоресурсов в сезонном аспекте в водоемах дельты р. Волги // Естественные науки. № 2. С. 42.
Неврова Е.Л., Снигирева А.А., Петров А.Н., Ковалева Г.В. 2015. Руководство по изучению морского микрофитобентоса и его применению для контроля качества среды. Севастополь; Симферополь: Н. Орiанда.
Охапкин А.Г. 1994. Фитопланктон Чебоксарского водохранилища. Cамара: СамВен.
Охапкин А.Г., Андриянова Н.В., Максимова В.А. и др. 2015. Динамика гидрохимического состава вод нижнего течения р. Оки // Вода: химия и экология. № 5. С. 15.
Охапкин А.Г., Хедаириа Т. 2019. Диатомовые водоросли как компонент донных альгоценозов устьевого участка реки Оки // Вопросы современной альгологии. № 2 (20). С. 60. URL: http://algology.ru/1494.
Семенченко В.П. 2004. Принципы и методы биоиндикации текучих вод. Минск: Изд-во “Орех”.
Якимов В.Н. Шурганова Г.В., Черепенников В.В. и др. 2016. Методы сравнительной оценки результатов кластерного анализа структуры гидробиоценозов (на примере зоопланктона реки Линда Нижегородской области) // Биология внутр. вод. № 2. С. 94. https://doi.org/10.7868/S0320965216020169
Bere T. 2010. Benthic diatom community structure and habitat preferences along an urban pollution gradient in the Monjolinho River, São Carlos, SP, Brazil // Acta Limnol. Brasiliensia. V. 22. № 1. P. 80.
Bere T., Tundisi J.G. 2010. Biological monitoring of lotic ecosystems: the role of diatoms // Brazilian J. Biology. V. 70. P. 493.
Coste M., Boutry S., Tison-Rosebery J., Delamas F. 2009. Improvements of the Biological Diatom Index (BDI): Description and efficiency of the new version (BDI-2006) // Ecological Indicators. V. 9. P. 621.
Diatomeen im süßwasser-benthos von Mitteleuropa. 2013. Kanigstein: Koeltz Scientific Books.
Kelly M., Bennion H., Burgess A. et al. 2009. Uncertainty in ecological status assessments of lakes and rivers using diatoms // Hydrobiologia. V. 633. P. 5.
Koziychuk E.Sh. 2019. Phytomicrobenthos Diversity in Watercourses of the Killya Delta of the Danube River // Hydrobiol. J. V. 55. № 5. P. 44.
Krammer K., Lange-Bertalot H. 1986. Bacillariophyceae. Teil 1: Naviculaceae // Susswasserflora von Mitteleuropa. Jena: Gustav Fisher Verlag. Bd 2/1.
Krammer K., Lange-Bertalot H. 1988. Bacillariophyceae. Teil 2: Bacillariaceae, Epithemiaceae, Surirellaceae // Susswasserflora von Mitteleuropa. Jena: Gustav Fisher Verlag. Bd 2/2.
Krammer K., Lange-Bertalot H. 1991a. Bacillariophyceae. Teil 3: Centrales, Fragilariaceae, Eunotiaceae // Susswasserflora von Mitteleuropa. Jena: Gustav Fisher Verlag. Bd 2/3.
Krammer K., Lange-Bertalot H. 1991б. Bacillariophyceae. Teil 1–4: Achnanthaceae, Kritische Erganzungen zu Navicula (Lineolatae) und Gomphonema Gesamtliteraturverzeichnis // Susswasserflora von Mitteleuropa. Jena: Gustav Fisher Verlag. Bd 2/4.
Krizmaniĉ J., Iliĉ M., Vidakoviĉ D. et al. 2015. Diatoms of the Dojkinci River (Stara Planina Nature Park, Serbia) // Acta bot. Croat. V. 74. № 2. P. 317.
Okhapkin A.G., Genkal S.I., Sharagina E.M., Vodeneeva E.L. 2014. Structure and dynamics of phytoplankton in the Oka river mouth at the beginning of the 21th century // Inland Water Biology. V. 7. № 4. P. 357. https://doi.org/10.1134/S1995082914040142
Potapova M.G., Charles D.F. 2002. Benthic diatoms in USA rivers: distributions along spatial and environmental gradients // J. Biogeography. V. 29. P. 167.
Round F.E., Crawford R.M., Mann D.G. 1990. The Diatoms: biology and morphology of the genera. Cambridge: Cambridge University Press.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод