

Биология внутренних вод, 2023, № 2, стр. 152-163
Amphimonhystrella tropica sp. n. и Oncholaimus mekongis sp. n. (Nematoda) р. Меконг, Вьетнам
В. Г. Гагарин a, *, В. А. Гусаков a
a Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл., Россия
* E-mail: gagarin@ibiw.ru
Поступила в редакцию 01.04.2022
После доработки 24.05.2022
Принята к публикации 02.06.2022
- EDN: GMHYZB
- DOI: 10.31857/S0320965223020055
Аннотация
Приведено иллюстрированное описание двух новых видов свободноживущих нематод: Amphimonhystrella tropica sp. n. и Oncholaimus mekongis sp. n., обнаруженных в устье р. Меконг во Вьетнаме. Amphimonhystrella tropica по размерам тела близка к A. major Gagarin, Nguyen Dinh Tu, 2021 и A. paramonovi (Phan Ke Long, Nguyen Dinh Tu, Gagarin, 2020) comb. nov. От первого вида отличается формой фарингостомы, более толстым телом, более длинным хвостом, отсутствием заднего мешка матки и более длинными спикулами, от второго вида – более толстым телом, относительно более длинными спикулами и иной структурой рулька. Вид Daptonema paramonovi Phan Ke Long, Nguyen Dinh Tu, Gagarin, 2020 переводится в род Amphimonhystrella: A. paramonovi (Phan Ke Long, Nguyen Dinh Tu, Gagarin, 2020) comb. nov. Приведены дихотомический и рисуночный ключи для определения самцов рода Amphimonhystrella. Oncholaimus mekongis sp. n. по размерам тела и длине спикул близок к O. ball Kreis, 1934 и O. menzeli Schneider, 1973. От обоих видов отличается наличием крупной шиповидной щетинки перед клоакой у самцов, расположением вентральной папиллы на хвосте у самцов, более толстым телом, более короткими внешними губными щетинками и более длинным и стройным хвостом у самцов.
ВВЕДЕНИЕ
Свободноживущие круглые черви (Nematoda) – одна из наиболее разнообразных и многочисленных современных групп беспозвоночных животных, чьи представители успешно освоили всевозможные биотопы различных континентальных водоемов от промерзающих полярных озер до горячих минеральных источников. Плотность популяций нематод достигает ~1 × 106 экз./м3, но может быть и выше. Средняя биомасса доходит до 2 г/м2 сырой массы или 0.2–0.5 г углерода/м2 (Wieser, Kanwisher, 1960). Благодаря своему интенсивному метаболизму, нематоды с биомассой ~4% биомассы макрофауны создают до 30% продукции всего бентоса (Gerlach, 1978). Виды родов Amphimonhystrella и Oncholaimus – обычные донные обитатели солоноватых и морских водоемов (Gerlachi, Riemann, 1973, 1974).
Цель работы – описать новые виды из указанных родов, обнаруженные в процессе исследования донной мейофауны в дельте р. Меконг во Вьетнаме.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материал, в котором обнаружены новые виды нематод, собран в начале декабря 2019 г. в прибрежной и русловой зонах рукава Котьен (Cổ Chiên) дельты р. Меконг. Точные координаты и дополнительная информация о местах сбора указаны в соответствующих описаниях видов.
Пробы донных отложений собирали с помощью микробентометра С-1 с диаметром трубки 34 мм (~9 см2). В каждой точке отбирали по три пробы грунта и придонной воды (по 5–10 см каждого слоя), которые объединяли в одну интегральную пробу и фиксировали 4%-ным формалином. В лаборатории пробы фильтровали через сито с размером ячеи 82 × 82 мкм. Оставшееся на сите содержимое окрашивали красителем Бенгальский розовый по методике (Williams D.D., Williams N.E., 1974) и далее просматривали в камере Богорова под бинокулярным микроскопом. Обнаруженных червей извлекали с помощью пипетки и монтировали на глицериновые препараты на предметные стекла. Определение, измерение и фотографирование особей проводили с использованием светового микроскопа Nikon Eclipse 80i, оборудованного принадлежностями для наблюдения методом ДИК-контраста, цифровой камерой Nikon DS-Fi1 и ПК с программой NIS-Elements D 3.2 для анализа и документирования изображений с препаратов.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Описание новых видов. Отряд Monhysterida Filipjev, 1929. Семейство Xyalidae Chitwood, 1951.
Род Amphimonhystrella Timm, 1961. Диагноз (по: (Lorenzen, 1977), с изменениями). Длина тела от 380 до 800 мкм. Кутикула кольчатая. Шесть внутренних губных сенсилл в форме папилл. Шесть внешних губных сенсилл и четыре головные сенсиллы в форме тонких щетинок. Фовеи амфидов в форме круга, расположены сравнительно далеко от переднего конца тела. Фарингостома крупная, удлиненная, с сильно кутикулизированными стенками. Задний мешок матки у большинства видов имеется. Семенники парные. Передний семенник расположен слева от кишки, задний – справа. Спикулы сильно изогнуты вентрально. Рулек, как правило, имеется и у большинства видов с дорсальным отростком. Хвост удлиненно-конический, с терминальными щетинками.
Типовой вид: A. megastoma Timm, 1961
Amphimonhystrella tropica sp. n. (рис. 1, рис. 2; табл. 1).
Рис. 1.
Amphimonhystrella tropica sp. n.: а – голова самца; б – область вульвы; в – передний конец самца; г – область клоаки; д – хвост самки; е – хвост самца.
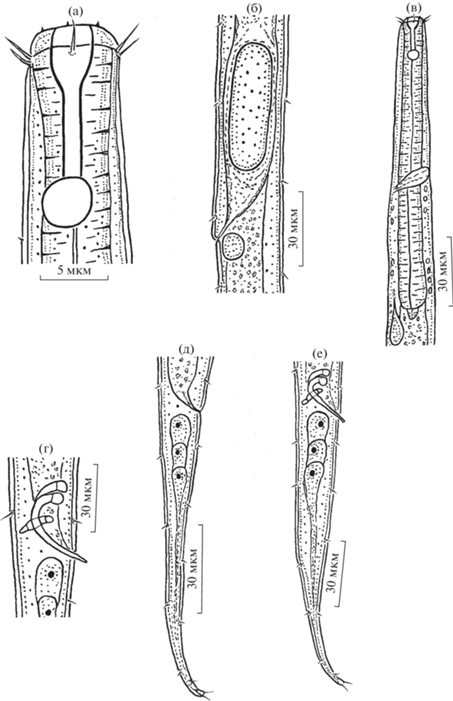
Рис. 2.
Самец (а, в, г, е, ж, и, к) и самка (б, д, з, л) Amphimonhystrella tropica sp. n.: а, б – общий вид; в, г, д – голова; е – передний конец тела; ж – область кардия; з – область вульвы; и – область клоаки; к, л – хвост. an – анус, ca – кардий, ce – целомит, cl – клоака, f.a – фовея амфида; gub – рулек; in – кишка; o.l.s – внешние губные щетинки; ph – фаринкс; phst – фарингостома; p.v.c – поствульварная клетка; re – ренетта; spic – cпикула; t – хвост; t.s – терминальная щетинка; v – вульва.

Таблица 1.
Морфометрическая характеристика Amphimonhystrella tropica sp. n.
| Признак | Голотип ♂ | Паратипы | ||
|---|---|---|---|---|
| 1♂ | 7♀♀ | |||
| min–max | среднее | |||
| L, мкм | 724 | 655 | 622–726 | 689 |
| a | 20 | 30 | 23–29 | 27 |
| b | 5.2 | 5.0 | 5.0–5.7 | 5.3 |
| c | 6.3 | 6.0 | 5.9–6.8 | 6.5 |
| c' | 4.6 | 5.5 | 6.1–7.1 | 6.8 |
| V, % | – | – | 56–61 | 59 |
| Ширина области губ, мкм | 6.1 | 6.3 | 5.1–6.9 | 5.9 |
| Ширина тела, мкм: | ||||
| в его среднем отделе | 36 | 22 | 22–30 | 26 |
| в области клоаки или ануса | 25 | 20 | 15–17 | 16 |
| Длина, мкм: | ||||
| внешних губных щетинок | 2.5 | 3.0 | 2.6–3.1 | 2.8 |
| стомы | 11.2 | 10.1 | 8.2–11.0 | 9.6 |
| фаринкса | 139 | 130 | 115–137 | 129 |
| хвоста | 115 | 110 | 92–117 | 107 |
| спикул (по дуге) | 41 | 37 | – | – |
| Расстояние, мкм: | ||||
| от переднего конца тела до фовеи амфидов | 10.4 | 8.9 | 7.2–11.0 | 9.3 |
| от заднего конца фаринкса до вульвы | – | – | 242–306 | 277 |
| от заднего конца фаринкса до клоаки | 470 | 415 | – | – |
| от вульвы до ануса | – | – | 148–187 | 176 |
Материал. Голотип ♂ (инвентарный номер препарата М23); паратипы: 1♀ (инвентарный номер препарата М20), 1♀ (инвентарный номер препарата М27), 3♀♀ (инвентарный номер препарата М32), 1♂ и 2♀♀ (инвентарный номер препарата М38). Препараты голотипа и паратипов хранятся в коллекции нематод из р. Меконг в лаборатории экологии водных беспозвоночных Института биологии внутренних вод РАН (пос. Борок, Ярославская обл., Россия).
Типовое местонахождение. Вьетнам, дельта р. Меконг, рукав Котьен (Cổ Chiên), центральная часть русла. Координаты станции: 09°59.503′ с.ш., 106°20.541′ в.д., глубина 16.0 м, грунт – серая плотная глина, соленость воды у дна 17.7 г/л. Сбор 05.12.2019 г.
Дополнительные местонахождения. Особи паратипов обнаружены в рукаве Котьен в следующих точках: 1. прибрежная часть русла, координаты: 09°59.433′ с.ш., 106°20.394′ в.д., глубина 3.5 м, грунт – серая заиленная глина с песком, соленость воды у дна 1.3 г/л, сбор 05.12.2019 г.; 2. центральная часть русла, координаты: 09°52.547′ с.ш., 106°31.773′ в.д., глубина 12.5 м, грунт – песок с серо-коричневым глинистым илом, соленость воды у дна 28.8 г/л, сбор 06.12.2019 г.; 3. прибрежная часть русла, координаты: 09°52.622′ с.ш., 106°31.965′ в.д., глубина 2.5 м, грунт – плотный песок с серой глиной, соленость воды у дна 12.7 г/л, сбор 06.12.2019 г.; 4. прибрежная часть русла, координаты: 09°56.529′ с.ш., 106°27.291′ в.д., глубина 4.0 м, грунт – плотный песок с серой глиной, соленость воды у дна 5.5 г/л, сбор 06.12.2019 г.
Описание. Морфометрическая характеристика голотипа и паратипов приведена в табл. 1.
Самцы. Мелкие, стройные черви. Передний конец тела сужен. Кутикула мелкокольчатая, тонкая. Толщина кутикулы в среднем отделе тела ~1 мкм. Соматические щетинки короткие и редкие. Шесть внутренних губных сенсилл в форме папилл. Шесть внешних губных сенсилл и четыре головные сенсиллы в форме тонких щетинок. Длина внешних губных щетинок 2.5–3.0 мкм, что равно 41–48% ширины области губ. Головные щетинки длиной ~1.5 мкм. Внешние губные щетинки и головные щетинки расположены близко друг к другу, образуя единый общий круг. Хейлостома сравнительно маленькая. Фарингостома длинная и разделена на два отдела: передний – широкий и короткий, задний – длинный и узкий. Стенки фарингостомы сильно кутикулизированы. Фовеи амфидов в форме круга диаметром 4.2–4.9 мкм, расположены на расстоянии 8.9–10.4 мкм от переднего конца тела. Фаринкс мускулистый, только слегка расширяется к своему основанию. Кардий маленький, вдается в просвет средней кишки. В полости тела, на уровне задней половины фаринкса имеются многочисленные мелкие целомиты. Небольшая мешковидная ренетта расположена на уровне начала средней кишки.
Семенников два, противопоставленные. Передний семенник расположен слева от кишки, задний – справа. Спикулы тонкие, с головками. Проксимальные концы спикул сильно изогнуты вентрально. Длина спикул 37–41 мкм, что в 1.8–1.9 раза больше диаметра тела в области клоаки. Рулек маленький, с двумя дорсальными отростками. Хвост удлиненно-конический, постепенно сужается. Три каудальные железы плохо заметны. Спиннерета короткая. Субтермальные щетинки на хвосте длиной 2.0–2.5 мкм.
Самки. По общей морфологии подобны самцам. Строение кутикулы и переднего конца тела как у самцов. Кутикула тонкокольчатая. Шесть внутренних губных сенсилл в форме папилл. Шесть внешних губных сенсилл и четыре головные сенсиллы в форме тонких щетинок. Длина внешних губных щетинок 1.8–2.0 мкм, длина головных щетинок около 1.5 мкм. Внешние губные щетинки и головные щетинки расположены в единый круг. Фарингостома крупная, разделена на два отдела: передний отдел широкий и короткий, задний отдел более длинный и узкий. Стенки фарингостомы сильно кутикулизированы. Длина общей стомы 8.2–11.0 мкм, что в 1.5–1.7 раза превышает ширину области губ. Фаринкс мускулистый, слегка расширяется к своему основанию. Кардий маленький, вдается в просвет средней кишки. В полости тела на уровне заднего конца фаринкса имеются многочисленные мелкие целомиты. Ренетта расположена на уровне начала средней кишки. Длина ректума равна или немного меньше диаметра тела в области ануса.
Яичник один, передний, прямой, сравнительно длинный, расположен слева от средней кишки. Вульва постэкваториальная, в форме продольной щели. Губы вульвы не склеротизированы и не выступают за контуры тела. Вагина сравнительно короткая и наклонена к переднему концу тела. Матка обширная, заполнена многочисленными сперматозоидами. У одной самки в матке обнаружено одно яйцо размером 51 × 26 мкм. Задний мешок матки отсутствует. Однако у всех самок наблюдается поствульварная округлая клетка. Хвост удлиненно-конический, постепенно сужается. Терминальные щетинки на хвосте длиной 2.0–2.5 мкм.
Дифференциальный диагноз. A. tropica sp. n. по размерам тела близка к видам А. major Gagarin, Nguyen Dinh Tu, 2021 и A. paramonovi (Phan Ke Long, Nguyen Dinh Tu, Gagarin, 2020) comb. nov. (табл. 2). От первого вида отличается формой фарингостомы (у нового вида фарингостома разделена на два отдела, у А. major фарингостома единая, цельная, слегка сужена к заднему концу); менее стройным телом (а = 20–30 против а = 32–38 у А. major); более длинным хвостом (с = = 5.9–6.8 против с = 8.0–8.5 у А. major); отсутствием заднего мешка матки (у самок А. major этот мешок имеется); более длинными спикулами (длина спикул 37–41 мкм против 25–26 мкм у А. major) и формой спикул (Gagarin, Nguyen Dinh Tu, 2021). От второго вида отличается более толстым телом (а = 20–30 против а = 30–40 у A. paramonovi), более длинными спикулами (длина спикул 37–41 мкм, против 22–26 мкм у A. paramonovi) и структурой рулька (у нового вида рулек с двумя дорсальными отростками, у A. paramonovi рулек без дорсальных отростков и в форме тонкого желоба) (Фан Ке Лонг и др., 2020).
Таблица 2.
Основные морфометрические признаки самцов валидных видов рода Amphimonhystrella
| Признак | A. bullacauda | A. megastoma | A. parva | A. unita | A. major | A. paramonovi | A. tropica sp. n. |
|---|---|---|---|---|---|---|---|
| L, мкм: | 443–487 | 524 | 388–447 | 485 | 740–797 | 519–609 | 724, 655 |
| a | 28–35 | 28 | 32–47 | 36 | 32–38 | 30–40 | 20, 30 |
| b | 4.0–4.4 | 5.2 | 4.1–4.7 | 4.9 | 5.7–6.2 | 5.1–6.2 | 5.2, 5.0 |
| c | 6.7–7.4 | 11.0? | 4.7–6.0 | 6.9 | 8.0–8.5 | 6.4–7.6 | 6.3, 6.0 |
| c' | 5.1–6.5 | 3.0? | 7.3–8.7 | 6.0 | 5.4–5.8 | 5.0–6.5 | 4.6, 5.5 |
| Длина, мкм: | |||||||
| стомы | 5.0–6.0 | 20 | 4.0–4.5 | ~6.0 | 12.5–13.0 | 10.0–10.8 | 11.2, 10.1 |
| внешних губных щетинок | 2.5–3.5 | 8.0 | 2.0–3.0 | ~1.5 | 4.0–4.5 | 2.0–2.5 | 2.5, 3.0 |
| спикулы (по дуге) | 14.0–15.5 | 20 | 15–17 | 16 | 25–26 | 22–26 | 41, 37 |
| Наличие и структура рулька | Имеется с дорсальным отростком | Отсутствует | Имеется с дорсальным отростком | Имеется с дорсальным отростком | Имеется с дорсальным отростком | Имеется без дорсального отростка | Имеется с дорсальным отростком |
| Отношение длины внешних губных щетинок к ширине области губ, % | 65–75 | ~100 | 45–55 | 35–45 | 65–70 | 35–45 | 41, 48 |
| Примечание. Значения признаков указаны согласно числу промерянных особей нематод. У вида A. unita и A. megastoma промерен только 1 самец. У нового вида A. tropica sp.n. промерены 2 самца (голотип и паратип, см. табл. 1), значения указаны через запятую. У остальных видов измерены более 3 особей самцов, указаны минимальное и максимальное значение признака. | |||||||
Этимология. Видовое название означает “тропический”, “из тропиков”.
Морфологические и таксономические замечания. Род Amphimonhystrella Timm, 1961 был обоснован для вида A. megastoma Timm, 1961 (Timm, 1961). Род морфологически очень близок к роду Daptonema Cobb, 1920 и отличается от него только по одному признаку: формой фарингостомы. У нового рода фарингостома крупная, длинная и сильно кутикулизирована, у видов рода Daptonema фарингостома маленькая, в форме мелкой воронки, ее стенки слабо кутикулизированы (Fonseca, Bezerra, 2014).
В состав рода Amphimonhystrella в настоящее время входят cемь видов. Вид A. megastoma Timm, 1961 описан по единственному самцу из прибрежной мелководной зоны Бенгальского залива (Timm, 1961). Вид характеризуется довольно крупной стомой, длинными внешними губными щетинками и отсутствием рулька в спикулярном аппарате самца (табл. 2; рис. 3). Вид A. unita Lorenzen, 1977 описан по одной самке и одному самцу из пролива Скагерак (Scagerak Strait) (Lorenzen, 1977). Имеет довольно маленькие размеры, короткие внешние губные щетинки, короткие спикулы (длиной 16 мкм) и очень длинный задний мешок матки. Вид A. bullacauda Tchesunov, Milyutina, 2005 найден в прибрежной зоне Вьетнама. Имеет довольно короткое и толстое тело и сравнительно маленькую стому (рис. 3, табл. 2). Вид A. parva Gagarin, Nguyen Vu Thanh, 2009 описан из мангровых зарослей в дельте р. Меконг (Вьетнам). Характеризуется сравнительно малыми размерами тела, сравнительно далеко от переднего конца тела расположенными фовеями амфид и короткими спикулами. Вид А. major Gagarin, Nguyen Tu, 2021 обнаружен в мангровых зарослях устья р. Ук (Вьетнам). Имеет сравнительно короткий хвост и сравнительно длинные внешние губные щетинки (рис. 3, табл. 2). Вид A. paramonovi (Phan Ke Long, Nguyen Dinh Tu, Gagarin, 2020) comb. nov. описан из мангровых зарослей в дельте р. Иэн (Вьетнам) как Daptonema paramonovi Phan Ke Long, Nguyen Dinh Tu, Gagarin, 2020 (Фан Ке Лонг и др., 2020). Поскольку нематоды данного вида имеют довольно крупную фарингостому с сильно кутикулизированными стенками, мы переводим A. paramonovi в род Amphimonhystrella Timm, 1961: A. paramonovi (Phan Ke Long, Nguyen Dinh Tu, Gagarin, 2020) comb. nov. Нематоды этого вида имеют сравнительно длинную фарингостому, разделенную на два отдела, и своеобразное строение рулька (в форме узкого желоба, лишенного дорсального отростка (рис. 3)). Вид A. tropica Gagarin, Gusakov sp. n. найден в устье р. Меконг (Вьетнам), имеет характерное строение фарингостомы (разделена на два отдела), довольно длинное и толстое тело и крупные спикулы (их длина 37–41 мкм) (рис. 1, 2; табл. 1).
Рис. 3.
Рисуночный ключ для определения самцов видов рода Amphimonhystrella Timm, 1961: а – A. megastoma Timm, 1961 (пo: Timm, 1961); б – A. bullacauda Tchesunov, Miljutina, 2005 (пo: Tchesunov, Miljutina, 2005); в – A. parva Gagarin, Nguyen Vu Thanh, 2009 (по: Гагарин, Нгуен Ву Тхань, 2009); г – A. unita Lorenzen, 1977 (пo: Lorenzen, 1977); д – A. major Gagarin, Nguyen Dinh Tu, 2021 (пo: Gagarin, Nguyen Dinh Tu, 2021); е – A. paramonovi (Phan Ke Long, Nguyen Dinh Tu, Gagarin, 2020) comb. nov. (по: Фан Ке Лонг, Нгуен Динь Ты, Гагарин, 2020); ж – A. tropica Gagarin, Gusakov sp. n. (по: Gagarin, Gusakov, настоящая статья).
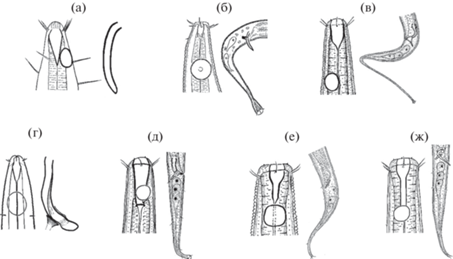
Ниже приведены рисуночный и дихотомический ключи для определения самцов видов рода Amphimonhystrella.
Дихотомический ключ для определения самцов видов рода Amphimonhystrella Timm, 1961
| 1. Длина внешних губных щетинок 8 мкм; рулек отсутствует | A. megastoma |
| – Длина внешних губных щетинок <8 мкм; рулек имеется | 2 |
| 2. Рулек без дорсального отростка | A. paramonovi |
| – Рулек с дорсальным отростком | 3 |
| 3. Фарингостома разделена на два отдела; длина спикуд 37–41 мкм | A. tropica sp. n. |
| – Фарингостома единая, цельная; длина спикул <30 мкм | 4 |
| 4. Длина тела 740–797 мкм; длина внешних губных щетинок 4.0–4.5 мкм | A. major |
| – Длина тела <500 мкм; длина внешних губных щетинок <4.0 мкм | 5 |
| 5. Длина стомы 4.0-4.5 мкм; с = 4.7–6.0 мкм | A. parva |
| – Длина тела 5 мкм и более; с более | 6 |
| 6. Длина стомы 5.0 мкм; длина внешних губных щетинок 1.5 мкм | A. unita |
| – Длина стомы 5.0–6.0 мкм; длина внешних губных щетинок 2.3–3.5 мкм | A. bullacauda |
Отряд Enoplida Filipjev, 1929. Сем. Oncholaimidae Filipjev, 1916
Род Oncholaimus Dujardin, 1845. Диагноз (по: Smol, Coomans, 2005). Левый субвентральный зуб крупнее правого субвентрального зуба и дорсального зуба. Самки монодельфные, продельфные с антидромным загнутым яичником. Система de Mana хорошо развита, терминальные протоки и поры присутствуют в различных вариантах и отсутствуют у неполовозрелых самок. Самцы диорхные. Спикулы короткие, рулек отсутствует. Хвост короткий.
Типовой вид: O. attenuatus Dujardin, 1845
Oncholaimus mekongis sp. n. (рис. 4, рис. 5; табл. 3).
Рис. 4.
Oncholaimus mekongis sp. n.: а – голова самца; б – область вульвы; в – передний конец самца; г – хвост самца; д – хвост самки; е – область клоаки.

Рис. 5.
Самец (а, в, г, ж, з, и) и самка (б, д, е, к) Oncholaimus mekongis sp. n.: а, б – общий вид; в – передний конец тела; г, д – голова; е – область вульвы; ж, з – область клоаки; и, к – хвост. an – анус, c.p – каудальная папилла; ca – кардий, cl – клоака, d.t – дорсальный зуб, in – средняя кишка, l.s.t – левый субвентральный зуб, ph – фаринкс; phst – фарингостома; pr.s – преклоакальная щетинка, r.s.t – правый субвентральный зуб, sp – спиннерета, spic – cпикула; t – хвост; v – вульва.

Таблица 3.
Морфометрическая характеристика Oncholaimus mekongis sp. n.
| Признак | Голотип♂ | Паратипы | ||
|---|---|---|---|---|
| 4♂♂ | 1♀ | |||
| min–max | среднее | |||
| L, мкм | 2253 | 1913–2173 | 2051 | 2318 |
| a | 26 | 23–29 | 27 | 29 |
| b | 5.7 | 5.4–5.8 | 5.6 | 6.0 |
| c | 24.8 | 22.2–27.5 | 24.5 | 30.9 |
| c' | 3.3 | 2.8–3.1 | 3.0 | 2.4 |
| V, % | – | – | – | 81.5 |
| Ширина области губ, мкм | 22 | 20–25 | 22 | 24 |
| Ширина стомы, мкм | 15 | 14–20 | 16 | 17 |
| Ширина тела, мкм: | ||||
| в его среднем отделе | 87 | 66–90 | 78 | 81 |
| в области клоаки или ануса | 28 | 26–31 | 29 | 31 |
| Длина, мкм: | ||||
| внешних губных щетинок | 4.7 | 4.3–4.6 | 4.4 | 4.4 |
| стомы | 28 | 27–29 | 29 | 30 |
| фаринкса | 398 | 357–378 | 368 | 386 |
| хвоста | 91 | 79–90 | 84 | 75 |
| спикул (по дуге) | 29 | 31–33 | 32 | – |
| Расстояние, мкм: | ||||
| от заднего конца фаринкса до вульвы | – | – | – | 1504 |
| от заднего конца фаринкса до клоаки | 1764 | 1470–1722 | 1549 | – |
| от вульвы до ануса | – | – | – | 353 |
Материал. Голотип ♂, инвентарный номер препарата 103/05; паратипы: 4♂♂, 1♀. Препараты голотипа, трех самцов-паратипов и самки-паратипа хранятся в Гельминтологическом музее РАН, Институт проблем экологии и эволюции им. А.Н. Северцова РАН, Центр паразитологии (г. Москва, Россия).
Типовое местонахождение. Вьетнам, дельта р. Меконг, рукав Котьен (Cổ Chiên), центральная часть русла. Координаты станции: 09°52.547′ с.ш., 106°31.733′ в.д., глубина 12.5 м, грунт – песок с серо-коричневым глинистым илом, соленость воды у дна 28.8 г/л. Сбор 06.12.2019 г.
Дополнительное местонахождение. Один самец из паратипов обнаружен в рукаве Котьен в прибрежной части русла в точке с координатами: 09°52.622′ с.ш., 106°31.965′ в.д., глубина 2.5 м, грунт – плотный песок с серой глиной, соленость воды у дна 12.7 г/л. Сбор 06.12.2019 г.
Описание. Морфометрическая характеристика голотипа и паратипов приведена в табл. 3.
Самцы. Тело длинное и тонкое. Кутикула гладкая, 2.0 мкм толщиной в среднем отделе тела. Соматические щетинки короткие и редкие. Голова плохо обособлена от остального тела. Губы сравнительно низкие. Шесть внутренних губных сенсилл в форме папилл. Шесть внешних губных сенсилл и четыре головные сенсиллы в форме коротких и тонких щетинок.
Длина внешних губных щетинок 4.3–4.6 мкм. Головные щетинки немного короче. Фарингостома (ротовая полость) обширная, бочонковидная, с сильно кутикулизированными стенками. В ротовой полости три зуба. Левый субвентральный зуб намного крупнее правого субвентрального и дорсального зубов. Длина ротовой полости в 1.3–1.6 раза больше ширины области губ. Фовеи амфидов кармановидные, расположены на уровне середины или передней половины длины ротовой полости. Фаринкс мускулистый, слегка расширяется к своему основанию. Кардий удлиненно-конический, вдается в просвет средней кишки. Ренетта и ее канал не обнаружены. Экскреторная пора расположена на расстоянии 65–72 мкм от переднего конца тела.
Семенники парные, противопоставленные и расположены слева от средней кишки. Спикулы тонкие, слегка изогнуты, их длина равна или слегка превышает диаметр тела в области клоаки. Рулек отсутствует. Перед клоакой расположена крупная, шиповидная щетинка. Вокруг клоаки также расположены мелкие щетинки. Хвост сравнительно короткий, удлиненно-конический, вентрально загнут. Одна папилла расположена на хвосте, вентрально и довольно близко к клоаке. Каудальные щетинки короткие. Каудальные железы и спиннерета хорошо развиты.
Самка. По общей морфологии подобны самцам. Строение кутикулы и переднего конца тела как у самцов. Кутикула гладкая. Шесть внутренних губных сенсилл в форме папилл. Шесть внешних губных сенсилл и четыре головные сенсиллы в форме тонких щетинок. Длина внешних губных щетинок 4.4 мкм. Головные щетинки немного короче. Фовеи амфидов расположены на уровне передней половины длины ротовой полости. Ротовая полость бочонковидная, с сильно кутикулизированными стенками. Ее размеры 30 × 17 мкм. В ротовой полости три зуба, причем левый субвентральный зуб намного крупнее правого субвентрального и дорсального зубов. Фаринкс мускулистый, слегка расширяется к своему основанию. Кардий удлиненно-конический, вдается в просвет средней кишки. Длина ректума примерно равна диаметру тела в области ануса.
Яичник один, передний, загнут. Вульва в форме поперечной щели. Вагина короткая, с тонкими стенками, наклонена к переднему концу тела. Матка обширная, заполнена сперматозоидами. В матке три яйца размером 111–152 × 49–68 мкм. Строение системы de Mana разглядеть не удалось. Хвост сравнительно короткий, удлиненно-конический, слегка загнут вентрально. Каудальные железы и спиннерета хорошо развиты.
Дифференциальный диагноз. В состав рода Oncholaimus Dujardin, 1845 входят ~120 валидных видов (Smol, Mithumbi, Sharma, 2014; Bezerra et al., 2022). Род можно разделить на несколько видовых групп. Виды, самцы которых имеют на хвосте вентральную папиллу, формируют видовую группу “oxyuris”. В ее состав в настоящее время входят 20 видов (Gagarin, Gusakov, 2016; Tahseen et al., 2016; Tauheed, Ahmad, 2016; Qun, Yong, 2017). O. mekongis sp. n. по размерам тела и длине спикул морфологически ближе всего к видам O. balli Kreis, 1932 и O. menzeli Schneider, 1937. Отличается от обоих видов наличием крупной шиповидной щетинки перед клоакой; положением вентральной папиллы на хвосте самцов (расположена в передней половине хвоста, у O. balli и O. menzeli – в задней половине), относительно более толстым телом (а = 23–29 против а = 57–63 у самцов O. balli и а = 75–83 у самцов O. menzeli), более короткими внешними губными щетинками (их длина 4.3–4.7 мкм, 14–17% ширины области губ против 7.5 мкм, 35% ширины области губ у O. balli и 6.5 мкм, 30–34% ширины области губ у O. menzeli); относительно более длинным и стройным хвостом у самцов (с = 22–28, с ' = 2.8–3.3 против с = 33–41, с ' = 2.2 у O. balli и с = 34–46, с ' = = 1.8–2.2 у O. menzeli) (Kreis, 1934; Schneider, 1937).
Этимология. Видовое название означает “меконгский”, “из Меконга”.
Список литературы
Гагарин В.Г., Нгуен Ву Тхань. 2009. Три новых вида монхистерид (Nematoda, Monhysterida) из мангровых зарослей в дельте реки Меконг, Вьетнам // Зоол. журн. Т. 88. № 19. С. 1170.
Фан Ке Лонг, Нгуен Динь Ты, Гагарин В.Г. 2020. Daptonema paramonovi sp. n. (Nematoda, Monhysterida) из мангрового биотопа во Вьетнаме // Зоол. журн. Т. 99. № 6. С. 616.
Bezerra T.N., Eiseindle U., Hedda M. et al. 2022. Nemys World Database of Nematodes. Oncholaimus Dujardin, 1845. World Register of Marine Species. [Online]. Available at: http://www.marinespecies.org/aphia.php?p=taxdetails&id=2573 on 2022-03-09
Gagarin V.G., Gusakov V.A. 2016. Oncholaimus rivalis sp. n. (Nematoda, Enoplida) from highly mineralized rivers of Lake Elʹton basin, Russia // Int. J. Nematol. V. 22. № 1, 2. P. 13.
Gagarin V.G., Nhuyen Dinh Tu. 2021. Two new species of the order Monhysterida Filipjev, 1939 (Nematoda) from the Yan Uc River Mouth in Vietnam // Inland Water Biol. V. 14. № 5. P. 517. https://doi.org/10.1134/S1995082921050059
Gerlach S.A. 1978. Food-chain relation-ship in subtidal silly sand marine sediments and the role of feiofauna in stimulating bacterial productivity // Oecologia. V. 13. P. 55.
Gerlach S.A., Riemann F. 1973, 1974. The Bremerhaven Checlist of Nematoda Adenophorea excluding the Dorylaimida. Part 1, 2 // Veröffentli. Inst. Meeresforschung in Bremerhaven. Suppl. 4. P. 1.
Fonseca G., Bezerra T.M. 2014. Order Monhysterida Filipjev, 1929 // Handbook of Zoology. Gastrotricha, Cycloneuralia and Gnathifera. V. 2. Nematoda. Berlin/ Boston: Walter de Gruter. P. 435.
Kreis H. 1934. Oncholaiminae Filipjev, 1916. Eine monographische studie // Capita Zoolog. Bd 4(5). S. 1.
Lorenzen S. 1977. Revision der Xyalidae (freilebenden Nematoden) aus der Grunlage einer Kritischen Analyse von 56 Arten aus Nord-und Ostsee // Veröf. Inst. Meeresforsch. Bremerhaven. Bd 16. № 3. S. 197.
Qun G., Yong H. 2017. Oncholaimus zhangi sp. nov. (Oncholaimidae, Nematoda) from the intertidal zone of the East China Sea // Chinese Journal of Oceanology and Limnology. V. 35. № 5. P. 1212.
Schneider W. 1937. Freilebenden Nematoden der dentschen limnologischen Sundaexpdition nach Sumatra, Java and Bali // Archiv Hydrobiol. Suppl. 15. Tropische Binnengewasser. Bd 7. S. 30.
Smol N., Coomans A. 2005. Order Enoplida // Freshwater nematodes: Ecology and Taxonomy. Wallingford: CABI Publising. P. 225.
Smol N., Muthumbi A., Sharma J. 2014. Order Enoplida // Handbook of Zoology. Gastrotricha, Cycloneuralia and Gnathifera. V. 2. Nematoda. Berlin/ Boston: Water de Gruyter. P. 193.
Tahseen Q., Siddiqi M.R., Mustaqim M. 2016. Description of two new species of Oncholaiminae Filipjev, 1916 (Nematoda, Enoplida) // Int. J. Nematol. V. 26. № 1, 2. P. 41.
Tauheed U., Ahmad W. 2016. Description of a new species of marine nematodes genus Oncholaimus Dujardin, 1845 (Oncholaimidae: Oncholaiminae) from Bay of Bengal, India // Int. J. Nematol. V. 26. № 1, 2. P. 51.
Tchesunov A.V., Milutina M.A. 2005. Marine free-living nematode species of the superfamily Monhysteridea from Arctic Abiss // Zootaxa. V. 1951. P. 19.
Timm R.W. 1961. The marine nematodes of the Bay of Bengal // Proceedings of the Pakistan Academy of Sciences. V. 1. P. 25.
Wieser W., Kanwisher J. 1960. Ecological and physiological studies on marine nematodes from a small salt marsh near Woods Hole, Massachusetts // Limnol., Oceanogr. V. 6. P. 262.
Williams D.D., Williams N.E. 1974. A counterstaining technique for use in sorting benthic samples // Limnol., Oceanogr. V. 19. № 1. P. 152.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод