

Ботанический журнал, 2019, T. 104, № 2, стр. 171-190
СТРОЕНИЕ И РАЗВИТИЕ ПЫЛЬНИКА НЕКОТОРЫХ ВИДОВ ПОДРОДА ESULA РОДА EUPHORBIA (EUPHORBIACEAE)
Г. М. Анисимова *
Ботанический институт им. В.Л. Комарова РАН
197376 С.-Петербург, ул. проф. Попова, 2, Россия
* E-mail: galina0353@mail.ru
Поступила в редакцию 27.12.2018
После доработки 13.01.2019
Принята к публикации 15.01.2019
Аннотация
Проведено сравнительное исследование развития и строения пыльника у 4 видов, относящихся к разным секциям рода Euphorbia (подрод Esula, сем. Euphorbiaceae). Установлено значительное сходство в способе формирования стенки микроспорангия с наружной стороны (тип двудольных), строении сформированной стенки пыльника (обычно из 4 слоев: эпидермиса, эндотеция, среднего слоя и тапетума), наличии симультанного типа микроспорогенеза, преимущественно тетраэдральных тетрад микроспор, 2-клеточных зрелых пыльцевых зерен, секреторного клеточного тапетума, эндотеция с фиброзными утолщениями. Детальный анализ позволил выявить ряд особенностей в строении пыльника изученных видов сем. Euphorbiaceae. Еще до образования археспория в формирующихся микроспорангиях дифференцируется группа инициальных клеток, из которых формируются слои стенки микроспорангия не только с дистальной, но и с проксимальной и латеральных сторон. На наружной тангентальной стенке клеток эпидермиса откладывается эпикутикулярный воск в виде извилистых неглубоких складок. Реорганизация клеточного секреторного тапетума в амебоидный сопровождается исчезновением стенок между клетками и локализацией орбикул на внутренней тангентальной, а у E. seguieriana и на 1/3 поверхности смежных радиальных мембран тапетума. Время исчезновения клеточных стенок в тапетуме различается – на стадии тетрад микроспор у E. seguieriana, распада тетрад и образования микроспор у E. stepposa, одиночных микроспор у S. iberica и E. petrophila. Поскольку клетки тапетума разрушаются во время созревания 2-клеточных пыльцевых зерен, то длительное их функционирование, вероятно, ингибирует формирование утолщений стенок клеток эндотеция и образование фиброзных поясков, которые появляются очень поздно (почти перед вскрыванием пыльника), а не на стадии микроспор. Обнаруженные флюоресцирующие глобулы в клетках эндотеция E. iberica (секция Esula) и E. petrophila (секция Pithyusa), вероятно, являются сферокристаллами, выявленными ранее другими авторами в клетках тесты видов секции Pithyusa.
Сравнительно-эмбриологические исследования позволяют оценить значимость ряда признаков при решении спорных вопросов систематики и филогении. Среди таких таксонов, которые активно изучаются в настоящее время, является сем. Euphorbiaceae. Составлена общая эмбриологическая характеристика этого семейства, при этом в нее включены как женские, так и мужские эмбриональные структуры (Kapil, 1961; Batygina, Kolesova (Titova), 1983; Murgia et al., 1986; Kapil, Bhatnagar, 1994; Tokuoka, Tobe, 1995, 1999; Kamelina, 2009). На основе эмбриологического изучения (Franca, De-Paula, 2017) было подтверждено выделение рода Pera из сем. Euphorbiaceae в самостоятельное сем. Peraceae по молекулярно-генетическим данным (Wurdack, Davis, 2009).
Подобным образом в настоящее время анализируется род Euphorbia, особенно его подрод Esula Pers., выделенный наряду с подродами Athymalus Neck. ex Rchb. (= Rhizanthium (Boiss.) Wheeler), Euphorbia и Chamaesyce Raf. на основании молекулярно-филогенетического анализа. Недавно система подрода Esula была существенно пересмотрена с использованием ядерных (ITS) и хлоропластных (ndhF) маркеров (Riina et al., 2013; Geltman, 2016). На основании построения филогенетического древа выделены секции, объем и характеристики которых отличаются от принятых ранее. Именно поэтому необходимы сравнительно-эмбриологические исследования, позволяющие получить подробные данные по развитию различных эмбриональных структур. Результаты недавних исследований морфогенеза семязачатка и семени у 6 видов из секций Helioscopia и Esula подрода Esula Pers. рода Euphorbia L. позволили дополнить список признаков, полезных для целей систематики этого рода (Titova et al., 2015, 2018; Vinogradova, 2017). Такими признаками могут быть характеристики пыльника у видов подрода Esula, однако они изучены недостаточно.
Исходя из этого, цель настоящей работы состояла в сравнительном анализе развития пыльника у 4 видов из 2 секций подрода Esula Pers. рода Euphorbia L.: Euphorbia iberica Boiss. (секция Esula (Pers.) Dumort.), E. petrophila C. A. Mey., E. seguieriana Neck. и E. stepposa Zoz. (секция Pithyusa (Raf.) Lazar.).
МАТЕРИАЛ И МЕТОДИКА
Материалом для исследования послужили растения 4 видов рода Euphorbia (подрод Esula, сем. Euphorbiaceae): E. iberica, E. petrophila, E. seguieriana и E. stepposa. Сбор и фиксация материала проведены Г.Ю. Виноградовой в мае 2014 г.: E. seguieriana и E. stepposa – Пятигорск, подножие горы Бештау; E. iberica и E. petrophila – Пятигорск, гора Машук (окрестности ЭБС БИН РАН).
Для фиксации репродуктивных структур использовали смесь FAA (70° этиловый спирт, 40% формалин, ледяная уксусная кислота в пропорции 100:7:7). Дегидратация, инфильтрация парафином и окраска препаратов проводилась по общепринятой методике (Pausheva, 1974). Срезы толщиной 10–12 мкм сделаны на микротоме Microm HM 325 (Carl Zeiss, Germany), окрашивание фуксин-сернистой кислотой по Фёльгену (ядра клеток) с подкраской гематоксилином по Эрлиху (цитоплазма) и алциановым синим (клеточные стенки), а также сафранином с подкраской алциановым синим по Картису (Prozina, 1960). Исследование препаратов проводили с помощью микроскопа Axioplan 2 (Carl Zeiss, Germany), рисунки выполнены с помощью рисовального аппарата РА-7 на этом же микроскопе. Фотографии выполнены с помощью цифровой камеры-насадки (Axiocam MRc5) на микроскопе Axioplan (Carl Zeiss, Германия).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Соцветие. У изученных видов, как и у других молочаев, генеративные структуры формируются в особом цимозном соцветии – циатии. Каждый циатий снаружи прикрыт бокаловидной обверткой, образованной пятью сросшимися листочками с 4 экстрафлоральными нектарниками продолговато-овальной формы. Внутри соцветия располагаются тычиночные и единственный пестичный цветки. Тычиночные цветки, не имеющие околоцветника и состоящие только из одной тычинки на короткой цветоножке, собраны по 2–4 (чаще 3) в 5 кластеров по периферии циатия. Количество тычинок в циатиях исследованных видов обычно 15, однако у E. iberica их может быть 10, а у E. seguieriana 20. Они располагаются, как правило, в виде 3 кругов (верхний, средний и нижний) (табл. I, 2), но в некоторых циатиях E. iberica число кругов составляет только 2 (верхний и нижний) либо 4 (верхний, 2 средних и нижний) у E. seguieriana.
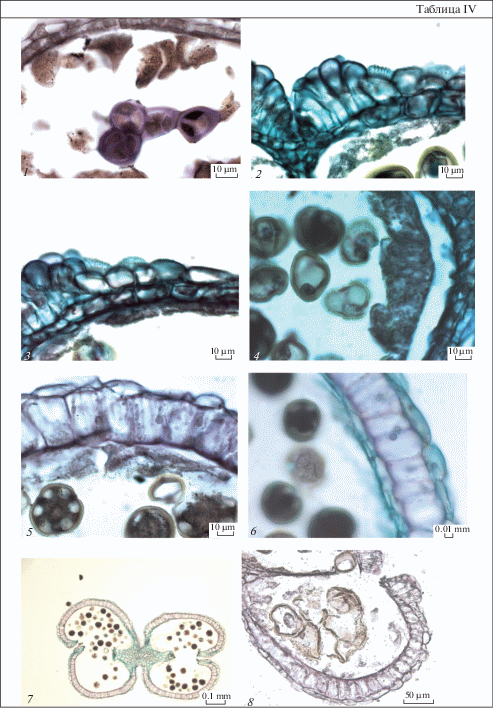
Таблица I. Строение пыльника у Euphorbia seguieriana.
1 – расположение тычиночных цветков в виде 3 кругов в циатии; 2 – формирование стенки микроспорангия до мейоза; 3 – микроспороциты; 4 – микроспоры после распада тетрад; 5 – микроспоры перед делением; 6 – пыльник на стадии микроспор; 7 – стенка микроспорангия на стадии 2-клеточных пыльцевых зерен. Масштабные линейки, мкм: 1, 6 – 50, 2–5, 7 – 10.
Table I. Anther structure in Euphorbia seguieriana.
1 – arrangement of staminate flowers as 3 circles in cyathium; 2 – wall microsporangium formation before meiosis; 3 – microsporocyte; 4 – microspores after tetrad decay; 5 – microspore before division; 6 – anther at microspore stage; 7 – wall microsporangium at stage of 2-celled pollen grains. Scale bars, µm: 1, 6 – 50, 2–5, 7 – 10.
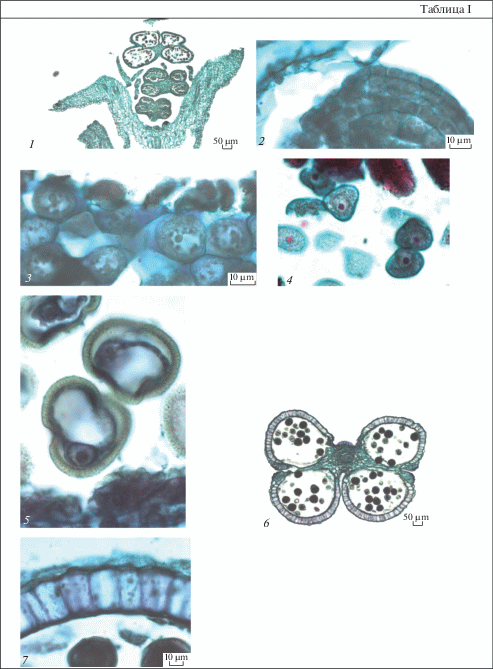
Первыми закладываются тычинки верхнего круга, затем среднего (или средних) и последними – нижнего. Тычинки одного и того же циатия различаются длиной тычиночных нитей, размерами пыльников и темпами развития. У тычинок наружного круга тычиночные нити самые длинные, а пыльники наиболее крупные. Микроспорогенез также начинается в пыльниках тычинок верхнего круга, затем среднего и нижнего, но к моменту созревания пыльцы этот процесс синхронизируется. В некоторых пыльниках наблюдается незначительная асинхронность в развитии микроспорангиев.
Пыльники 4-гнездные. Развитие тычинки начинается с заложения меристематического бугорка. По мере его развития происходит дифференциация на тычиночную нить и пыльник, форма которого из цилиндрической становится 4-лопастной.
Обычно при типизации стенки микроспорангия последовательность возникновения ее слоев характеризуют только с наружной стороны, начиная с момента дифференциации археспория (Davis, 1966), полагая, что слои ее внутренней и латеральных частей возникают из меристемы связника. Некоторые авторы считают, что с внутренней стороны они формируются из париетального слоя, располагающегося под спорогенными клетками (Babro, Voronova, 2018). Другие исследователи описывают сначала деление инициальной клетки в субэпидермальном слое микроспорангия, приводящее к образованию археспориальной и лежащей под ней париетальной клеток. Деление археспориальной клетки приводит к формированию над ней также париетальной клетки, а в дальнейшем париетального слоя, из которого и образуется стенка микроспорангия с наружной стороны (Torshilova, Batygina, 2005). Однако вопрос о происхождении слоев латеральных частей стенки микроспорангия остается нерешенным. Исследование, проведенное И.И. Шамровым (Shamrov, 2008), показало, что в субэпидермальном слое развивающегося пыльника дифференцируется группа инициальных клеток (центральная и латеральные), которые образуют как слои стенки микроспорангия, так и сам археспорий, а затем спорогенные клетки и микроспороциты. После периклинального деления центральной субэпидермальной клетки наружу отделяется археспориальная клетка, а внутрь – клетка, из которой формируется проксимальная (ближайшая к связнику) часть стенки микроспорангия. Латеральные субэпидермальные инициальные клетки за счет делений образуют латеральные части стенки микроспорангия. В результате периклинальных делений археспориальных клеток возникают наружный (париетальные клетки) и внутренний (спорогенные клетки) слои. Клетки париетального слоя делятся периклинальными и антиклинальными делениями, в результате чего образуются слои, входящие в состав дистальной части стенки микроспорангиев. Данные, полученные нами по изучению формирования стенки микроспорангия у Euphorbia, как и у ранее исследованных видов Kalanchoe (Anisimova, 2014, 2016), согласуются с представлениями Шамрова о том, что еще до выделения археспория в субэпидермальном слое каждого гнезда пыльника дифференцируется группа инициальных клеток, которые образуют как слои микроспорангия с проксимальной и латеральных сторон, так и археспориальные клетки. Из последних в дальнейшем возникают париетальные клетки, дающие слои стенки микроспорангия с дистальной стороны по типу двудольных (Davis, 1966), или центробежному типу (Teryokhin et al., 1993) (рис. 1, 1–4; 2, 1–4; табл. I, 2).
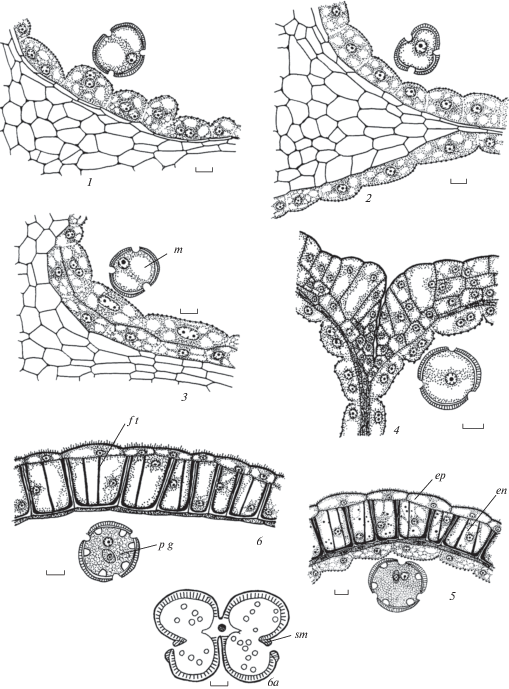
Рис. 1.
Строение и развитие пыльника у Euphorbia seguieriana.
1 – дифференциация инициалей стенки микроспорангия и образование археспория; 2–4 – формирование стенки микроспорангия и спорогенной ткани, клетки которой находятся перед микроспорогенезом, 5 – первые признаки реорганизации клеточного секреторного тапетума в амебоидный на стадии тетрад микроспор, произошло разрушение клеточных стенок; 6, 7 – продолжение реорганизации клеточного секреторного тапетума, на внутренней тангентальной и в основании радиальных мембран тапетума происходит образование орбикул; 6а – схема пыльника на стадии микроспор;
Fig. 1. Structure and development of the anther in Euphorbia seguieriana.
1 – differentiation of microsporangium wall initials and archesporium appearance; 2–4 – microsporangium wall formation and sporogenous tissue, cells of which are before microspogenesis, 5 – first sings of cellular secretory tapetum reorganization into ameboid tapetum at microspore tetrads, cell walls are destroyed; 6, 7 – continuation of cellular secretory tapetum reorganization, on inner tangential and in the base of radial membranes the formation of orbucules occurs; 6а – anther scheme at microspore stage;
Рис. 1. Строение и развитие пыльника у Euphorbia seguieriana (продолжение).
8 – стенка зрелого пыльника с фиброзными утолщениями в клетках эндотеция, тапетум и средний слой почти разрушены; aс – археспориальная клетка, cvb – проводящий пучок связника, en – эндотеций, ep – эпидермис, epw – эпикутикулярный воск, ft – фиброзные утолщения в связнике, m – микроспора, mc – микроспороциты, ml – cредний слой, mwi – инициали стенки микроспорангия, o – орбикулы, pg – пыльцевое зерно, pl – париетальный слой, sc – спорогенные клетки, sm – стомиум, st – спорогенная ткань, t – тапетум, tm – тетрада микроспор. Масштабные линейки, мкм: 1–8–10, 6a – 100.
Fig. 1. Structure and development of the anther in Euphorbia seguieriana (continued).
8 – mature anther wall with fibrous thickenings in endothecium cells, tapetum and middle layer are almost destroyed; aс – archesporium cell, cvb – connective vascular bundle, en – endothecium, ep – epidermis, epw – epicuticular wax, ft – fibrous thickenings in the connective, m – microspore, mc – microsporocyte, ml – middle layer, mwi – microsporangium wall initials, o – orbicules, pg – pollen grain, pl – parietal layer, sc – sporogenous cells, sm – stomium, st – sporogenous tissue, t – tapetum, tm – tetrad of microspores. Scale bars, µm: 1–8 –10, 6a – 100.
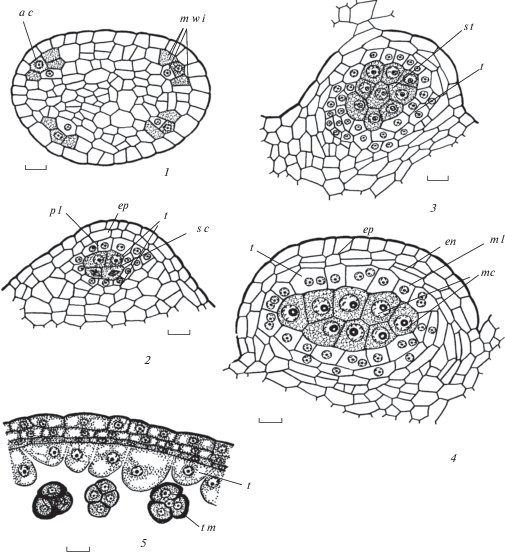
Сформированная стенка пыльника представлена эпидермисом, эндотецием, средним слоем и тапетумом (рис. 3, 1, 2; 4, 3, 4). Клетки эпидермиса крупные, почти квадратные у E. iberica (рис. 3, 1, 3) и вытянуты в тангентальном направлении у E. seguieriana (рис. 1, 4, 6) и E. stepposa (рис. 2, 4, 5). Эндотеций состоит из крупных удлиненных (на поперечных срезах квадратных) клеток. Средний слой представлен узкими, вытянутыми вдоль поверхности пыльника клетками (рис. 1, 4; 2, 4). Отдельные клетки эндотеция и среднего слоя могут делиться, и тогда они становятся нерегулярно двуслойными (рис. 1, 4; 2, 4). Секреторный клеточный тапетум на этой стадии представлен, как правило, одноядерными клетками.
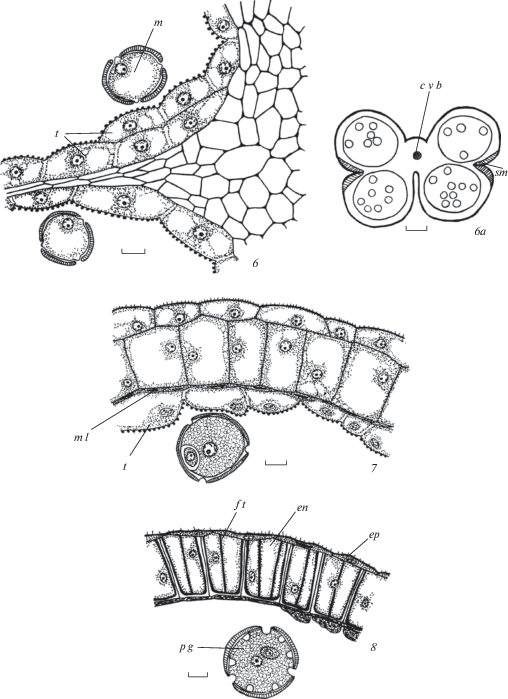
Рис. 2.
Строение и развитие пыльника у Euphorbia stepposa.
1 – дифференциация инициалей стенки микроспорангия и образование археспория; 2–4 – формирование стенки микроспорангия и спорогенной ткани, клетки которой находятся перед микроспорогенезом, 5 – первые признаки реорганизации клеточного секреторного тапетума в амебоидный на стадии тетрад микроспор и их распада на отдельные микроспоры, произошло разрушение клеточных стенок; 6 – продолжение реорганизации клеточного секреторного тапетума, на внутренней тангентальной мембране тапетума происходит образование орбикул;
Fig. 2. Structure and development of the anther in Euphorbia stepposa.
1 – differentiation of microsporangium wall initials and archesporium appearance; 2–4 – microsporangium wall formation and sporogenous tissue, cells of which are before microspogenesis, 5 – first sings of cellular secretory tapetum reorganization into ameboid tapetum at microspore tetrads and their decay into single microspores, cell walls are destroyed; 6 – continuation of cellular secretory tapetum reorganization, on inner tangential membrane of tapetum the formation of orbucules occurs;
Рис. 2. Строение и развитие пыльника у Euphorbia stepposa (продолжение).
7, 7а – схема пыльника и строение стомиума на стадии микроспор; 8, 9 – продолжение реорганизации клеточного секреторного тапетума, на внутренней тангентальной мембране тапетума происходит образование орбикул; 10 – стенка зрелого пыльника с фиброзными утолщениями в клетках эндотеция, тапетум и средний слой почти разрушены. Обозначения те же, что и на рис. 1. Масштабные линейки, мкм: 1–9 –10, 7a – 100, 10 – 50.
Fig. 2. Structure and development of the anther in Euphorbia stepposa (continued).
7, 7а – anther scheme and stomium structure at microspore stage; 8, 9 – continuation of cellular secretory tapetum reorganization, on inner tangential membrane of tapetum the formation of orbucules occurs; 10 – mature anther wall with fibrous thickenings in endothecium cells, tapetum and middle layer are almost destroyed. For abbreviations see Fig. 1. Scale bars, µm: 1–9 –10, 7a – 100, 10 – 50.
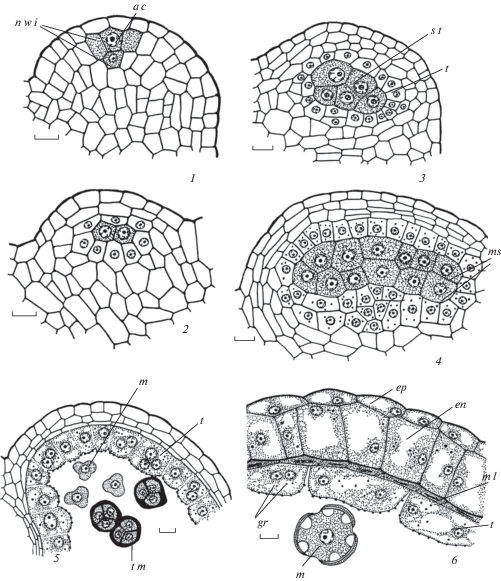
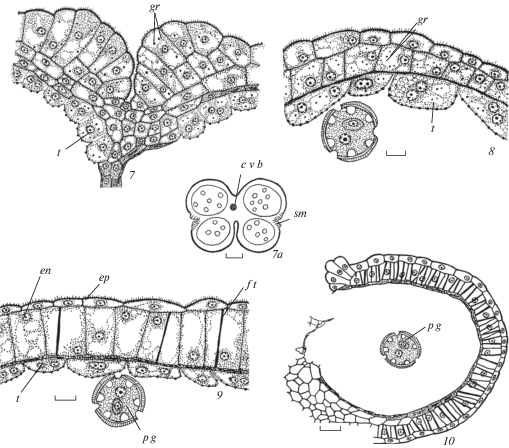
Рис. 3.
Строение пыльника у Euphorbia iberica.
1 – сформированная стенка микроспорангия на стадии тетрад микроспор; 2 – начало реорганизации клеточного секреторного тапетума в амебоидный тапетум, на внутренней тангентальной мембране тапетума происходит образование орбикул; 2а – схема пыльника на стадии микроспор; 3, 4 – стенка микроспорангия в период формирования 2-клеточных пыльцевых зерен, средний слой и тапетум разрушаются; 5 – стенка зрелого пыльника с фиброзными утолщениями в клетках эндотеция, тапетум и средний слой не выявляются. gr – гранулы. Остальные обозначения те же, что и на рис. 1 и 2. Масштабные линейки, мкм: 1–5 –10, 2а, 5a – 100.
Fig. 3. Anther structure in Euphorbia iberica.
1 – formed microsporangium wall at microspore stage; 2 – beginning of cellular secretory tapetum reorganization into ameboid tapetum, on inner tangential membrane the formation of orbucules occurs; 2а – anther scheme at microspore stage; 3, 4 – microsporangium wall during two-celled pollen grain formation, middle layer and tapetum are destroyed; 5 – mature anther wall with fibrous thickenings in endothecium cells, tapetum and middle layer are not revealed. gr – granules. For other abbreviations see Figures 1 and 2. Scale bars, µm: 1–5 –10, 2а, 5a – 100.
В ходе созревания пыльника клетки эпидермиса приобретают обычно линзовидную двояковыпуклую форму. На их наружной тангентальной стенке откладывается эпикутикулярный воск в виде извилистых неглубоких складок, которые на продольных срезах имеют вид волосков (рис. 1, 7, 8; 2, 8, 9; 3, 2–5; 4, 5, 6; табл. I, 7; II, 6, 7; III, 5; IV, 2, 3, 5, 6). В области стомиума клетки эпидермиса (на поперечном срезе 3–7 клеток каждого микроспорангия) крупнее остальных и вытянуты в радиальном направлении (рис. 1, 6а; 2, 7, 7а; 3, 2а, 5а; 4, 4; табл. I, 6; II, 1, 3; III, 1, 3; IV, 2). У E. petrophila в клетках стомиума в периплазматическом пространстве между плазмалеммой и клеточной оболочкой выделяется слой розового цвета (при окраске сафранином), пронизанный пектиновыми дендритами, через которые на поверхность кутикулы выделяется эпикутикулярный воск (табл. III, 6). Особые клетки в области стомиума морфологически похожи на папилловидные секреторные клетки рыльца и, возможно, участвуют в процессе привлечения насекомых во время опыления.
Вскрываются пыльники продольными щелями, лартрозно. При этом происходит разрыв и разрушение мелких клеток в области стомиума на границе смежных микроспорангиев одной теки (рис. 1, 6а; 2, 7, 7а, 10; 3, 2а, 5а; 4, 4, 6а; табл. I, 6; II, 1; IV, 7).
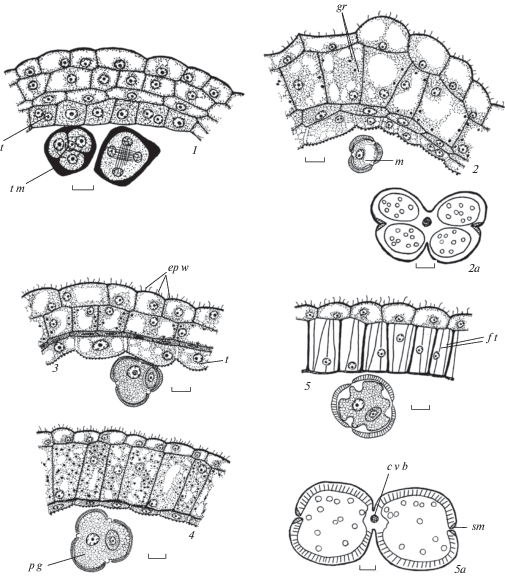
Таблица II. Строение пыльника у Euphorbia iberica.
1–4 – пыльник на стадии микроспор: 1 – общий вид пыльника, 2–4 – строение стенки микроспорангия и стомиума; 5 – стомиум во время вскрывания пыльника; 6, 7 – стенка микроспорангия почти зрелого пыльника, видны гранулы в клетках эндотеция; 8 – дегенерирующий пыльник. Масштабные линейки, мкм: 1, 5, 8 – 50, 2–4, 6, 7 – 10.
Table II. Anther structure in Euphorbia iberica.
1–4 – anther at microspore stage: 1 – anther general view, 2–4 – structure of wall microsporangium and stomium; 5 – stomium during anther dehiscence; 6, 7 – wall microsporangium of near mature anther, granules in endothecium cells are seen; 8 – degenerating anther. Scale bars, µm: 1, 5, 8 – 50, 2–4, 6, 7 – 10.
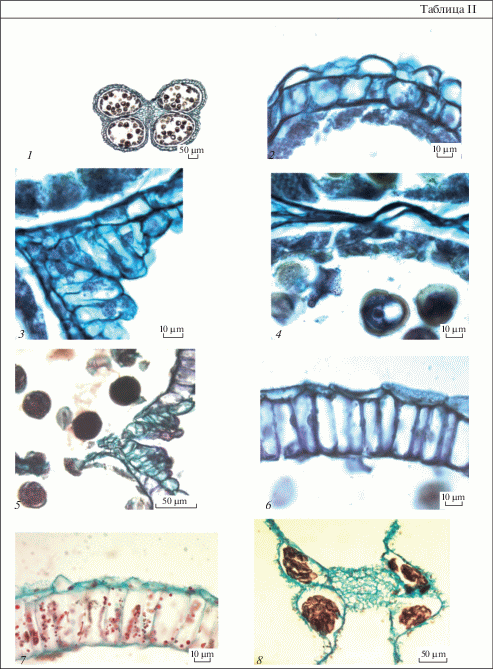
Таблица III. Строение пыльника у Euphorbia petrophila.
1–3 – пыльник на стадии микроспор: 1 – стомиум, 2, 3 – строение тапетума, клеточные стенки разрушены, видны отдельные протопласты; 4 – на внутренних тангентальных мембранах тапетума имеются орбикулы; 5 – стенка микроспорангия почти зрелого пыльника; 6 – стомиум в пыльнике перед вскрыванием, между плазмалеммой и клеточной оболочкой выделяется слой розового цвета. Масштабные линейки, мкм – 10.
Table III. Anther structure in Euphorbia petrophila.
1–3 – anther at microspore stage: 1 – stomium, 2, 3 – tapetum structure, cell walls are destroyed, separate protoplasts are seen; 4 – there are orbicules on inner tangential membranes of tapetum; 5 – wall microsporangium of near mature anther; 6 – stomium in the anther before dehiscence, the rose-color layer between plasmalemma and cell wall is recognized. Scale bars, µm – 10.
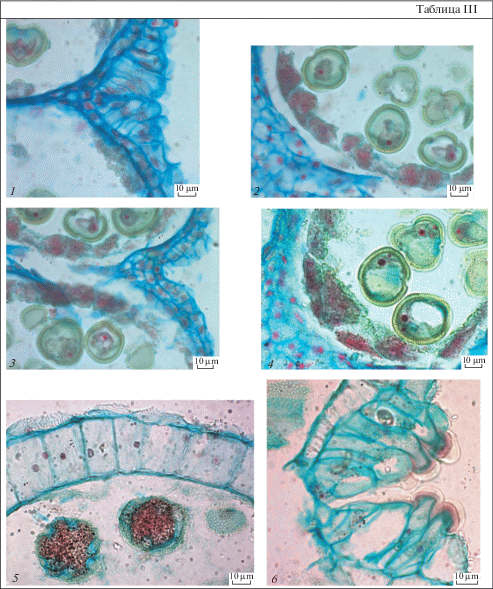
Таблица IV. Строение пыльника у Euphorbia stepposa.
1 – тетрады микроспор; 2, 3 – строение стомиума и стенки микроспорангия, на наружной тангентальной стенке клеток эпидермиса откладывается эпикутикулярный воск в виде извилистых неглубоких складок; 4 – строение клеток тапетума на стадии микроспор; 5, 6 – стенка микроспорангия почти зрелого пыльника, в клетках эндотеция глобулы; 7 – вскрывшийся пыльник; 8 – дегенерирующий пыльник. Масштабные линейки, мкм: 1–6, 8 – 10, 7 – 100.
Table IV. Anther structure in Euphorbia stepposa.
1 – tetrads of microspores; 2, 3 – structure of stomium and microsporangium wall, on the outer tangential walls of epidermal cells epicuticular wax is deposited in the form of sinuous, shallow folds; 4 – structure of tapetum cells at microspore stage; 5, 6 – wall microsporangium of near mature anther, globules in the endothecium cells are seen; 7 – dehiscent anther; 8 – degenerating anther. Scale bars, µm: 1–6, 8 – 10, 7 – 100.
Клетки эндотеция тоже увеличиваются в размерах, их внутренние тангентальные и радиальные стенки неравномерно утолщаются. Фиброзные пояски в них появляются очень поздно – в ходе созревания пыльцевых зерен (рис. 1, 8; 2, 9, 10; 3, 5; 4, 5, 6; табл. I, 7; II, 6, 7; III, 5; IV, 5, 6), при этом их толщина намного меньше по сравнению с толщиной стенок.
Клетки среднего слоя первыми подвергаются деструкции. Они постепенно сжимаются и в зрелом пыльнике представлены в виде узкого темно-окрашенного сплошного слоя (рис. 1, 7, 8; 2, 8, 9; 3, 2–5; 4, 5).
Еще до мейоза густоплазменные клетки тапетума увеличиваются в размерах, становятся таблитчатыми и в результате деления ядер становятся 1–2-ядерными у E. seguieriana (рис. 1, 4), а у E. stepposa на стадии тетрад в некоторых случаях 3-ядерными (рис. 2, 5), как в дистальной, так и других частях стенки микроспорангия (рис. 2, 7; 4, 1–4). Со стороны связника и перегородки между теками клетки тапетума часто крупнее и могут делиться, формируя обычно 2 слоя (рис. 1, 6; 2, 7; 4, 3). Наиболее крупные клетки тапетума наблюдали у E. stepposa.
У изученных видов клеточный секреторный тапетум в премейотическом и мейотическом периодах характеризуется клеточной структурой. Однако уже на стадии тетрад микроспор у E. seguieriana (рис. 1, 5), распада тетрад и образования микроспор у E. stepposa (рис. 2, 5), одиночных микроспор у S. iberica (рис. 3, 2) и E. petrophila (рис. 4, 1–4) стенки между клетками тапетума начинают разрушаться (табл. II, 2–4; III, 2–4; IV, 2–4). Следует отметить, что процесс исчезновения клеточных стенок сопровождается выпячиванием образующихся протопластов внутрь полости микроспорангия. Протопласты клеток тапетума остаются в пристенном положении, где разрушаются, причем их остатки с ядрами сохраняются почти до созревания пыльника. Тапетум в виде густоплазменного слоя сохраняется вплоть до объединения микроспорангиев в теках. Цитоплазма вакуолизируется, протопласты постепенно уменьшаются в размерах, а их содержимое лизирует. Таким образом, у изученных видов Euphorbia происходит реорганизация клеточного секреторного тапетума в амебоидный (протопласты тапетума выпячиваются в полость микроспорангия, внедряясь между микроспорами) (Kamelina, 1994).
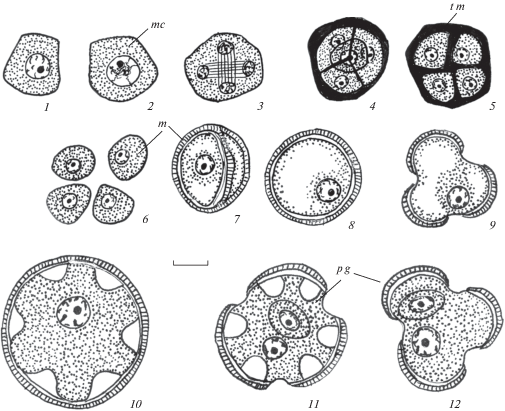
Рис. 4.
Строение пыльника у Euphorbia petrophila.
1–3 – строение клеток внутренней части тапетума на стадии микроспор, одно- и двуслойный тапетум, начало реорганизации клеточного секреторного тапетума в амебоидный тапетум, на внутренней тангентальной мембране тапетума происходит образование орбикул; 4–6 – строение стомиума и стенки микроспорангия в период формирования 2-клеточных пыльцевых зерен, средний слой и тапетум разрушаются; 6а – схема пыльника во время вскрывания. Обозначения те же, что и на рис. 1–3. Масштабные линейки, мкм: 1–5 – 10, 2а, 5a – 100.
Fig. 4. Anther structure in Euphorbia petrophila.
1–3 – structure of inner part of tapetum cells, one – and two-layered tapetum, beginning of cellular secretory tapetum reorganization into ameboid tapetum, on inner tangential membrane the formation of orbucules occurs; 4–6 – stomium structure and microsporangium wall during two-celled pollen grain formation, middle layer and tapetum are destroyed; 6а – anther scheme during dehiscence. For abbreviations see Figs. 1–3. Scale bars, µm: 1–6 – 10, 6a – 100.
Другой особенностью развития клеточного секреторного тапетума у видов Euphorbia является образование тапетальной мембраны вокруг каждого протопласта клеток тапетума после разрушения стенок между ними. Выявлены различия в локализации орбикул на тапетальной мембране. Они образуются обычно на внутренней тангентальной (вероятно, Pinaceae-тип), а у E. seguieriana и на 1/3 поверхности смежных радиальных мембран тапетума (Poaceae-тип) (Ogorodnikova, 1994).
Известно, что главной функцией тапетальной ткани является снабжение микроспороцитов и формирующихся микроспор питательными веществами, среди которых важную роль играют полисахариды, накапливающиеся внутри полости микроспорангия. Клетки тапетума также секретируют фермент каллазу для растворения каллозных оболочек тетрад микроспор, образуют спорополленин экзины и орбикул, контактируют с развивающимися пыльцевыми зернами. Кроме того, в тапетуме синтезируются вещества, ингибирующие процессы специализации клеток эндотеция в течение всего микроспорогенеза. После завершения синтеза спорополленина ингибирующее действие заканчивается, и тапетум начинает разрушаться, что обычно соответствует стадии вакуолизированных микроспор. Только после этого происходит формирование утолщений в клетках эндотеция (Chauhan, 1977). При длительном существовании тапетума, особенно у растений с мужской цитоплазматической стерильностью, наблюдается абортирование пыльцы, клетки эндотеция не переходят к формированию фиброзных поясков, и пыльники не вскрываются (Chauhan, 1979, Chauhan, Gupta 2006). Как уже было отмечено, у изученных нами видов Euphorbia тапетум сохраняет активность почти до зрелого состояния пыльника. Такое длительное существование тапетума, вероятно, ингибирует формирование утолщений стенок клеток эндотеция и образование фиброзных поясков. Обычно эти процессы можно наблюдать уже на стадии одиночных микроспор. У изученных видов утолщения клеточных стенок и фиброзные пояски в клетках эндотеция появляются очень поздно – в ходе созревания пыльцевых зерен, при этом их толщина намного меньше по сравнению с толщиной стенок.
Микроспорогенез происходит по симультанному типу (рис. 5, 1–3; табл. I, 3). Тетрады микроспор тетраэдральные, редко изобилатеральные, окружены толстой каллозой (рис. 1, 5; 2, 5; 3, 1; 5, 4, 5; табл. IV, 1). Постепенно каллоза растворяется и тетрады распадаются на микроспоры треугольной формы с закругленными углами (рис. 1, 6; 2, 5; 3, 2; 4, 1–3; 5, 6; табл. I, 4). Затем они вакуолизируются, округляются, увеличиваясь в размерах (рис. 5, 7, 8, 10; табл. I, 5; II, 4; III, 2–4; IV, 4). Перед делением ядро микроспоры оказывается в пристенном положении (рис. 5, 9). Зрелые пыльцевые зерна 2‑клеточные, 3-бороздные (рис. 1, 6–8; 2, 6, 8–10; 3, 2–5; 4, 4–6; 5, 11, 12).
Рис. 5.
Микроспорогенез и формирование пыльцевого зерна у Euphorbia stepposa (1, 8, 9, 11), E. seguieriana (2, 3, 5–7), E. iberica (4, 12) и E. petrophila (10).
1, 2 – микроспороцит; 3–5 – формирование тетрады микроспор; 6–10 – стадия микроспоры; 11, 12 – пыльцевое зерно. Обозначения те же, что и на рисунках 1–4. Масштабная линейка, мкм –10.
Fig. 5. Microsporogenesis and pollen grain formation in Euphorbia stepposa (1, 8, 9, 11), E. seguieriana (2, 3, 5–7), E. iberica (4, 12) and E. petrophila (10).
1, 2 – microsporocyte; 3–5 formation of microspore tetrad; 6–10 – microspore stage; 11, 12 – pollen grain. For abbreviations see Figures 1–4. Scale bar, µm –10.
В ходе развития пыльника в его тканях были выявлены включения, часто в виде флюоресцирующих глобул разного размера: у E. iberica и E. petrophila в клетках эндотеция (рис. 3, 2–4; 4, 5), у E. stepposa – тапетума, эндотеция и стомиума (рис. 2, 4–8). В зрелом пыльнике перед вскрыванием эти глобулы не обнаружены.
Для всех исследованных видов характерно наличие пыльников с деструктивными признаками в клетках слоев стенки микроспорангия. Также выявлены аномальные пыльцевые зерна, которые были гипертрофированы, имели меньшие размеры, деформированы и имели мало цитоплазмы (табл. II, 8; IV, 8).
Таким образом, проведено сравнительное исследование развития и строения пыльника у 4 видов рода Euphorbia (подрод Esula, сем. Euphorbiaceae), относящихся к разным секциям: Euphorbia iberica (секция Esula), E. petrophila, E. seguieriana и E. stepposa (секция Pithyusa). У них установлено значительное сходство в строении сформированной и зрелой стенок пыльника, микроспорогенезе, строении пыльцевого зерна, способе вскрывания пыльника. Оно проявляется в способе формирования стенки микроспорангия с наружной стороны (тип двудольных), строении сформированной стенки пыльника (обычно из 4 слоев: эпидермиса, эндотеция, среднего слоя и тапетума), наличии симультанного типа микроспорогенеза, преимущественно тетраэдральных тетрад микроспор, 2-клеточных пыльцевых зерен, секреторного клеточного тапетума, эндотеция с фиброзными утолщениями. Эти сведения обычно приводятся в любой традиционной работе по развитию и строению пыльника, и они совпадают с литературными данными по сем. Euphorbiaceae (Banerji, 1951; Kapil, 1961; Davis, 1966; Batygina, Kolesova (Titova), 1983; Murgia et al., 1986; Kapil, Bhatnagar, 1994; Kamelina, 2009).
Однако детальный анализ позволил нам выявить ряд особенностей, которые были обнаружены впервые для сем. Euphorbiaceae. Прежде всего, была описана последовательность возникновения слоев стенки микроспорангия не только с дистальной, а также с проксимальной и латеральных сторон из группы инициальных клеток, которые дифференцируются первыми еще до образования археспория.
Вторая особенность касается строения клеток эпидермиса. Было обнаружено, что в ходе созревания пыльника на их наружной тангентальной стенке откладывается эпикутикулярный воск в виде извилистых неглубоких складок, которые на продольных срезах имеют вид волосков. В области стомиума клетки эпидермиса крупнее остальных и вытянуты в радиальном направлении. При вскрывании пыльников продольными щелями происходит разрыв и разрушение мелких клеток на границе смежных микроспорангиев одной теки, но при этом крупные клетки сохраняются. У E. petrophila в клетках стомиума в периплазматическом пространстве между плазмалеммой и наружной клеточной стенкой выделяется слой розового цвета, пронизанный пектиновыми дендритами, через которые на поверхность кутикулы выделяется эпикутикулярный воск.
Клетки эндотеция также характеризуются спецификой развития. Утолщение внутренних тангентальных и радиальных стенок и появление фиброзных поясков отмечается очень поздно – в ходе созревания пыльцевых зерен, при этом их толщина намного меньше по сравнению с толщиной стенок.
У изученных видов клеточный секреторный тапетум в премейотическом и мейотическом периодах характеризуется клеточной структурой. Однако уже на стадии тетрад микроспор у E. seguieriana, распада тетрад и образования микроспор у E. stepposa, одиночных микроспор у E. iberica и E. petrophila стенки между клетками тапетума начинают разрушаться, что сопровождается выпячиванием образующихся протопластов внутрь полости микроспорангия. Это позволяет констатировать для изученных видов Euphorbia реорганизацию клеточного секреторного тапетума в амебоидный. Его особенностью является образование тапетальной мембраны вокруг каждого протопласта клеток тапетума после разрушения стенок между ними. Выявлены различия в локализации орбикул на тапетальной мембране. Они образуются обычно на внутренней тангентальной, а у E. seguieriana и на 1/3 поверхности смежных радиальных мембран тапетума.
Выявлена взаимосвязь между функционированием тапетума и строением клеток эндотеция. У изученных видов тапетум сохраняет активность почти до зрелого состояния пыльника. Такое его длительное существование, вероятно, ингибирует формирование утолщений стенок клеток эндотеция и образование фиброзных поясков. Обычно эти процессы можно наблюдать уже на стадии одиночных микроспор. Однако у видов Euphorbia утолщения клеточных стенок и фиброзные пояски в клетках эндотеция появляются очень поздно – в ходе созревания пыльцевых зерен после разрушения клеток тапетума.
Как мы уже отмечали, на основании развития семени у видов секции Helioscopia и Esula выявлены различия, проявляющиеся в степени массивности нуцеллуса, внутреннего интегумента и эндопахихалазы, степени развития системы постхалазального ветвления проводящего пучка рафе, характере развития халазальной части эндосперма, а также наличии сферокристаллов в клетках тесты. В последнем случае сферокристаллы выявлены у видов секции Esula (Titova et al., 2018). Возможно, обнаруженные нами флюоресцирующие глобулы разного размера в тканях пыльника E. iberica (секция Esula) также являются сферокристаллами. С другой стороны, они были выявлены в секции Pithyusa (E. stepposa и E. petrophila), причем не только в клетках эндотеция, но также тапетума и стомиума.
Список литературы
[Anisimova] Анисимова Г.М. 2014. Развитие и строение пыльника у Kalanchoe laxiflora и K. tubiflora (Crassulaceae) в условиях интродукции. – Труды международной научной конференции. Ботаника: история, теория, практика (к 300-летию основания Ботанического института им. В.Л. Комарова Российской академии наук). СПб. С. 8–12.
[Anisimova] Анисимова Г.М. 2016. Строение пыльника, микроспорогенез и пыльцевое зерно у Kalanchoe nyikae (Crassulaceae). – Бот. журн. 101 (12): 1378–1389.
[Babro, Voronova] Бабро А.А., Воронова О.Н. 2018. Развитие мужских репродуктивных структур у Helianthus ciliaris и H. tuberosus (Asteraceae). – Бот. журн. 103 (9): 1093–1108.
Banerji I. 1951. Pollen and embryo sac of two Euphorbiaceae. – Proc. Indian Acad. Sci. Sect. B. 34 (3): 172–181.
[Batygina, Kolesova (Titova)] Батыгина Т.Б., Колесова (Титова) Г.Е. 1983. Семейство Euphorbiaceae. – В кн.: Сравнительная эмбриология цветковых растений. Л. Т. 2. С. 267–277.
Chauhan S.V.S. 1977. Dual role of the tapetum. – Curr. Sci. 46 (19): 674–675.
Chauhan S.V.S. 1979. Development of endothecium in relation to tapetal behaviour in some male sterile plants. – Phytomorphology. 29 (3–4): 245–251.
Chauhan S.V.S., Gupta H.K. 2006. Suppression of endothecium development by malformed tapetum in the anthers of chemically treated Lens culinaris. – Phytomorphology. 56 (1–2): 10–16.
Davis G.L. 1966. Systematic embryology of angiosperms. New York etc. 528 p.
Franca R.O., De-Paula O.C. 2017. Embryology of Pera (Peraceae, Malpighiales): systematics and evolutionary implication. – J. Plant Res. 130 (4): 709–721.
[Geltman] Гельтман Д.В. 2016. Подрод Esula рода Euphorbia (Euphorbiaceae): система, филогения, географический анализ: Дис. … д-ра биол. наук. М. 348 с.
[Kamelina] Камелина О.П. 1994. Новый подход к классификации типов тапетума. – В кн.: Эмбриология цветковых растений. Терминология и концепции. СПб. С. 56–60.
[Kamelina] Камелина О.П. 2009. Систематическая эмбриология цветковых растений. Двудольные. Барнаул. 501 с.
Kapil R.N. 1961. Some embryological aspects of Euphorbia dulcis L. – Phytomorphology. 11: 24–36.
Kapil R.N., Bhatnagar A.K. 1994. The contribution of embryology to systematics of the Euphorbiaceae – Ann. Missouri Bot. Gard. 81 (2): 145–159.
Murgia M., Wilms H.J., Cresti M., Cesca G. 1986. Ultrastructure of pollen development in Euphorbia dulcis L. 1. Diploid plants. – Acta Bot. Neerl. 35 (4): 405–424.
[Ogorodnikova] Огородникова В.Ф. 1994. Тапетальная мембрана. – В кн.: Эмбриология цветковых растений. Терминология и концепции. СПб. С. 49–50.
[Pausheva] Паушева З.П. 1974. Практикум по цитологии растений. М. 288 с.
[Prozina] Прозина М.Н. 1960. Ботаническая микротехника. М. 206 с.
Riina R., Peirson J.A., Geltman D.V., Molero J., Frajman B., Pahlevani A., Barres L., Morawetz J.J., Salmaki Y., Zarre S., Kryukov A., Bruyns P.V., Berry P.E. 2013. A worldwide molecular phylogeny and classification of the leafy spurges, Euphorbia subgenus Esula (Euphorbiaceae). – Taxon. 62 (2): 316–342.
[Shamrov] Шамров И.И. 2008. Формирование спорангиев высших растений. – Бот. журн. 93 (12): 1817–1845.
[Teryokhin et al.] Терёхин Э.С., Батыгина Т.Б., Шамров И.И. 1993. Классификация типов стенки микроспорангия у покрытосеменных. Терминология и концепции – Бот. журн. 78 (6): 16–24.
[Titova et al.] Титова Г.Е., Яковлева О.В., Жинкина Н.А. 2015. Развитие семени у Euphorbia palustris (Euphorbiaceae). – Бот. журн. 100 (3): 226–248.
[Titova et al.] Титова Г.Е., Яковлева О.В., Жинкина Н.А., Гельтман Д.В. 2018. Развитие семени у некоторых видов секций Helioscopia и Esula подрода Esula рода Euphorbia (Euphorbiaceae) – Бот. журн. 103 (11): 1355–1389. https://doi.org/10.7868/S0006813618110017
Tokuoka T., Tobe H. 1995. Embryology and systematics of Euphorbiaceae sensu lato; a review and perspective. – J. Plant Res. 108 (1): 97–106. https://doi.org/10.1007/BF02344312
Tokuoka T., Tobe H. 1999. Embryology of tribe Drypeteae, an enigmatic taxon of Euphorbiaceae. – Plant Syst. Evol. 215(1/4): 189–208.
[Torshilova, Batygina] Торшилова А.А., Батыгина Т.Б. 2005. Развитие стенки пыльника тычиночного цветка Dioscorea nipponica (Dioscoreaceae) – Бот. журн. 90 (8): 1208–1215.
[Vinogradova] Виноградова Г.Ю. 2017. Морфогенез женских репродуктивных структур у видов Euphorbia (Euphorbiaceae), различающихся по типу развития зародышевого мешка. – Бот. журн. 102 (8): 1060–1093.
Wurdack K.J., Davis C.C. 2009. Malpighiales phylogenetics: gaining ground on one of the recalcitrant clades in the angiosperm tree of life – Amer. J. Bot. 96: 1551– 1570. https://doi.org/10.3732/ajb.0800207
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал