

Ботанический журнал, 2019, T. 104, № 3, стр. 442-454
ФОРМИРОВАНИЕ ГИНЕЦЕЯ У ACER GINNALA (ACERACEAE)
1 Российский государственный педагогический университет им. А.И. Герцена
191186 С.-Петербург, наб. р. Мойки, 48, Россия
2 Ботанический институт им. В.Л. Комарова РАН
197376 С.-Петербург,
ул. Проф. Попова, 2, Россия
* E-mail: ivan.shamrov@gmail.com
Поступила в редакцию 15.01.2019
После доработки 08.02.2019
Принята к публикации 12.02.2019
Аннотация
Бикарпеллятный гинецей Acer ginnala (Aceraceae) изменяется в ходе развития и на самых ранних стадиях лишь в нижней части завязи демонстрирует черты 2-гнездной синкарпии: наличие синасцидиатной и закрытой симпликатной зон. В средней части завязи наблюдается расхождение смежных краев плодолистиков и в центре симпликатной зоны формируется щель. Верхняя часть завязи представлена асимпликатной зоной разного строения. Она образована пликатными областями, в которых вентральные края каждого плодолистика смыкаются между собой (верхняя часть завязи) либо остаются раскрытыми (большая часть стилодиев). В дальнейшем основное значение приобретает постгенитальное срастание, и гинецей становится синкарпным на всем протяжении завязи. В средней и верхней частях завязи формируется закрытая симпликатная зона: в средней части она характеризуется совершенным срастанием, тогда как в верхней части вентральные края в каждом плодолистике лишь сомкнуты, в центре сохраняется щель, а между объединяющимися плодолистиками еще видны признаки несовершенного срастания.
Плацентация – сутуральная. В каждом гнезде симпликатной зоны возникают два семязачатка, развивающиеся сначала как ортотропные и располагающиеся со смещением. Еще до оплодотворения один из них начинает дегенерировать, при этом сохраняющийся семязачаток продолжает развитие на более массивной плаценте. Фертильный семязачаток, закладывающийся латерально на плаценте, постепенно оказывается в центре. Вследствие удлинения фуникулуса он начинает изгибаться в области микропиле, проявляя признаки асимметрии со стороны изгиба, в результате чего семязачаток становится геми-ортотропным. Проводящая система гинецея центрально-осевая и образована вентральными и дорсальными пучками, которые связаны между собой по всей стенке завязи.
На ранней стадии в завязи с дорсальной стороны каждого плодолистика начинается образование крыла. На границе объединения двух плодолистиков образуются две выемки на месте будущего разделения дробного плода. После оплодотворения формируется плод двукрылатка, разделяющийся на два односемянных мерикарпия.
Проблемы строения гинецея и принципы его типизации до сих пор не получили однозначного толкования (Shamrov, 2015). Исторически сложились два направления исследований, объясняющих его организацию: в соответствии с теорией кондупликатного плодолистика типы различаются, главным образом, строением в средней части завязи, тогда как согласно теории пельтатного плодолистика гинецей характеризуется зональностью и представляет в продольном направлении серию разных типов и промежуточных форм. Оба подхода, несмотря на преимущества и более объективное рассмотрение гинецея с позиций теории пельтатного плодолистика, механистически объясняют как разнообразие типов, так и возможные направления их эволюции. При характеристике типов очень часто обращается внимание лишь на возможность плодолистика или плодолистиков обеспечивать закрытость завязи с формированием гнезд, в которых располагаются семязачатки. Плацентации придается второстепенное значение либо плаценты описываются как структуры, на которых возникают семязачатки, и им даже отводится роль в образовании перегородок в завязи. Крайне важно, что формы плацент коррелируют с типами гинецея, при этом они характеризуются спецификой в строении, функционировании и занимают определенное положение в завязи (Shamrov, 2013). Особенности строения плацент, возможно, более значимый признак по сравнению с числом гнезд в зависимости от степени объединения плодолистиков. Еще одной особенностью изучения гинецея является то обстоятельство, что его характеристика часто дается только на одной из средних стадий формирования цветка (обычно в период развития зародышевого мешка). Поэтому актуальным является исследование морфогенеза гинецея в динамике.
Большой интерес представляет изучение гинецея у растений, строение которого различается в разных зонах и изменяется в ходе развития. К таким растениям относятся виды сем. Aceraceae. Обычно указывается, что гинецей у них синкарпный и состоит из двух сросшихся плодолистиков (Prozhina, 1953; Yudin, 1957; Takhtajan, 1966, 2009; Pshennikova, 2000). По отдельным стадиям его развития приводятся фрагментарные сведения (Hall, 1951; Khushalani, 1963; Peck, Lersten, 1991a). Еще одной особенностью является образование крыла в стенке завязи и формирование дробного плода двукрылатки. Морфогенез гинецея в динамике с учетом особенностей его строения в разных зонах не исследован.
В связи с этим нами было проведено исследование гинецея у одного из представителей сем. Aceraceae. В качестве модельного объекта был выбран вид Acer ginnala Maxim., ранее почти неизученный в этом отношении.
МАТЕРИАЛ И МЕТОДЫ
Материалом для исследования послужили растения Acer ginnala, произрастающие на систематическом участке Агробиостанции Российского государственного педагогического университета им. А.И. Герцена в поселке Вырица Гатчинского р-на Ленинградской обл. (табл. I, 1).
Таблица I. Ветка цветущего растения, цветки и плоды у Acer ginnala.
1 – ветка цветущего растения; 2, 3 – андромоноэция: обоеполые (бутоны) и тычиночные (вскрытые пыльники) цветки на одном растении; 4, 5 – ветки растения с плодами; 6 – дробные плоды-двукрылатки, различающиеся по размерам.
Plate I . Flowering branch, flowers and fruits in Acer ginnala.
1 – flowering branch; 2, 3 – andromonoecy: bisexual (flower buds) and staminate (opening anthers) flowers in the same plant; 4, 5 – branches with fruits, 6 – separate two-winged fruits differing in sizes.

Были изучены бутоны и цветки на разных стадиях развития. Материал фиксировали в смеси FАА (70% этиловый спирт, ледяная уксусная кислота и формалин в пропорции 100:7:7) и обрабатывали по общепринятой методике (Pausheva, 1974). Срезы толщиной 10–20 мкм окрашивали гематоксилином по Гейденгайну. Рисунки выполняли с применением рисовального аппарата на микроскопе Leica DM 1000, а микрофотографии – с помощью этого же микроскопа с использованием цифровой фотокамеры Leica EC3. Для получения изображений растений использован метод сканерографии (Gerasimov, 2004).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ОБСУЖДЕНИЕ
Цветки Acer ginnala желтовато-белого цвета. На одном растении встречаются как обоеполые, так и тычиночные цветки (табл. I, 2, 3), т.е. этот вид является андромоноэцичным (Demyanova, 2010). Как было показано ранее (Hall, 1951), эта половая форма характерна для многих видов рода Acer (A. platanoides L., A. ginnala, A. pensylvanicum L., A. pseudoplatanus L., A. saccharum Marsh., A. spicatum Lam.). Некоторым видам присуща андродиэция, при этом на одних особях развиваются обоеполые цветки, а на других – тычиночные (A. rubrum L., A. saccharinum) либо диэция, когда тычиночные и пестичные цветки формируются на разных растениях (A. negundo L.).
Мужские цветки зацветают первыми, тогда как в обоеполых цветках A. ginnala ни андроцей, ни гинецей не готовы к опылению, а сами цветки оказываются еще закрытыми (табл. I, 2, 3). Подобная разновременность в функционировании разных типов цветков в пределах соцветия или растения, препятствующая самоопылению, характерна для многих видов клена (Aksyenova, 1975). В обоеполых цветках, являющихся актиноморфными, 5-членными и пентациклическими, околоцветник состоит из 5 элементов чашечки и 5 элементов венчика. Андроцей представлен 8 тычинками, расположенными в 2 круга (рис. 1, 1, 7; 3, 5, 6), при этом 2 наружные тычинки короче остальных. Гинецей образован 2 плодолистиками, завязь почти верхняя. В тычиночных цветках из репродуктивных структур имеются только 8 тычинок в виде 2 кругов.
Рис. 1.
Строение гинецея в период мегаспорогенеза у Acer ginnala.
1–3 – нижняя часть завязи, синасцидиатная (1) и закрытая симпликатная (2, 3) зоны; 4, 5 – средняя часть завязи, закрытая (4) и открытая (5) симпликатные зоны; 6, 7 – верхняя часть гинецея, асимпликатная зона разного строения: вентральные края каждого плодолистика смыкаются между собой (верхняя часть завязи) (6) либо остаются раскрытыми (большая часть стилодиев) (7). dvb – дорсальный проводящий пучок, ip – несовершенное срастание, is – внутренняя тычинка, os – наружная тычинка, ov – семязачаток, p – плацента, pvb – плацентарный проводящий пучок, vvb – вентральный проводящий пучок. Масштабные линейки, мкм: 1–5 –200; 6, 7 – 500.
Fig. 1. Gynoecium structure during megasporogenesis in Acer ginnala.
1–3 – lower part of the ovary, synascidiate (1) and closed symplicate (2, 3) zones; 4, 5 – middle part of the ovary, closed (4) and open (5) symplicate zones; 6, 7 – upper part of the gynoecium, asymplicate zone of different structure: ventral borders of each carpelle are closed up (upper part of ovary) (6) or remain open (most part of stylodia) (7). dvb – dorsal vascular bundle, ip – imperfect fusion, is – inner stamen, os – outer stamen, ov – ovule, p – placenta, pvb – placentary vascular bundle, vvb – ventral vascular bundle. Scale bars, µm: 1–5 –200; 6, 7 – 500.
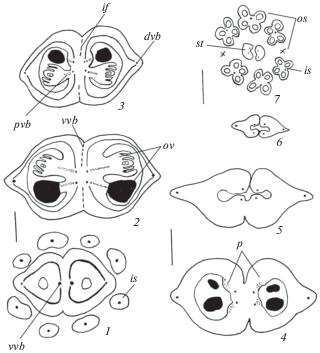
В развитии гинецея обоеполых цветков были выделены следующие стадии: 1) гинецей в период дифференциации структур семязачатка (мегаспорогенез), 2) гинецей в период формирования структур семязачатка (развитие зародышевого мешка), 3) гинецей перед опылением (сформированные семязачатки со зрелым зародышевым мешком).
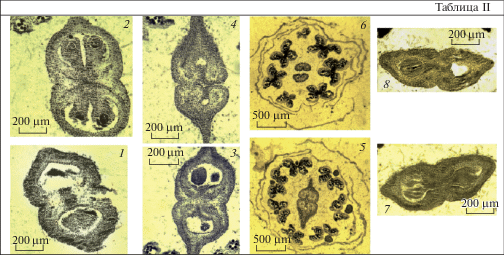
Гинецей во время дифференциации структур семязачатка (мегаспорогенез). Он образован 2 плодолистиками и ему присуще зональное строение. Завязь в нижней части 2-гнездная. В ней формируются конгенитально синасцидиатная (с базальной септой) и постгенитально симпликатная закрытая (с медианной септой) зоны. В симпликатной зоне еще видны границы между объединяющимися плодолистиками, как результат срастания несовершенного типа, при этом они полностью срастаются в центре. По строению она напоминает лежащую под ней синасцидиатную зону, лишенную семязачатков (рис. 1, 1–3; табл. II, 1, 2). В средней части завязи наблюдается расхождение смежных краев плодолистиков и в центре симпликатной зоны формируется щель (рис. 1, 4, 5; табл. II, 3, 4). Верхняя часть гинецея представлена асимпликатной зоной разного строения. Как в апокарпном гинецее, она образована пликатными областями, в которых вентральные края каждого плодолистика смыкаются между собой (верхняя часть завязи) (рис. 1, 6; табл. II, 5) либо остаются раскрытыми (большая часть стилодиев) (рис. 1, 7; табл. II, 6).
Таблица II. Строение гинецея в период мегаспорогенеза (1–6) и развития зародышевого мешка (7, 8) у Acer ginnala (поперечные срезы).
1, 2 – нижняя часть завязи, синасцидиатная (1) и закрытая симпликатная (2) зоны, в симпликатной зоне формируются 2 семязачатка в каждом гнезде; 3, 4 – средняя часть завязи, закрытая (3) и открытая (4) симпликатные зоны. 5, 6 – верхняя часть гинецея, асимпликатная зона разного строения: вентральные края каждого плодолистика смыкаются между собой (верхняя часть завязи) (5) либо остаются раскрытыми (большая часть стилодиев) (6); 7, 8 – симпликатная зона, в ней формируется только один семязачаток в каждом гнезде. Масштабные линейки, мкм: 1–4, 7, 8 – 200, 5, 6 – 500.
Plate II . Gynoecium structure during megasporogenesis (1–6) and embryo sac development (7, 8) in Acer ginnala (cross sections). 5, 6 – upper part of the ovary, asymplicate zone of different structure: ventral borders of each carpelle are closed up (upper part of ovary) (5) or remain open (most of stylodia) (6); 7, 8 – symplicate zone, only one ovule is formed in each locule. Scale bars, µm: 1–4, 7, 8 – 200, 5, 6 – 500.
1, 2 – lower part of the ovary, synascidiate (1) and closed symplicate (2) zones, 2 ovules are formed in each locule of symplicate zone; 3, 4 – middle part of the ovary, closed (3) and open (4) symplicate zones.
Плацентация – сутуральная, представленная центральной синплацентой в синасцидиатной, центрально-угловой в симпликатной и простой угловой плацентами в асимпликатной зонах, кроме стилодиев. В каждом гнезде на границе синасцидиатной и симпликатной зон возникают 2 семязачатка, ориентированные относительно плаценты под углом в 90° и развивающиеся сначала как ортотропные. Семязачатки каждого плодолистика располагаются со смещением относительно друг друга (рис. 1, 2–4). Проводящая система гинецея центрально-осевая и образована вентральными и дорсальными пучками, которые связаны между собой по всей стенке нижней части завязи (рис. 1, 1–3). Эта связь теряется на границе закрытой и открытой областей симпликатной зоны. Дорсальные пучки выявляются по всей длине завязи. Вентральные пучки каждого плодолистика в синасцидиатной зоне попарно объединены в гомокарпеллятные пучки. Они становятся самостоятельными в закрытой симпликатной зоне и от них отходят плацентарные пучки в семязачатки. На этой стадии развития вентральные пучки выявляются не только в завязи, но и в стилодиях (рис. 1, 1–7). Слияние вентральных пучков попарно в 2 синтетических гомокарпеллятных пучка было обнаружено у видов группы A, в которую, кроме A. ginnala, включены A. preudoplatanus, A. platanoides, A. spicatum (Hall, 1951).
Уже на этой стадии с дорсальной стороны завязи каждого плодолистика начинают выявляться признаки образования крыла, при этом за счет локальных делений клеток стенки завязи дорсальный проводящий пучок постепенно смещается на периферию крыла. В завязи на границе объединения двух плодолистиков образуются 2 выемки на месте будущего разделения дробного плода (рис. 1, 1–6; табл. II, 1–5).
Таким образом, в завязи на этой стадии развития только на границе синасцидиатной и закрытой симпликатной зон формируются семязачатки. Все остальные зоны являются стерильными и создают для развивающихся семязачатков и затем семян дополнительные пространства, способствующие их оптимальному размещению как до опыления, так и в период развития плода.
Гинецей в период формирования семязачатка (развитие зародышевого мешка). В ходе дальнейшего морфогенеза продолжаются процессы постгенитального объединения плодолистиков. В нижней и средней частях завязи формируются закрытые симпликатные зоны с медианной септой и 2-гнездным строением (границы между объединяющимися плодолистиками не выявляются, что свидетельствует о процессах совершенного срастания) (рис. 2, 1–3; табл. II, 7, 8). В верхней части завязи также начинается формирование закрытой симпликатной зоны, однако вентральные края в каждом плодолистике лишь сомкнуты, в центре сохраняется щель, а между объединяющимися плодолистиками еще видны признаки несовершенного постгенитального срастания (рис. 2, 4).
Рис. 2.
Строение гинецея в период развития зародышевого мешка у Acer ginnala.
1–4 – симпликатная зона в нижней (1), средней (2, 3) и верхней (4) частях завязи, смещение развивающихся семязачатков относительно друг друга в разных гнездах снизу вверх. Обозначения те же, что и на рис. 1. Масштабная линейка, мкм: 200.
Fig. 2. Gynoecium structure during embryo sac development in Acer ginnala.
1–4 – symplicate zone in lower (1), middle (2, 3) and upper (4) ovary parts, displacement of developing ovules relative to each other in different locules from bottom to top. For abbreviations see Fig. 1. Scale bar, µm: 200.
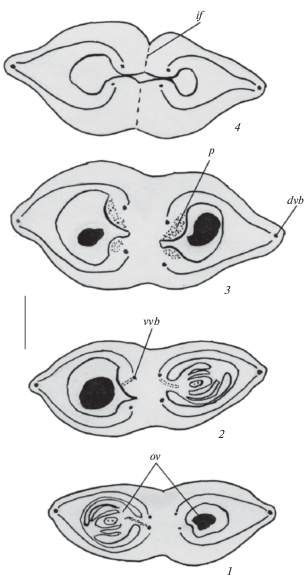
На этой стадии развития отчетливо выявляются признаки разрастания завязи, особенно в средней части. Увеличивающиеся в размерах семязачатки смещаются относительно друг друга в разных гнездах снизу вверх, занимая не только нижнюю, но и среднюю части завязи. В обоих гнездах из двух семязачатков один начинает дегенерировать. Этому предшествует разрастание одной из плацент каждого плодолистика, при этом сохраняющиеся семязачатки, закладывающиеся латерально на плаценте, постепенно оказываются в центре и начинают изгибаться в области микропиле, проявляя признаки асимметрии со стороны изгиба (рис. 2, 1–3; табл. II, 7, 8). Следует отметить, что эпидермальные клетки плаценты, особенно той, на которой развивается фертильный семязачаток, начинают функционировать как плацентарный обтуратор. Синасцидиатная зона в нижней части и формирующаяся симпликатная зона в верхней части завязи остаются стерильными, хотя развивающиеся семязачатки начинают их частично заполнять. Наблюдается усложнение хода проводящих пучков в завязи. Связь между вентральными и дорсальными пучками выявляется уже по всей длине завязи, при этом вентральные пучки на большем протяжении остаются самостоятельными. Продолжается увеличение размеров одностороннего крыла, особенно в длину (рис. 2, 1–4).
Гинецей перед опылением. Он становится синкарпным (преимущественно постгенитально) и 2-гнездным на всем протяжении завязи, при этом в нижней и верхней частях сохраняются стерильные зоны (рис. 3, 1, 2; табл. III, 1, 4). В каждом гнезде формируется по одному крассинуцеллятному и битегмальному семязачатку, которые располагаются преимущественно в средней части завязи. Семязачаток изменяет свое положение относительно плаценты. Этому способствует увеличение длины фуникулуса, что приводит к повороту семязачатка в геми-положение, и он из ортотропного становится геми-ортотропным (согласно представлениям Shamrov, 2017). Таким образом, сохраняющийся семязачаток занимает пространство, предназначенное для двух развивающихся семязачатков, и располагается по центру в каждом гнезде завязи. Однако исходно он возникает сбоку на плаценте, которая в ходе развития разрастается (также удлиняется фуникулус), что приводит к смещению семязачатка, и он становится геми-ортотропным (рис. 3, 3; табл. III, 2).
Рис. 3.
Строение гинецея перед опылением у Acer ginnala.
1–3 – симпликатная зона в нижней (1, 2) и средней (3) частях завязи; 4 – асимпликатная зона при переходе от верхней части завязи к стилодиям; 5–8 – строение стилодиев в свободной части, 2 наружные тычинки короче остальных (5, 6) и срастание стилодиев в дистальной части в головчатое рыльце, покрытое папилловидными клетками (7, 8). sg – рыльце, st – стилодий. Остальные обозначения те же, что и на рис. 1 и 2. Масштабные линейки, мкм: 1–3 –200, 4–7 – 500, 8 – 100.
Fig. 3. Gynoecium structure before pollination in Acer ginnala.
1–3 – symplicate zone in lower (1, 2) and middle (3) ovary parts; 4 – asymplicate zone in going from upper ovary part to stylodia; 5–8 – structure of stylodia in free part, 2 outer stamens are shorter than the others (5, 6), and fusing of stylodia into head papillate stigma in distal part (7, 8). sg – stigma, st – stylodium. For other abbreviations see Figs. 1 and 2. Scale bars, µm: 1–3 – 200, 4–7 – 500, 8 – 100.
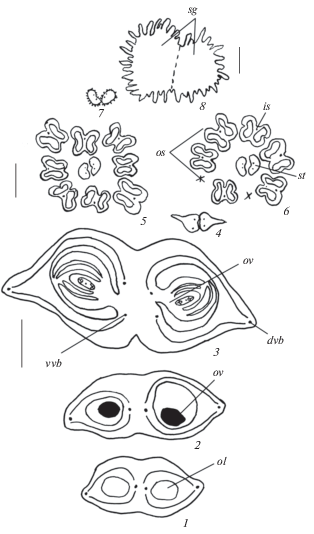
Таблица III. Строение гинецея перед опылением у Acer ginnala (поперечные срезы).
1–4 – симпликатная зона в нижней (1), средней (2, 3) и верхней (4) частях завязи; 5, 6 – строение рыльца. Масштабные линейки, мкм: 1–4 – 200, 5, 6 – 100.
Plate III . Gynoecium structure before pollination in Acer ginnala (cross sections).
1–4 – symplicate zone in lower (1), middle (2, 3) and upper (4) ovary parts; 5, 6 – structure of stigma. Scale bars, µm: 1–4 – 200, 5, 6 – 100.
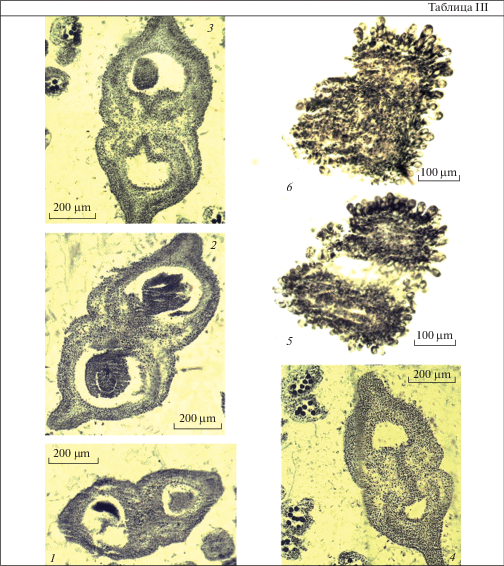
К началу опыления лопасти рыльца обоих стилодиев объединяются в дистальной части в одну головчатую структуру. Поверхность рыльца сухая и покрыта папилловидными клетками (рис. 2, 5–8; табл. III, 5, 6), что было отмечено ранее для A. ginnala и других 5 видов Acer (Peck, Lersten, 1991b).
После оплодотворения на базе гинецея, главным образом 2-гнездной завязи, формируется дробный плод двукрылатка, каждая часть которого (мерикарпий) является односемянной (табл. I, 4, 5). Размеры крыла мерикарпиев могут сильно различаться (табл. I, 6). За счет одностороннего крыла части плода (однокрылатки) под действием ветра переносятся на значительное расстояние от материнского растения.
Как уже было отмечено, гинецей у представителей сем. Aceraceae изучен недостаточно. Обычно указывается, что он состоит из 2 сросшихся плодолистиков (Prozhina, 1953; Khushalani, 1963; Takhtajan, 1966, 2009; Peck, Lersten, 1991a). Есть указания на то, что гинецей является синкарпным (Takhtajan, 1966, 2009). При исследовании гинецея у Acer saccharicum Marsh. ssp. nigrum [Michx. f.] Desm. (Peck, Lersten, 1991) было выявлено, что бикарпеллятный гинецей имеет короткий столбик, заканчивающийся лопастями рыльца. Рыльца покрыты папилловидными клетками, как и у изученного нами Acer ginnala. Также в этой работе было установлено, что сначала завязь двугнездная только в нижней части. Верхняя часть завязи остается не разделенной на гнезда и открывается в короткий столбик.
Результаты исследования, проведенного нами, во многом совпадают с данными C.J. Peck и N.R. Lersten (1991a). Однако мы смогли уточнить характер изменений, происходящих во время развития гинецея. На ранних стадиях развития нижняя часть завязи является 2-гнездной и образована синасцидиатной и закрытой симпликатной зонами. Последняя очень похожа по строению на синасцидиатную зону, которая, в отличие от закрытой симпликатной зоны, лишена семязачатков. В средней части завязи имеется открытая симпликатная зона, в центре которой формируется щель. Верхняя часть завязи и стилодии образованы асимпликатной зоной. В ходе развития в средней, а затем и верхней частях завязи происходит постгенитальное срастание плодолистиков, которое становится совершенным в средней и несовершенным в верхней частях. В результате этого гинецей становится полностью синкарпным и характеризуется наличием трех типов септ: базальной в нижней, медианной в средней и апикальной в верхней частях. Столбик не образуется, стилодии до конца остаются свободными, кроме дистальной части, где они объединяются в головчатое рыльце.
Тип семязачатка у видов рода Acer обсуждается (Alimova, 1985). У многих из них его описывают как анатропный: A. platanoides, A. campestre L., A. mono Max., A. pseudoplatanus, A. tataricum L., A. ginnala, A. semenovii Rgl. et Herd., A. negundo (Yudin, 1957), A. oblongum L. (Khushalani, 1963) A. saccharicum (Peck, Lersten, 1991a). Однако у A. platanoides было показано, что cемязачатки являются ортотропными, а не анатропными (Prozhina, 1953). Наши данные также не совпадают с результатами исследования, проведенного В.Г. Юдиным (Yudin, 1957). У A. ginnala семязачаток развивается не по анатропному типу. Сначала он похож на ортотропный, но в ходе развития происходит его смещение из латерального положения в центральное. С учетом удлинения фуникулуса и изгибания в области микропиле семязачаток у этого вида можно определить как геми-ортотропный. По-видимому, исходя из иллюстраций, приведенных в статье, семязачаток у A. saccharicum (Peck, Lersten, 1991a) также является геми-ортотропным. В отличие от A. ginnala он располагается не в трансверсальной, а в сагиттальной плоскости, при этом хорошо видно, что семязачаток занимает боковое положение и является асимметричным. Он находится на разросшейся плаценте, которая принята авторами за наружный интегумент со стороны рафе. При таком положении в завязи семязачаток не может быть анатропным. Клетки эпидермы плаценты A. saccharicum сильно вытянуты в радиальном направлении, и, вероятно, как и у A. ginnala, функционируют в качестве плацентарного обтуратора.
Из других особенностей отметим время и характер развития крыла с дорсальной стороны каждого плодолистика. В.Г. Юдин (Yudin, 1957) отмечает, что это происходит после оплодотворения, а разделительный слой между мерикарпиями образуется с момента формирования семени. И односторонние крылья, и разделительный слой начинают выявляться с ранних стадий развития гинецея, что было отмечено нами уже с начала мегаспорогенеза в семязачатке. Другое несовпадение с результатами этого автора заключается в том, что, по его мнению, второй семязачаток в каждом гнезде завязи не развивается из-за отсутствия его опыления. Как было показано в нашем исследовании, у A. ginnala каждый второй семязачаток в гнезде начинает дегенерировать еще до опыления во время формирования зародышевого мешка. Это связано с тем, что одна из плацент начинает разрастаться и оба семязачатка в гнезде, перекрывая друг друга, не могут разместиться в его пространстве. Такая особенность развития семязачатков связана с тем, что число заложившихся семязачатков не всегда способно реализоваться. В завязи на разных стадиях наблюдается их абортирование, что приводит к снижению числа семян в плоде и появлению мало- и односемянных плодов. Примеры видов растений из разных семейств (Araliaceae, Alismataceae, Brassicaceae, Euphorbiaceae, Menispermaceae, Paeonaceae, Rosaceae, Umbelliferae и др.) были приведены нами ранее в одной из обзорных статей (Shamrov, 2018).
Список литературы
[Alimova] Алимова Г.К. 1985. Cемейство Aceraceae. – В кн.: Сравнительная эмбриология растений. Т. 3. Л. С. 183–185.
[Aksyonova] Аксенова Н.А. 1975. Клены. М. 96 с.
[Gerasimov] Герасимов С. 2004. Оружие пролетариата. – В мире растений. 4: 10–12.
[Demyanova] Демьянова Е.И. 2010. Антэкология. Пермь. 116 с.
Hall B.A. 1951. The floral anatomy of the genus Acer. – Amer. J. Bot. 38 (10): 793–799.
Khushalani I. 1963. Floral morphology and embryology of Acer oblongum. – Phyton. 10 (3–4): 275–284.
[Pausheva] Паушева З.П. 1974. Практикум по цитологии растений. М. 288 с.
Peck C.J., Lersten N.R. 1991a. Gynoecial ontogeny and morphology, and pollen tube pathway in black maple, Acer saccharum ssp. nigrum (Aceraceae). – Amer. J. Bot. 78 (2): 247–259.
Peck C.J., Lersten N.R. 1991b. Papillate stigmas in Acer (Aceraceae). – Bull. Torrey Bot. Club. 118 (1): 20–23.
[Prozhina] Прозина М.Н. 1953. Эмбриологическое исследование клена остролистного (Acer platanoides L.) в связи с плохим плодоношением в условиях Калмыкии. – Бюл. Моск. общ-ва испыт. прир. Отд. биологии. 58 (2): 66–75.
[Pshennikova] Пшенникова Л. М. 2000. Анатомическое строение околоплодника семян семи видов кленов. – В кн.: Растения в природе и культуре. Владивосток. С. 182–184.
[Shamrov] Шамров И.И. 2013. Еще раз о типах гинецея покрытосеменных растений. – Бот. журн. 98 (5): 568–595.
[Shamrov] Шамров И.И. 2015. Эмбриология и воспроизведение растений. СПб. 200 с.
[Shamrov] Шамров И.И. 2017. Морфологические типы семязачатков цветковых растений – Бот. журн. 102 (2): 129–146. https://doi.org/https://doi.org/10.1134/S0006813618020011
[Shamrov] Шамров И.И. 2018. Особенности морфогенеза, разнообразие и возможные преобразования семязачатков цветковых растений. – Бот. журн. 103 (2): 163–186. https://doi.org/
[Yudin] Юдин В.Г. 1957. Анатомо-морфологические особенности развития плодов некоторых видов клена. – Бот. журн. 42 (2): 260–272.
[Takhtajan] Тахтаджян А.Л. 1966. Система и филогения цветковых растений. Л. 611 с.
Takhtajan A. 2009. Flowering plants. Springer. 871 p. https://doi.org/https://doi.org/10.1007/978-1-4020-9609-9
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал