

Ботанический журнал, 2019, T. 104, № 5, стр. 729-740
СТРУКТУРА И СОСТОЯНИЕ ПОПУЛЯЦИЙ ТРЕХ РЕДКИХ ВИДОВ РОДА HEDYSARUM (FABACEAE) НА ЮЖНОМ УРАЛЕ
Л. М. Абрамова 1, А. Н. Мустафина 1, *, О. А. Каримова 1, З. Х. Шигапов 1
1 Южно-Уральский ботанический сад-институт – обособленное структурное подразделение Уфимского федерального исследовательского центра РАН
450080 г. Уфа, ул. Менделеева, 195/3, Россия
* E-mail: alfverta@mail.ru
Поступила в редакцию 12.02.2019
После доработки 11.03.2019
Принята к публикации 12.03.2019
Аннотация
Представлены результаты изучения онтогенетической структуры ценопопуляций трех редких видов рода Hedysarum L. в Республике Башкортостан и Оренбургской области: H. grandiflorum Pall., H. argyrophyllum Ledeb. и H. gmelinii Ledeb. Плотность исследованных ценопопуляций от 1.7 до 15.1 экз./м2. Усредненный онтогенетический спектр, центрированный с максимумом на средневозрастных особях. Ценопопуляции варьируют от молодых до зрелых (H. grandiflorum и H. gmelinii) и от молодых до стареющих (H. argyrophyllum). Cостояние популяций исследованных видов на Южном Урале стабильное, дополнительных мер по охране в регионе не требуется. Для более малочисленного H. argyrophyllum возможно придание охраняемого статуса территориям с хорошо сохранившимися локалитетами.
По Южному Уралу проходят границы ареалов многих европейских, сибирских и азиатских видов растений, здесь они нередко представлены изолированными и фрагментированными локалитетами, и являются реликтами третичной и плейстоценовых флор (Kucherov at al., 1987). На границе ареалов популяции редких видов находятся зачастую в стрессовых условиях, обладают своеобразием внутренней организации, структуры и других биологических особенностей. Актуальная задача для этих видов растений – проведение инвентаризации местонахождений, оценка структуры и состояния популяций в различных условиях местообитаний и изучение их биологии и экологии, позволяющее понять причины их редкости.
Наши исследования посвящены изучению структуры и состояния ценопопуляций (ЦП) трех редких видов рода Hedysarum L. в Республике Башкортостан (РБ) и Оренбургской области (ОО).
В состав рода Hedysarum входят более 200 видов, распространенных во внетропических областях северного полушария, особенно многочисленных в Передней и Средней Азии (Flora…, 1996). В Республике Башкортостан встречается 5 видов рода, в Оренбургской области – 7 видов, большая часть которых включена в Красные книги этих регионов.
H. grandiflorum Pall. и H. argyrophyllum Ledeb. относятся к секции Subacaula (Boiss.) B. Fedtsch., а H. gmelinii Ledeb. – к секции Multicaula (Boiss.) B. Fedtsch. (Flora…, 1996). H. grandiflorum – восточноевропейский вид с довольно широким ареалом (Волжско-Камское и Волжско-Донское междуречье, Заволжье, Приуралье, часть Украины и Казахстана), включен в Красную книгу РФ с категорией 3 – редкий вид (Red…, 2008), охраняется в 11 регионах России (Red…, 2004(2005)), включая все регионы Южного Урала. H. argyrophyllum – южноуральский эндемичный вид (Knyazev, 2014), охраняется в Республике Башкортостан и в Челябинской области, также отнесен к категории 3 (Red…, 2004(2005); Red…, 2011). H. gmelinii – реликт сибирского происхождения, основной ареал которого простирается от гор Средней Азии и Алтая до Якутии и Монголии (Knyazev, 2014). На Южном Урале и Средней Волге расположен изолированный реликтовый фрагмент ареала. Вид охраняется в Республике Татарстан и Ульяновской области (Red…, 2004(2005)). Входит в список объектов растительного мира, которые нуждаются на территории Республики Башкортостан в особом внимании к их состоянию в природной среде и мониторинге. Ареалы всех 3-х видов на территории Южного Урала почти не пересекаются.
H. grandiflorum и H. argyrophyllum – близкородственные виды; последний является географической расой H. grandiflorum s. l. и замещает его к востоку от линии Мелеуз–Оренбург–Соль-Илецк (Knyazev, 2014). В ряде локалитетов по этой границе выявлены полиморфные по окраске цветков популяции.
Цель работы – изучение особенностей онтогенетической структуры и современного состояния ценопопуляций H. grandiflorum, H. argyrophyllum и Hedysarum gmelinii.
МАТЕРИАЛЫ И МЕТОДИКИ ИССЛЕДОВАНИЯ
Для проведения сравнительного анализа структуры и демографических параметров популяций видов рода Hedysarum в 2016 г. на территории ряда административных районов Предуралья Республики Башкортостан и Оренбургской области на градиенте Давлеканово–Соль-Илецк (450 км) обследованы 10 ценопопуляций (ЦП) H. grandiflorum, 6 ЦП H. argyrophyllum и 7 ЦП H. gmelinii. Две ЦП H. argyrophyllum обследованы в Зауралье (рис. 1). Название ЦП давалось по ближайшему к ней населенному пункту или географическому объекту. (табл. 1).
Рис. 1.
Схема расположения ценопопуляций изучаемых видов на территории Южного Урала.
Fig. 1. The location of the cenopopulations of the studied species in the South Urals.
H. grandiflorum (полиморфные по окраске) – H. grandiflorum (form polymorphic in colour)
H. grandiflorum (розовоцветковые формы) – H. grandiflorum (form with pink flowers)
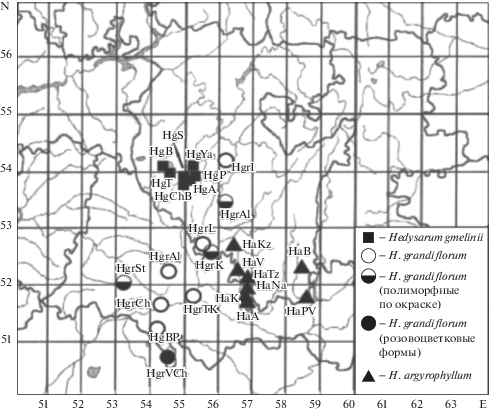
Таблица 1.
Характеристика природных местообитаний ЦП изучаемых видов рода Hedysarum. Table 1. Characteristics of the natural habitats of coenopopulations of studied species of the genus Hedysarum
| № ЦП № СР |
Местонахождение Location |
Местообитание Habitat |
Плотность, экз./м2 Density, ind./m2 |
Антропогенная нагрузка Anthropogenic load |
|---|---|---|---|---|
| H. grandiflorum | ||||
| 1 | д. Исмагилово 54°17792, 55°95766 | Средняя часть склона юго-западной экспозиции с уклоном 25°. Каменистость до 20% (известняковый щебень). Типчаково-тырсовая петрофитная степь | 6.6 | Сильная (выпас, рекреация) |
| 2 | г. Алебастровая 53°44675, 56°08250 | Средняя часть склона юго-западной экспозиции, с уклоном 20°. Почвы задернованы. Типчаково-тырсовая настоящая степь | 2.6 | Умеренная (рекреация) |
| 3 | г. Кумертау 52°79460, 55°79946 | Пологий склон (до 5°) южной экспозиции. Каменистость 30%. Пустынноовсецово-коржинскоковыльная петрофитная степь | 8.2 | Сильная (выпас) |
| 4 | д. Лена 52°80169, 55°61000 |
Склон юго-западной экспозиции с уклоном 20°. Почвы задернованы, имеются обнажения глин. Красноватоковыльно-пустынноовсецовая настоящая степь | 4.7 | Слабая (выпас) |
| 5 | д. Александровка 52°68783, 54°45135 | Склон южной экспозиции с уклоном до 5°. Почвы задернованы. Типчаково-тырсовая настоящая степь | 7.5 | Умеренная (выпас) |
| 6 | Старобелогорcкие меловые горы 51°84029, 54°07860 | Верхняя часть склона западной экспозиции с уклоном 10°. Почвы задернованы. Пустынноовсецово-типчаковая настоящая степь | 6.4 | Слабая (выпас) |
| 7 | д. Татарская
Карагала 51°93255, 55°12514 |
Южный склон оврага, с уклоном до 10°. Почвы задернованы. Типчаково-полынковая настоящая степь | 7.3 | Сильная (выпас, хозяйственная деятельность) |
| 8 | Чесноковские меловые горы 51°69117, 54°02935 | Верхняя часть склона северо-восточной экспозиции с уклоном 10°. Каменистость до 70% (меловой щебень). Типчаково-мордовниковая кальцефитная степь | 6.1 | Незначительная (рекреация) |
| 9 | д. Большая Песчанка 51°21739, 54°14879 | Средняя часть берегового обрыва юго-западной экспозиции с уклоном 30°. Почвы – загипсованные глины. Солянковиднополынная петрофитная степь | 15.7 | Слабая |
| 10 | Верхнечебендинские меловые горы 50°67782, 54°47114 | Верхняя часть склона северо-восточной экспозиции с уклоном 15°. Каменистость до 60% (меловой щебень). Пустынножитняково-мордовниковая кальцефитная степь | 6.6 | Слабая |
| H. argyrophyllum | ||||
| 11 | г. Кызлартау 52°80197, 56°52170 | Верхняя часть склона южной экспозиции с уклоном 20°. Каменистость до 10% (известняковый щебень). Пустынноовсецово-типчаковая петрофитная степь | 15.1 | Средняя (выпас) |
| 12 | г. Высокая, 52°23317, 56°47654 | Верхняя часть склона восточной экспозиции с уклоном 20°. Каменистость до 20% (известняковый щебень). Солянковидно-полынно-лессингоковыльная петрофитная степь | 4.4 | Слабая (выпас) |
| 13 | Тазларовские шишки 52°15989, 56°72584 |
Верхняя часть склона восточной экспозиции с уклоном 10°. Каменистость до 80% (известняковый щебень). Типчаковая петрофитная степь | 2.8 | Слабая (выпас) |
| 14 | д. Нижняя Акберда 52°00793, 56°75959 |
Верхняя часть склона юго-западной экспозиции с уклоном 10°. Каменистость до 90% (известняковый щебень). Тимьянно-типчаковая петрофитная степь | 4.3 | Слабая |
| 15 | ур. Кучтапкан 51°91214, 56°78196 | Верхняя часть склона юго-восточной экспозиции с уклоном 20°. Каменистость до 30% (известняковый щебень). Пустынноовсецово-красивейшековыльная настоящая степь | 1.9 | Слабая |
| 16 | д. Андреевка 51°69878, 56°89590 | Средняя часть склона южной экспозиции с уклоном 5°. Каменистость до 70% (известняковый щебень). Пустынноовсецово-красивейшековыльная настоящая степь | 2.8 | Слабая (выпас) |
| 17 | г. Балтатау 52°39919, 58°35927 | Вершина горы. Каменистость до 70% (породы метаморфического происхождения). Гвоздикоиглолистно-горноколосниковая петрофитная степь | 1.7 | Слабая (выпас) |
| 18 | Таштугайские
горы 51°87409, 58°53939 |
Верхняя часть склона западной экспозиции с уклоном 2°. Каменистость до 50% (породы метаморфического происхождения). Полынково-красноватоковыльная петрофитная степь | 3.5 | Слабая (выпас) |
| H. gmelinii | ||||
| 19 | д. Бурангулово 54°30446, 54°53505 | Нижняя часть склона западной экспозиции с уклоном 20°. Почвообразующие породы – песчаники. Тырсово-степномятликовая петрофитная степь | 12.2 | Средняя (выпас) |
| 20 | г. Таштюбе 54°20143, 54°71255 | Нижняя часть склона северо-западной экспозиции, с уклоном 20°. Почвообразующие породы – песчаники. Пустынноовсецово-коржинскоковыльная петрофитная степь | 1.7 | Слабая |
| 21 | г. Ярыштау 54°14697, 55°08565 | Средняя часть склона северо-западной экспозиции с уклоном 35°. Почвообразующие породы – песчаники. Пустынноовсецово-коржинскоковыльная петрофитная степь | 2.8 | Слабая (рекреация) |
| 22 | д. Алышево 54°04731, 55°06045 | Склон западной экспозиции с уклоном 5–10°. Почвообразующие породы – песчаники. Пустынноовсецово-коржинскоковыльная петрофитная степь | 1.8 | Средняя (выпас) |
| 23 | г. Пикарская 53°98509, 55°12591 | Верхняя часть склона западной экспозиции с уклоном 5–10°. Почвообразующие породы – песчаники. Пустынноовсецово-тырсовая петрофитная степь | 6.1 | Средняя (выпас) |
| 24 | г. Сатыртау 53°95939, 55°06461 | Нижняя часть склона восточной экспозиции с уклоном 20–25°. Почвообразующие породы – песчаники. Пустынноовсецово-коржинскоковыльная петрофитная степь | 12.2 | Средняя (выпас) |
| 25 | д. Чятай-Бурзян 53°80025, 54°82978 | Вершина склона юго-восточной экспозиции с уклоном 10–15°. Пустынноовсецово-тырсовая петрофитная степь | 10.0 | Средняя (выпас) |
Онтогенез видов H. grandiflorum и H. gmelinii описан В.Н. Ильиной (Ilyina, 2007, 2011), онтогенез H. argyrophyllum, как викарианта H. grandiflorum, протекает по сходному с ним типу и специально не изучался. Онтогенетическую структуру ценопопуляций вида в разных эколого-фитоценотических условиях обитания изучали методом трансект (Tsenopopulyatsii…, 1976). Для определения особенностей демографической структуры и плотности ЦП в каждой из них на трансекте закладывалось 25 пробных площадок размером 1 м2. Порядок заложения линейный или шахматный, и шаг трансекты (5 или 10 м) определялся площадью, занимаемой конкретной ценопопуляцией и проективным покрытием вида.
При определении онтогенетической структуры ЦП, согласно стандартным критериям (Rabotnov, 1950; Uranov, 1975; Tsenopopulyatsii…, 1976), учитывались следующие онтогенетические состояния: ювенильные (j), имматурные (im), виргинильные (v), молодые генеративные (g1), средние генеративные (g2), старые генеративные (g3), субсенильные (ss). На основании полученных данных построены онтогенетические спектры ЦП.
Для характеристики онтогенетической структуры ЦП применяли общепринятые демографические показатели: индекс восстановления (Zhukova, 1995) и индекс старения (Glotov, 1998). Для оценки состояния ЦП был применен критерий “дельта–омега” Л.А. Животовского (Zhivotovskii, 2001), основанный на совместном использовании индексов возрастности (∆) (Uranov, 1975) и эффективности (ω) (Zhivotovskii, 2001), на основании которых определялась принадлежность их к следующим типам: молодые, зреющие, зрелые, переходные, стареющие, старые.
Анализ данных провели в MS Excel 2010 c использованием стандартных показателей (Zaitsev, 1990).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ОБСУЖДЕНИЕ
Краткая характеристика местообитаний ЦП изучаемых видов (ЦП каждого вида располагаются по градиенту с севера на юг) приводится ниже.
Распределение особей по онтогенетическим состояниям и демографические показатели исследованных популяций представлены в таблице и на рисунке (табл. 2, рис. 2). По классификации А.А. Уранова и О.В. Смирновой (Uranov, Smirnova, 1969) изученные ЦП H. grandiflorum, H. argyrophyllum и H. gmelinii относятся к нормальным, большинство из них – неполночленные, ряд ЦП H. grandiflorum и H. gmelinii – полночленные. Отсутствие в спектрах большинства ЦП субсенильных и сенильных особей связано с сокращением онтогенеза за счет быстрого отмирания растений в старом генеративном состоянии: особи проходят полный онтогенез только в благоприятных условиях (при достаточной влажности почвы и отсутствии существенных антропогенных нарушений).
Таблица 2.
Распределение особей по онтогенетическим состояниям и демографические показатели ценопопуляций видов рода Hedysarum Table 2. Distribution of individuals on ontogenetic states and demographic indicators of coenopopulations of Hedysarum genus species
| Номер ЦП Number of coenopopulation |
*Онтогенетическое состояние, % *Ontogenetic state, % |
Демографические показатели Demographic indicators |
|||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| р | j | im | v | g1 | g2 | g3 | ss | ∆ | ω | Тип ЦП Coenopopulation type |
IВ IRec |
IСТ IAg |
|
| Hedysarum grandiflorum | |||||||||||||
| 9 | 6.9 | 51.3 | 14.3 | 9.7 | 5.4 | 8.2 | 2.6 | 1.8 | 0.12 | 0.26 | молодая young |
4.68 | 0.02 |
| 5 | 5.3 | 36.9 | 11.2 | 8.0 | 7.5 | 27.8 | 3.2 | 0.0 | 0.20 | 0.44 | « | 1.46 | 0.00 |
| 4 | 6.0 | 11.1 | 16.2 | 27.4 | 6.8 | 30.8 | 1.7 | 0.0 | 0.23 | 0.53 | « | 1.39 | 0.00 |
| 1 | 0.6 | 23.6 | 18.8 | 11.5 | 11.5 | 28.5 | 5.5 | 0.0 | 0.24 | 0.52 | « | 1.19 | 0.00 |
| 3 | 0.5 | 19.5 | 17.6 | 16.1 | 13.2 | 30.2 | 2.9 | 0.0 | 0.24 | 0.54 | « | 1.15 | 0.00 |
| 10 | 1.8 | 12.1 | 13.3 | 30.9 | 9.7 | 25.5 | 4.2 | 2.4 | 0.25 | 0.54 | « | 1.43 | 0.02 |
| 6 | 4.2 | 27.2 | 15.7 | 3.7 | 7.9 | 40.8 | 0.5 | 0.0 | 0.25 | 0.54 | « | 0.95 | 0.00 |
| 8 | 0.0 | 14.4 | 21.6 | 20.9 | 5.2 | 34.6 | 3.3 | 0.0 | 0.25 | 0.55 | « | 1.32 | 0.00 |
| 7 | 15.8 | 17.5 | 15.8 | 7.1 | 4.9 | 26.2 | 9.3 | 3.3 | 0.26 | 0.46 | « | 1.00 | 0.04 |
| 2 | 0.0 | 1.5 | 9.1 | 7.6 | 7.6 | 66.7 | 7.6 | 0.0 | 0.42 | 0.84 | зрелая mature |
0.22 | 0.00 |
| Hedysarum argyrophyllum | |||||||||||||
| 14 | 0.0 | 15.7 | 7.4 | 35.2 | 5.6 | 26.9 | 6.5 | 2.8 | 0.24 | 0.55 | молодая young |
1.50 | 0.03 |
| 13 | 0.0 | 0.0 | 19.7 | 39.4 | 9.9 | 14.1 | 12.7 | 4.2 | 0.28 | 0.54 | « | 1.62 | 0.04 |
| 18 | 0.0 | 4.5 | 21.6 | 20.5 | 10.2 | 43.2 | 0.0 | 0.0 | 0.28 | 0.64 | зреющая ripening |
0.87 | 0.00 |
| 11 | 0.0 | 16.7 | 12.2 | 14.1 | 8.2 | 43.8 | 5.0 | 0.0 | 0.30 | 0.63 | « | 0.75 | 0.04 |
| 12 | 0.0 | 5.5 | 14.7 | 20.2 | 1.8 | 27.5 | 19.3 | 11.0 | 0.41 | 0.60 | переходная transitional |
0.83 | 0.11 |
| 16 | 0.0 | 0.0 | 5.8 | 20.3 | 2.9 | 58.0 | 8.7 | 4.3 | 0.43 | 0.78 | зрелая mature |
0.38 | 0.04 |
| 15 | 0.0 | 0.0 | 0.0 | 12.5 | 6.3 | 81.3 | 0.0 | 0.0 | 0.44 | 0.91 | « | 0.14 | 0.00 |
| 17 | 0.0 | 0.0 | 2.4 | 0.0 | 0.0 | 38.1 | 31.0 | 28.6 | 0.67 | 0.75 | стареющая aging |
0.03 | 0.29 |
| Hedysarum gmelinii | |||||||||||||
| 19 | 14.5 | 30.6 | 13.2 | 11.5 | 4.3 | 20.4 | 4.3 | 1.3 | 0.18 | 0.37 | молодая young |
1.91 | 0.02 |
| 24 | 48.8 | 6.2 | 4.6 | 5.6 | 7.2 | 23.3 | 4.3 | 0.0 | 0.18 | 0.37 | « | 0.47 | 0.00 |
| 25 | 11.6 | 27.3 | 20.1 | 7.6 | 2.8 | 29.7 | 0.8 | 0.0 | 0.19 | 0.42 | « | 1.65 | 0.00 |
| 23 | 30.1 | 13.7 | 9.1 | 4.6 | 1.3 | 32.0 | 7.8 | 1.3 | 0.25 | 0.45 | « | 0.67 | 0.02 |
| 21 | 0.0 | 14.5 | 5.8 | 5.8 | 4.3 | 60.9 | 8.7 | 0.0 | 0.39 | 0.76 | зрелая mature |
0.35 | 0.00 |
| 22 | 0.0 | 4.5 | 0.0 | 13.6 | 13.6 | 68.2 | 0.0 | 0.0 | 0.39 | 0.85 | « | 0.22 | 0.00 |
| 20 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 88.1 | 7.1 | 2.4 | 0.52 | 0.97 | « | 0.00 | 0.02 |
*Примечание. Онтогенетическое состояние: j – ювенильное, im – имматурное, v – виргинильное, g1 – молодое генеративное, g2 – средневозрастное генеративное, g3 – старое генеративное, ss – субсенильное.*Note. Оntogenetic state: j – juvenile, im – immature, v – virginile, g1 – young generative, g2 – mature generative, g3 – old generative, ss – subsenile
Рис. 2.
Усредненные онтогенетические спектры ценопопуляций трех видов рода Hedysarum. По оси х: – онтогенетическое состояние: j – ювенильное, im – имматурное, v – виргинильное, g1 – молодое генеративное, g2 – средневозрастное генеративное, g3 – старое генеративное, ss – субсенильное; по оси у – доля особей данного онтогенетического состояния, %.
Fig. 2. The average ontogenetic spectrums of coenopopulations of three Hedysarum species. X-axis: ontogenetic state: j – juvenile, im – immature, v – virginile, g1 – young generative, g2 – mature generative, g3 – old generative, ss – subsenile; Y-axis: the proportion of individuals of this ontogenetic state, %.
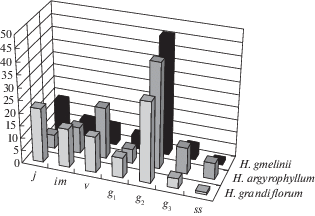
Онтогенетическая структура конкретных ЦП видов рода Hedysarum имеет два типа спектра – левосторонний и центрированный, в различной степени отличающиеся от усредненного. Это зависит от экологических условий обитания, степени антропогенной нагрузки и колебаний погодных условий, которые влияют на особенности прорастания семян и темпы развития особей в том или ином онтогенетическом состоянии.
В ЦП 7, 9 и 10 H. grandiflorum (Оренбургская обл.) онтогенетические спектры полночленные, в остальных популяциях отсутствуют проростки и/или постгенеративная фракция, и они являются неполночленными. Левосторонний спектр формируется в ЦП 5, 9 и 10, где максимум приходится на ювенильные или виргинильные особи (36.9, 51.3 и 30.9% соответственно). Эти популяции находятся в местах с невысоким проективным покрытием и небольшой плотностью травостоя, что позволяет прорастать семенам и развиваться молодым особям. В большинстве ценопопуляций H. grandiflorum формируется центрированный спектр, с абсолютным максимумом на средневозрастных генеративных особях (26.2–66.7%). Вероятно, это обусловлено каменистостью и сухостью субстрата, водной и ветровой эрозией, что приводит к элиминации особей на ранних этапах развития. В ЦП 1 и 4 количество ювенильных или виргинильных и средневозрастных генеративных особей примерно одинаково (23.6 и 28.5; 27.4 и 30.8% соответственно).
Все ЦП H. argyrophyllum являются неполночленными, в них отсутствуют проростки. В ЦП 13 формируется левосторонний одновершинный спектр, с максимумом на виргинильных особях (39.4%), левосторонний двухвершинный – в ЦП 14, один пик приходится на виргинильные особи (35.2%), второй – на средневозрастные генеративные особи (26.9%). Остальные популяции характеризуются центрированным спектром с преобладанием средневозрастных генеративных особей (27.5–813%). Незначительно представлены особи молодой фракции или они полностью отсутствуют. Произрастание особей в верхней части склона способствует периодическому смыву семян весенними водами, а резкое пересыхание субстрата в жаркие периоды года приводит к гибели молодых растений, что отрицательно влияет на прорастание семян и способствует выпадению этих стадий.
Полный набор онтогенетических состояний ЦП H. gmelinii представлен в ЦП 19 и 23. В остальных случаях наблюдаются различные отклонения. Левосторонний одновершинный спектр формируется в ЦП 24, максимум приходится на проростки (48.8%). Левосторонний двухвершинный спектр выявлен в ЦП 19, один пик приходится на ювенильные особи (30.6%), второй – на средневозрастные генеративные особи (20.4%). Эти популяции находятся в умеренно нарушенных выпасом местах, с разреженным травостоем, что способствует прорастанию семян и развитию молодых особей.
В остальных популяциях формируется центрированный онтогенетический спектр, с максимумом на средневозрастных генеративных особях (29.7–88.1%). В ЦП 20, 21 и 22 формируется центрированный спектр, с абсолютным максимумом на средневозрастных генеративных особях (60.9–88.1%). ЦП 20 характеризуется полным отсутствием прегенеративной фракции. Возможно, это связано с тем, что при отсутствии постоянных нарушений в степных сообществах наблюдается старение популяций вследствие слабого возобновления из-за задернения почвы. В ЦП 23 и 25 онтогенетический спектр имеет второй пик, который приходится в ЦП 23 на проростки (30.1%), в ЦП 25 – на ювенильные особи (27.3%).
Оценка исследованных ценопопуляций видов рода Hedysarum по классификации “дельта–омега” (табл. 2) показала, что почти все ЦП H. grandiflorum являются молодыми (∆ = 0.12–0.26, ω = 0.26–0.46). В основном это ЦП, где хорошо представлено возобновление, и преобладают молодые особи. ЦП 2 относится к зрелым (∆ = 0.42, ω = 0.84), где максимум приходится на средневозрастные генеративные особи. Эта ЦП произрастает вблизи крупного населенного пункта в условиях антропогенного воздействия (рекреация). Проведено также сравнение индексов восстановления (IВ) и старения (IСТ), отражающих динамические процессы популяции. В большинстве ЦП индекс восстановления (IВ) выше единицы (1.00–4.68), что говорит о высоком уровне пополнения молодыми особями и преобладании прегенеративной фракции. Индекс старения в большинстве ЦП равен нулю, поскольку отсутствуют постгенеративные особи. В ЦП 7, 9 и 10 индекс старения близок к нулю (0.02–0.04), это связано с тем, что большая часть особей отмирает в старом генеративном или субсенильном состоянии.
Ценопопуляции H. argyrophyllum варьируют от молодых до стареющих. Две ЦП 13 и 14 относятся к молодым (∆ = 0.24; 0.28, ω = 0.55; 0.54), здесь преобладает прегенеративная фракция, ЦП 18 и 11 – зреющие (∆ = 0.28. 0.30, ω = 0.64, 0.63), за счет накопления особей прегенеративного периода и средневозрастных генеративных особей, и полного отсутствия постгенеративной фракции. ЦП 12 относится к переходной (∆ = 0.41, ω = 0.60), в которой также наблюдается накопление прегенеративных особей, но уже значительна доля старых и постгенеративных растений. ЦП 16 и 15 являются зрелыми (∆ = 0.43; 0.44, ω = 0.78; 0.91). ЦП 17 относится к стареющей (∆ = 0.67; ω = 0.75) со значительной долей старых генеративных и субсенильных особей. Сравнение индекса восстановления и старения показало, что у большинства ЦП индекс восстановления ниже единицы (0.03–0.87). Индекс старения равен или близок нулю (0.00–0.29), это связано с тем, что большая часть особей отмирает в старом генеративном состоянии.
ЦП H. gmelinii (19, 23–25) относятся к молодым (∆ = 0.18–0.25, ω = 0.37–0.45). В них преобладают прегенеративные особи. ЦП 20–22 – зрелые (∆ = 0.39–0.52, ω = 0.76–0.97), в составе которых доля средневозрастных генеративных особей велика, а доля прегенеративных мала. Индекс восстановления в ЦП 19 и 25 выше единицы (1.65 и 1.91 соответственно), в остальных значительно ниже (0.22–0.67), а в ЦП 20 он равен нулю, что свидетельствует о полном отсутствии прегенеративной фракции и об интенсивном отмирании особей в старом генеративном состоянии.
ЗАКЛЮЧЕНИЕ
Большая часть исследованных ЦП отличаются довольно высокой плотностью и неполночленным онтогенетическим спектром, связанным с быстрым отмиранием растений после завершения генеративного состояния. Состояние популяций, по нашим наблюдениям, не зависит от типа растительности, но связано с антропогенными нагрузками. При этом небольшие нагрузки способствуют улучшению прорастания семян и развитию проростков, поскольку вызывают разрежение травостоя. При сильных нагрузках плотность популяций снижается.
Типичные (усредненные) онтогенетические спектры исследованных видов сходны – они центрированные с выраженным максимумом на средневозрастных генеративных растениях. Видовые отличия связаны с условиями произрастания: так у H. grandiflorum, произрастающего в наиболее благоприятных условиях и в целом являющегося видом с более широким распространением, в спектрах высока доля прегенеративных растений и возобновление хорошее. Два других вида отличаются меньшим пополнением молодыми растениями: как у H. argyrophyllum, растущего в наиболее неблагоприятных засушливых условиях крайнего юга и юго-востока региона, так и у H. gmelinii, произрастающего в сообществах с более плотным травостоем, препятствующим прорастанию семян и развитию молодых растений. По классификации “дельта–омега” наибольшее разнообразие ЦП (от молодых до стареющих) отмечено у H. argyrophyllum, два других вида представлены преимущественно молодыми, реже зрелыми ЦП. Сходные данные получены нами при сравнении ЦП H. grandiflorum Республики Башкортостан с таковыми Средней Волги (Abramova et al., 2016): республиканские популяции, расположенные на границе ареала, преимущественно молодые, тогда как в ЦП центральной части ареала в Самарской области преобладают зрелые популяции.
В целом следует отметить, что все три вида на Южном Урале представлены довольно большим числом достаточно крупных по численности популяций. Наиболее многочисленные и с высокой плотностью ЦП принадлежат H. grandiflorum, охраняемого на федеральном уровне, в которых отмечено хорошее возобновление. ЦП H. argyrophyllum, особенно в Зауральской части ареала, по нашим данным, отличаются более слабым возобновлением, низкой плотностью и нередко малочисленностью. Для этого вида необходим постоянный контроль за их состоянием и придание статуса ООПТ ряду хорошо сохранившихся локалитетов (например, в Республике Башкортостан – горам Высокая, Балтатау, Тюлькюлитау и др.). Хотя H. gmelinii не требует дополнительных мер по охране, необходим регулярный мониторинг природных популяций этого вида на территории Республики.
Список литературы
Abramova L.M., Karimova O.A., Mustafina A.N. 2014. Characteristic of coenopopulations of a rare species Hedysarum grandiflorum Pall. in stony steppes of the Cis–Urals. – Italian Sci. Rev. 2 (11): 241–244.
[Abramova et al.] Абрамова Л.М., Ильина В.Н., Каримова О.А., Мустафина А.Н. 2016. Сравнительный анализ структуры популяций Hedysarum grandiflorum (Fabaceae) в Самарской области и Республике Башкортостан. – Растит. ресурсы. Т. 52. № 2. С. 225–239.
[Flora…] Флора Восточной Европы. 1996. том IX. СПб. 456 с.
[Glotov] Глотов Н.В. 1998. Об оценке параметров возрастной структуры популяций растений. – В кн.: Жизнь популяций в гетерогенной среде. Ч. 1. Йошкар-Ола. С. 146–149.
[Ilyina] Ильина В.Н. 2007. Онтогенез копеечника крупноцветкового (Hedysarum grandiflorum Pall.). – В кн.: Онтогенетический атлас растений. Т. V. Йошкар-Ола. С. 126–132.
[Ilyina] Ильина В.Н. 2011. Онтогенез копеечника Гмелина (Hedysarum gmelinii Ledeb.). – В кн.: Онтогенетический атлас растений. Т. VI. Йошкар-Ола. С. 102–107.
[Knyazev] Князев М.С. 2014. Бобовые (Fabaceae Lindl.) Урала: видообразование, географическое распространение, историко-экологические свиты: Дис. …докт. биол. наук. СПб. 607 с.
[Kucherov et al.] Кучеров Е.В., Мулдашев А.А., Галеева А.Х. 1987. Охрана редких видов растений на Южном Урале. М. 204 с.
[Rabotnov] Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах. – В кн.: Тр. БИН АН СССР. Сер. 3. Геоботаника. Т. 6. М.; Л. С. 7–204.
[Red…] Красная книга Республики Башкортостан. 2011. Т. 1. Растения и грибы. Уфа. 384 с.
[Red…] Красная книга Российской Федерации (растения и грибы). 2008. М. 855 с.
[Red…] Красный список особо охраняемых редких и находящихся под угрозой исчезновения животных и растений. 2004(2005). Ч. 3.1 (Семенные растения). М. 352 с.
[Tsenopopulyatsii…] Ценопопуляции растений (очерки популяционной биологии). 1988. М. 181 с.
[Uranov] Уранов А.А. 1975. Возрастной спектр фитоценопопуляции как функция времени и энергетических волновых процессов. – Биол. науки. 2: 7–34.
[Uranov, Smirnova] Уранов А.А., Смирнова О.В. 1969. Классификация и основные черты развития популяций многолетних растений. – Бюлл. МОИП. Отд. биол. 79(1): 119–135.
[Zaitsev] Зайцев Г.Н. 1990. Математика в экспериментальной биологии. М. 296 с.
[Zhivotovskii] Животовский Л.А. 2001. Онтогенетическое состояние, эффективная плотность и классификация популяций. – Экология. 1: 3–7.
[Zhukova] Жукова Л.А. 1995. Популяционная жизнь луговых растений. Йошкар-Ола. 224 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал