

Ботанический журнал, 2019, T. 104, № 6, стр. 835-858
ПЕРИСТОМ МХОВ: ПРОБЛЕМЫ РАЗВИТИЯ И ПРОБЛЕМЫ ТЕРМИНОЛОГИИ
1 Московский государственный университет им. М.В. Ломоносова, биологический факультет
119234 Москва,
Ленинские Горы, д. 1, стр. 12, Россия
2 Главный ботанический сад им. Н.В. Цицина
127276 Москва, ул. Ботаническая, 4, Россия
* E-mail: misha_ignatov@list.ru
Поступила в редакцию 18.04.2019
После доработки 14.05.2019
Принята к публикации 16.05.2019
Аннотация
Рассмотрена история становления представлений о строении перистома, особенностях его развития и современное состояние этого вопроса. Терминология, введенная для описания строения перистома и связанная с функциональными особенностями отдельных слоев клеток, нуждается в уточнении, поскольку выявляющиеся исключения показывают большее разнообразие вариантов строения, чем было выявлено ранее. Фундаментальная закономерность формирования элементов перистома из периклинальных клеточных стенок трех соседних слоев клеток в целом остается неизменной, и названия слоев не требуют переименования, хотя их определение, данное изначально, требует корректировки. Особо обсуждается перистом Buxbaumia, который не следует считать переходным между нематодонтным и артродонтным, как это обычно представляется в современной литературе.
В отличие от животных, растения ведут прикрепленный образ жизни, поэтому, решение проблем, связанных с расселением, имеет для них особое значение. Семенные растения широко задействуют перенос диаспор с помощью животных, тогда как для споровых растений это второстепенный метод распространения. Возможно, зоохория у них пока остается сильно недооцененной; в частности, работы последних лет все чаще представляют сенсационные данные о переносе мхов и птицами на тысячи километров, и млекопитающими (Lewis et al., 2014; Chmielewski, Eppley, 2019). Но все же основным способом распространения спор мхов можно считать анемохорию; при этом их высвобождение из коробочек осуществляется дозированно, в благоприятных условиях, что регулируется гигроскопическими движениями перистома – специального органа, свойственного только мхам, состоящего из зубцов, расположенных по краю устья коробочки и большей частью прикрывающих его после сбрасывания крышечки.
Утверждение об исключительно важной роли перистома в расселении мхов основано на общих соображениях здравого смысла, как и заключение о том, что цветок покрытосеменных имеет основным предназначением привлечение насекомых. При всех исключениях вряд ли кто-то возьмется с этим спорить. Говоря о столь же важной роли перистома для рассеивания спор мхов, мы основываемся как на огромном разнообразии его строения, так и на параллелизмах в его преобразовании в разных эволюционных линиях при попадании видов в другие условия существования. Особенно хорошо известны параллелизмы при переходе к эпифитизму (Huttunen et al., 2004, 2012). И, конечно же, перистом для систематики мхов имеет не меньшее значение, чем цветок для систематики покрытосеменных. Как классификация покрытосеменных основывается преимущественно на признаках цветка со времен Линнея, так и система мхов в работе Й. Гедвига (Hedwig, 1801), с которой берет начало официальная номенклатура мхов, подразделяла мхи на не имеющие перистома, с простым перистомом и с двойным перистомом. Таким образом, центральное положение перистома в науке о разнообразии мхов – бриологии очевидно.
Работ, представляющих детальное изучение перистома, было относительно немного, причем они не полностью совпадают в своей терминологии и акцентах, что усложняет задачу данного обзора: с одной стороны, требуется описать строение перистома, с другой – показать эволюцию представлений о его строении и изменение смысла отдельных терминов, применяемых для его описания. Сначала мы дадим общее, сравнительно обобщенное представление о строении наиболее распространенного у мхов артродонтного перистома, затем рассмотрим работы, на основе которых сформировалось это представление, далее остановимся на последних работах, их результатах и вопросах необходимости уточнения терминологии, и, наконец, отметим проблемы, которые не могут считаться окончательно решенными и требуют дополнительного изучения. На протяжении всего обзора будет много отступлений, касающихся развития представлений о естественной системе мхов, поскольку именно систематики часто больше писали о перистоме, нежели анатомы: например, в классическом учебнике бриологии 1932 года под редакцией Ф. Вердоорна в главе Х. Диксона о системе мхов (Dixon, 1932) про перистомы написано больше, чем в разделе о морфологии и анатомии мхов (Wijk, 1932). Практически все группы, обсуждаемые в данном обзоре, описаны и проиллюстрированы во “Флоре мхов средней части Европейской России”11 и “Флоре мхов России”22,33 (Ignatov, Ignatova, 2003; Ignatov et al., 2017, 2018a).
1. ОБЩИЕ ЗАКОНОМЕРНОСТИ РАЗВИТИЯ ПЕРИСТОМА
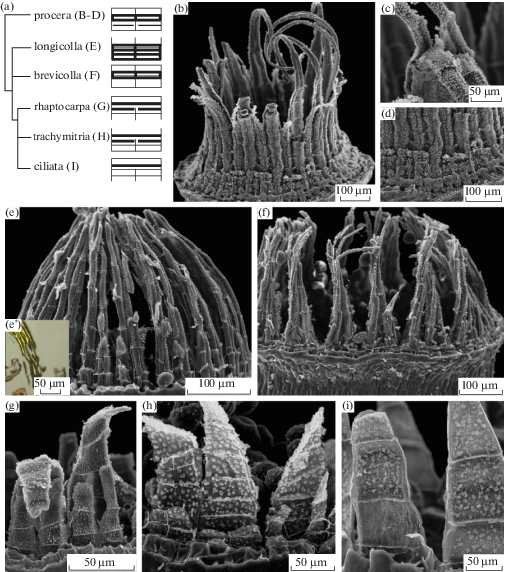
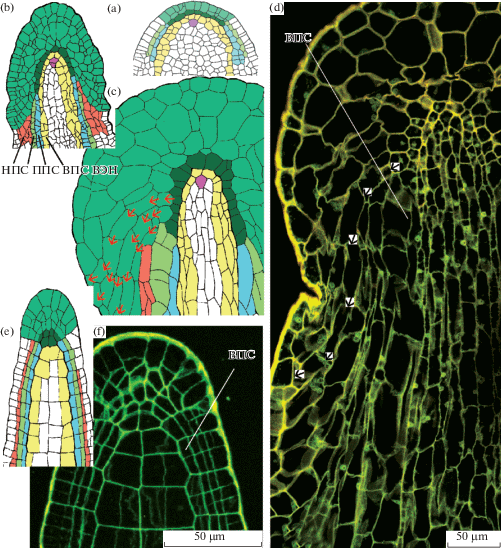
Развитие спорофита мхов изучают на растениях на последовательных стадиях развития, которые, однако, в известной степени можно наблюдать в пределах одного ювенильного спорофита на серии срезов. Рисунок 1 показывает три варианта представления стадий развития, позволяющие составить 3D реконструкцию (рис. 1C) по реально наблюдаемым срезам (рис. 1A, B).
Рис. 1.
Последовательные стадии развития спорофита, показанные на фотографиях анатомических срезов Encalypta longicolla (A) и Funaria arctica (D) (по Ignatov et al., 2018с); рисунках срезов Arrhenopterum heterostichum (B) (по Blomquist, Robertson, 1941, схематично); схеме объемной реконструкции развития спорофита Polytrichum juniperinum (С) (по Wenderoth, 1941, схематично). A – продольный срез с обозначением уровней, на которых сделаны поперечные срезы A1–A6; B1–B8 – поперечные срезы; B9 – продольный срез; схематичные изображения C1 апикальной и С2–С5 субапикальных клеток, показывающие их форму и порядок делений; D – поперечный срез на стадии разделения зубцов между собой (между клетками превичного перистомного слоя (ППС) появляются полости (стрелки)). ВПС, НПС – внутренний, наружный перистомные слои соответственно.
Fig. 1. Sequentional stages of sporophyte development, shown in micrographs of serial sections of Encalypta longicolla (A) and Funaria arctica (D) (after Ignatov et al., 2018с); serial sections of Arrhenopterum heterostichum (B) (schematically redrawn from Blomquist, Robertson, 1941), and Polytrichum juniperinum (C) reconstruction (schematically redrawn from Wenderoth, 1941). A – longitudinal section showing position of sections in A1–A6; B1–B8 – transverse sections; B9 – longitudinal section; schemes of apical (C1), and subapical (С2–С5) cells showing their shape and sequence of further divisions; D – transverse section at early stage of peristome tееth formation: spaces between cells of primary peristomial layer (PPL) arrowed. ВПС, ППС, НПС: inner, primary and outer peristomial layers (IPL, PPL and OPL) respectively.
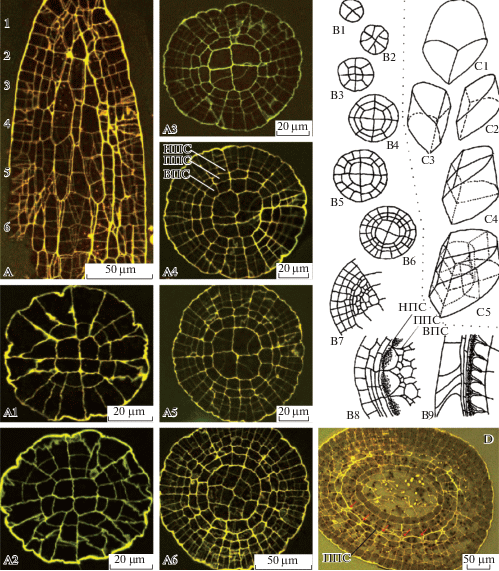
Уже после нескольких делений зиготы апикальная клетка спорофита приобретает широко клиновидное основание, задающее правильное расположение клеток в дистальной части спорофита (рис. 1С). На сериях поперечных срезов под апикальной клеткой спорофита можно видеть срез округлой формы, поделенный сначала на две, а затем на 4 равные клетки, выглядящие на поперечном срезе как четверти круга, разделенного крестом (рис. 1 B1, C1–4). Далее по мере развития (чему в определенной мере соответствуют срезы ниже у образцов на более поздних стадиях развития) периклинальные деления приводят к восьмиклеточному на поперечном срезе строению (рис. 1 B3). Четыре центральные клетки этой стадии, называемые эндотецием, дают впоследствии ниже уровня устья коробочки спорогенную ткань и колонку, а из четырех периферических, называемых амфитецием, развиваются: в верхней части коробочки – перистом и крышечка, а ниже устья – наружная стенка спорового мешка, структуры воздушной полости и стенка урночки.
В формировании типичного артродонтного перистома из 16 зубцов (рис. 1 и 2), представленного примерно у 90% видов мхов, принимают участие 3 внутренних, прилегающих к эндотецию, концентрических слоя клеток, называемые: внутренний перистомный слой (ВПС), первичный перистомный слой (ППС) и наружный перистомный слой (НПС). Клетки этих трех слоев, как правило, разрушаются, но не целиком, а с сохранением периклинальных стенок, так что наружная стенка ВПС срастается с внутренней стенкой ППС, а наружная стенка ППС срастается с внутренней стенкой НПС, с образованием двух кругов элементов перистома – эндостома и экзостома (еще раз оговоримся, что речь идет в данном случае о типичном случае развития двойного артродонтного перистома). Схема сохраняющихся и разрушающихся частей клеток этих слоев, представленная на рис. 2, иллюстрирует развертку 1/8 части среза (соответствуя, таким образом, двум из 16 зубцов перистома). Схема позволяет видеть гомологичность зубцов простого перистома Dicranidae элементам эндостома (внутреннего круга двойного перистома) других подклассов, хотя зубцы простого перистома Dicranidae внешне сходны с зубцами экзостома Funariidae и Bryidaea и совсем не похожи на элементы их эндостома (рис. 3). То, что зубцы в разных группах мхов не всегда гомологичны друг другу, отмечали уже авторы в XIX веке, обратившие внимание на то, что слой, формирующий наружный споровый мешок, продолжается у Dicranum в зубец простого перистома, тогда как у Bryum и Funaria этот же слой продолжается в слой, формирующий тонкий пленчатый эндостом (Lantzius-Beninga, 1850, цит. по Maier, Price, 2014), см. схемы справа на рис. 2. В сущности тот же вывод сделал Г. Филибер (Philibert, 1884a,b, цит. по Taylor, 1962), отметивший у Dicranum и близких видов отсутствие на наружной стороне зубцов срединной линии (рис. 3, Dicranum), которая есть у мхов с двойным перистомом (рис. 3, Bryum).
Рис. 2.
Схема трех основных типов строения артродонтных перистомов, характерных для трех подклассов класса Bryopsida. ВПС, ППС, НПС – перистомные слои: внутренний, первичный, наружный соответственно. Левый ряд – развертки 1/8 среза развивающегося спорофита, где пунктиром показаны разрушающиеся стенки клеток перистомных слоев, а жирными линиями – сохраняющиеся стенки, образующие зубцы; жирными стрелками – места, где соответствующий круг перистома расщеплен до основания, тонкими стрелками – расщеплен только вверху. Правее показан вид снаружи: соответствующих зубцов, где элементы экзостома даны жирными линиями, эндостома – тонкими линиями, границы клеток ВПС (не видимые снаружи) – пунктиром. Крайний правый ряд: продольные срезы нижних частей зубцов перистома и верхушки спорового мешка (отмечен звездочкой), иллюстрирующие продолжение ВПС в наружную стенку спорового мешка (схематичная перерисовка части иллюстраций Ланциуса-Бенинга, по Maier, Price, 2014) для родов Bryum, Dicranum и Funaria.
Fig. 2. Scheme of three main types of arthrodontous peristomes of three subclasses of Bryopsida. ВПС, ППС, НПС: inner, primary and outer peristomial layers (IPL, PPL and OPL) respectively. Left-median column shows 1/8 sector of sporophyte transverse sections, where dash-line: cell walls decomposed during maturation; heavy lines: retaining and incrassating cell walls; heavy arrows: places of splits to the base; thin arrows: places of splits only distally. Right-median column shows teeth from outside (heavy lines for exostome, thin for endostome, dash lines for IPL cell borders not seen from outside). Right column of longitudinal sections through bases of adult peristomes of Funaria, Dicranum, and Bryum, shows that the layer forming the outer spore sac continues into inner amphithecial layer and plays different role in peristome formation, thus “teeth” of Dicranum and “teeth” of Bryum and Funaria are not homologues (schematically redrawn from Lantzius-Beninga, 1850, cited from Maier, Price, 2014).
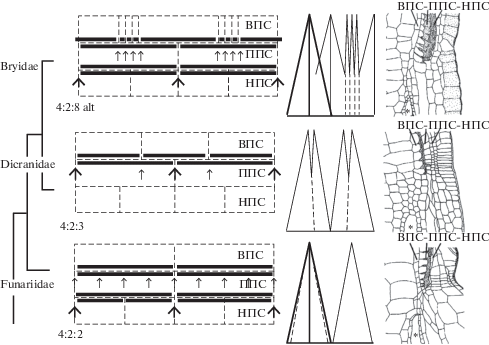
Рис. 3.
Филогенетическое древо мхов, показывающее положение порядков в системе и примеры строения перистомов.
Fig. 3. Phylogenetic tree of mosses, showing systematic position of orders and peristome structures.
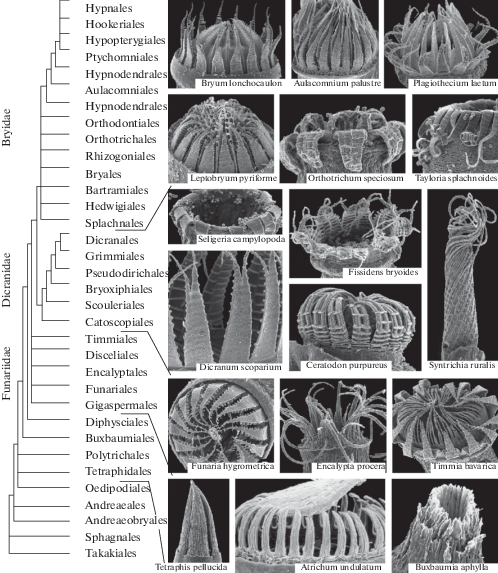
Несмотря на сходство в развитии перистомов (рис. 1 и 2), их структурное разнообразие очень велико (рис. 3). Следует также отметить, что число зубцов перистома у большинства мхов одно и то же – 16. Еще одна общая закономерность в строении перистома состоит в том, что у двойных перистомов элементы наружного круга, или зубцы, если они нормально развитые, всегда более мощные, гигроскопические, а элементы внутреннего круга (сегменты и реснички, обычно сидящие на базальной мембране) нежные, прозрачные. Принадлежность основных типов строения перистомов к тем или иным эволюционным линиям показана на рис. 2 и 3. Мы вынуждены в данном случае несколько забежать вперед, используя самые последние данные о филогении мхов, как для простоты изложения, так и потому, что они не раз публиковались в обзорах последних десятилетий (Ignatov, Ignatova, 2003; Ignatov et al., 2017) и, возможно, имеющееся у читателя представление о перистоме базируется именно на них. И, наконец, имея определенное представление о разнообразии объекта, проще оценить все превратности процесса формирования данного представления.
2. ИСТОРИЯ ИЗУЧЕНИЯ И СТАНОВЛЕНИЕ СОВРЕМЕННОГО ПРЕДСТАВЛЕНИЯ ОБ ОСНОВНЫХ ТИПАХ СТРОЕНИЯ ПЕРИСТОМОВ
Й. Гедвиг, как уже отмечено выше, положил перистом в основу классификации мхов, причем строение перистома было отражено им в названиях родов: например, Tetraphis (с четырьмя зубцами), Fissidens (с расщепленными зубцами), и т.д. (Hedwig, 1801). В последующих работах систематиков мхов первой трети XIX века названия родов на основе признаков перистома умножились, причем некоторые из них, в частности, Anomodon и Antitrichia явно отражают слабые возможности оптики того времени: первое название означает “с аномальным перистомом”, поскольку В.Д. Гукер и Т. Тэйлор ошибочно определили положение сегментов эндостома как расположенных между зубцами экзостома, а не во внутреннем круге; во втором случае С. Бридель счел расположение зубцов экзостома и сегментов эндостома супротивным, что также не соответствует действительности. Совершенствование оптики и ее доступность привели к тому, что середина XIX века стала богатой на замечательные открытия; в первую очередь следует вспомнить работу В. Гофмейстера по чередованию поколений у растений (Hofmeister, 1851). В бриологии же в то время были выявлены все три основных типа строения перистомов (рис. 2), однако осознание важности их структурных особенностей и преобразований, которые были впоследствии учтены при разработке системы мхов, произошло спустя десятилетия; этот процесс, как мы попытаемся показать, не завершен и по сей день.
Надо сказать, что в 1855 году было закончено издание “Bryologia Europaea” (Bruch et al., 1836–1855) – фундаментальной сводки, иллюстрированной не одним десятком тысяч изображений деталей строения всех видов мхов, встречающихся в Европе, причем почти для всех были даны рисунки перистома анфас и в профиль, в последнем случае на радиальном срезе через устье коробочки. Клетки устья коробочки хотя и были прорисованы, но мелко, так что большинство этих иллюстраций не позволяет уяснить строение перистома, которое, однако, было изучено более тщательно С. Ланциусом-Бенинга (Lantzius-Beninga, 1847, 1850; см. также комментированный перевод Maier, Price, 2014). Сделав продольные срезы перистома десятков видов, он нашел два варианта того, во что продолжается наружная стенка спорового мешка: у Dicranum – в зубец “наружного перистома”, а у Hypnum – в зубец “внутреннего перистома”. Смысл этого наблюдения мы обсудим чуть ниже, при обсуждении работ Филибера.
Другим важным наблюдением стало подразделение типов строения перистомов на артродонтные и нематодонтные, сделанное В. Миттеном (Mitten, 1859), в общем-то, походя, при группировке родов мхов во Флоре востока Индии. Миттен отметил, что виды Polytrichaceae не имеют поперечных перегородок на зубцах, которые есть у всех прочих мхов, что обуславливает их членистый вид.
Следует отметить, что эти наблюдения относятся к додарвиновской эпохе, времени, когда один из ведущих бриологов мира Карл Мюллер рассматривал строение мхов через призму креационизма, объясняя различия видов и родов мхов через различия в соотношении элементов творения (Müller, 1853, цит. по Frahm, 2012). Иными словами, когда ботаники располагали роды растений в “естественные” системы, идея о том, что один тип строения может происходить из другого, еще не была сама собой разумеющейся.
Следующей важной работой, выявившей фундаментальную общность ранних стадий развития перистомов у разных групп видов, стала работа Ф. Киниц-Герлоффа (Kienitz-Gerloff, 1878), которая и по сей день остается одной из наиболее полных и детально иллюстрированных сводок. В ней показаны первые деления спорофита (начиная со стадии в несколько клеток), стадии развития слоев, дающих и перистом, и спорогенную ткань, которые многократно использовались впоследствии в учебниках и обзорных статьях.
Вскоре после этого объемистый труд о строении перистома был опубликован Г. Филибером в серии 18 небольших статей (прямое отношение к данной теме имеют 14) (Philibert, 1884a–1889b). В них рассматривается большинство типов строения перистома мхов и вводятся термины “гаплолепидный перистом” (зубец наружного перистома на дорсальной стороне не разделен срединной линией) и “диплолепидный перистом” (зубец наружного перистома на дорсальной стороне разделен срединной линией). Это важное различение в современной литературе нередко представляется так, что Филиберу мы обязаны пониманием того, что зубцы гаплолепидного перистома гомологичны эндостому, а не экзостому, притом, что первый по своему строению и роли в процессе рассеивания спор гораздо более сходен с экзостомом, чем с эндостомом. На самом же деле Филибер писал, что у дикрановых мхов эндостома (внутреннего перистома) вообще нет, а работы Ланциуса-Бенинга он, возможно, и знал (Maier, Price, 2014), но нигде на них не сослался. Работы Филибера написаны по-французски, судя по всему, таким языком, что не всем было под силу их прочесть. Широкое внимание к ним привлек Е.К. Тэйлор (Taylor, 1962), который изложил их на английском языке. Во введении к своему “переводу” Тэйлор отмечает, что это не вполне обычный перевод, поскольку текст Филибера изобилует отступлениями самого разного толка, мало относящимися к сути дела, так что объем текста “перевода” получился менее трети объема оригинального текста.
Работы Филибера (последняя из которых вышла в 1896 г.), возможно, и не стали бы столь широко известными, если бы выделенные им типы строения перистомов, в первую очередь, разделение мхов с артродонтными перистомами на группы гаплолепидных и диплолепидных (оставим пока за скобками третий тип, гетеролепидные, представленный у Encalyptaceae), не легли в основу классификации мхов М. Флейшера в “Die Musci der Flora von Buitenzorg” (Fleischer, 1900–1923). Несмотря на выглядящее очень “узко локальным” название (Buitenzorg – старое голландское название города на западе острова Ява в Индонезии), именно во введении к первому из четырех томов этой флоры была приведена система мхов, впоследствии взятая, как обычно считается, В.Ф. Бротерусом для обработки мхов в мировом масштабе в Pflanzenreich А.А. Энглера (Brotherus, 1903–1909); во втором издании этой флоры (Brotherus; 1924–1925) представлена система, по которой располагали роды мхов и в гербариях, и в публикациях, на протяжении всего XX века, называя ее системой Флейшера–Бротеруса. Справедливости ради уточним, что оригинальная система Флейшера делила бриевые мхи (т.е. мхи за исключением сфагновых, андреэевых и архидиевых) на 3 группы неопределенного ранга: Arthrodontei (с тремя подгруппами: Haplolepideae, Heterolepideae и Diplolepideae), Amphodontei (буксбаумиевые) и Archodontei (тетрафисовые, политриховые и давсониевые, т.е. нематодонтные в современном понимании). В первом издании сводки Бротеруса строение перистомов не было главным признаком, на котором основывалась система; Бротерус следовал авторитету Шимпера, который в “Corollarium Bryologia Europaea” (Schimper, 1856) считал важнейшим разделение мхов на верхоплодные и бокоплодные (сфагновые Шимпер мхами не считал, андреэевые включал в верхоплодные). При этом Бротерус в первом издании никак не сгруппировал семейства верхоплодных мхов, хотя порядок их расположения более или менее соответствовал тому, который приводил Флейшер (Fleischer, 1900–1923). Во втором издании (Brotherus, 1924–1925), в Pflanzenfamilien Энглера, система мхов включала три подкласса (сфагновые, андреэевые и бриевые), а бриевые группировались в 14 порядков, которые образовывали 3 надпорядка: настоящие бриевые (11 порядков), буксбаумиевые (1) и политриховые (2 порядка, поскольку давсониевые принимались в ранге порядка).
Чтобы понять, насколько систематика тех времен “забывала” или путала важные достижения в изучении строения перистома, можно рассмотреть, как менялось представление о нематодонтном перистоме. Изначально он был определен как перистом, зубцы которого не имеют поперечных перегородок (Mitten, 1859). Филибер сохранил это понимание и отнес Tetraphis к артродонтным мхам, поскольку его зубцы были разделены поперечными перегородками, хотя и отметил его изолированное положение. Флейшер объединил тетрафисовые, политриховые и давсониевые в Archodontei. Бротерус (в своей системе “Флейшера–Бротеруса”) отнес Tetraphis в надпорядок Eubryales. Диксон (Dixon, 1932) включил Tetraphis в нематодонтные, куда он отнес также Buxbaumiales, Diphysciales, Schistostegales и Calomniales (последний порядок уже с середины XX века все относят к печеночникам). Выделяя группу нематодонтных мхов, Диксон обращал внимание на пластинчатую протонему и корневищеобразные структуры, поскольку перистом у Schistostega полностью редуцирован, а у печеночников его и в принципе быть не может. Таким образом, постепенно смысл нематодонтности изменился: к зубцам политрихового типа чаще всего добавляли прочие типы строения, у которых клетки перистомных слоев оставались цельными. Используя это определение, Филибер отмечал, что, в сущности, нематодонтными является часть видов рода Encalypta, особенно E. longicollis, у которой зубцы перистома достигают 5–6 клеток в толщину. Вместе с тем, практически тот же самый факт образования зубцов из целых клеток, отмеченный и Ланциусом-Бенинга, и самим Филибером для Splachnum, почему-то не дал ему повод назвать такой перистом нематодонтным. Ш. Эдвардс (Edwards, 1984) оставил в группе видов с нематодонтным перистомом только Polytrichaceae и Tetraphidaceae, хотя их отличия обсуждаются в тексте столько раз, что вообще непонятно, почему, собственно, Tetraphis отнесен в эту группу. Более того, обзор Эдвардса (Edwards, 1984), который на данный момент в мире является и самым последним, и наиболее полным, нацелен на доказательство того, что перистом политриховых вообще не гомологичен артродонтному перистому, и представление об их гомологии возникло просто из-за того, что их называли одним и тем же словом.
Нет смысла приводить прочие, еще более витиеватые экскурсы о других терминах или, если угодно, “пониманиях” разных типов строения перистома. Отметим лишь здесь тот же аспект, который отмечали и Ш. Эдвардс (Edwards, 1984), и В.Н. Тихомиров44 (Tikhomirov, 1987), когда миграция термина из области морфологии в систематику и обратно “гипнотизирует” и дезориентирует исследователей. Тихомиров проиллюстрировал это на, казалось бы, всем понятном термине цветок, отдельные признаки которого, объявлявшиеся теми или иными авторами ключевыми, вели поиски предка покрытосеменных по ложному следу.
При всей точности отдельных наблюдений исследователей XIX века они не обратили внимания на удивительное постоянство числа клеток в перистомных слоях. Впервые это сделали А. Эванс и Г. Гукер (Evans, Hooker, 1913), показавшие, что на поперечных срезах Ceratodon purpureus три прилегающие к эндотецию слоя сложены, считая снаружи вовнутрь, 48-ю, 24-мя и 32-я клетками соответственно. Этот факт направил Х. Бломквиста и Л. Робертсон к развитию наблюдений в этом направлении, и в итоге, на примере изучения развития Arrhenopterum (Aulacomnium) heterostichum, они сформулировали представление об особо важной роли среднего из трех слоев амфитеция, который они назвали первичным перистомным слоем, ППС, дав также названия и двум другим слоям, НПС и ВПС (Blomquist, Robertson, 1941). Еще через некоторое время Эдвардс (Edwards, 1979) предложил число клеток в перистомных слоях отображать в виде формулы, беря при этом не всю окружность, а лишь ее восьмую часть, так что формулы оказались наглядными и удобными (рис. 2) и вошли в широкое употребление. Две работы Эдвардса (Edwards, 1979, 1984) о строении перистома стали хрестоматийными. Помимо формул перистома, в них подробно обсуждается взаимоотношение групп мхов на основе гомологии отдельных структур. В методическом отношении собственные новые исследования Эдвардса продолжили традицию Ланциуса-Бенинга: они основаны на изучении развитых перистомов, с использованием продольных срезов. Из работ этого цикла следует отметить еще обзор С. Креулена (Kreulen, 1972).
Направление, начатое Киниц-Герлоффом и продолженное в упомянутых работах Эванса и Гукера, а затем Бломквиста и Робертсон, и ряде других работ (Wernderoth, 1931; Saito, Shimoze, 1955; Saito, 1956; Snider, 1975; Stone, 1961; Mueller, 1973), основанное на изучении ранних стадий развития спорофитов, было продолжено в 1980-е годы А. Д. Шоу с соавторами (Shaw, Anderson, 1988; Shaw et al. 1987, 1989a, b). В этих работах были тщательно описаны детали развития перистомов основных типов: гаплолепидных мхов (с формулой перистома 4:2:3) и диплолепидных мхов как с супротивным (с 4:2:4 супр.), так и с очередным типом расположения элементов перистома (с 4:2:4 очер.). Впоследствии работы этого направления продолжились для особо интересных объектов (Schwartz, 1994; Goffinet et al., 1999; Estébanez et al., 2006; Budke et al., 2007; Shimamura, Deguchi, 2008).
Основная проблема, которая возникла в результате работ этого цикла, связана с Diphyscium (Shaw et al., 1987). Выяснилось, что перистом Diphyscium проходит в своем развитии стадию 4:2:3, характерную для гаплолепидных мхов и, как теперь известно, диплолепидных очередных, которые в филогенетическом древе отделены от Diphyscium градой, образованной группами с супротивным расположением элементов экзо- и эндостома. Возможный ответ на этот вопрос обсуждается ниже, где идет речь о строении перистома Catoscopium.
Заканчивая обзор работ XX века, нельзя не отметить две выдающиеся публикации, хотя и не привнесшие новизны в контексте данного обзора. Во-первых, это работа Дейла Мюллера (Mueller, 1973) по развитию перистома с использованием появившихся к тому моменту возможностей трансмиссионной электронной микроскопии. До настоящего времени нет ни одной работы, сравнимой с ней по детальности характеристик всех стадий развития перистома. Вторая работа по развитию перистома Polytrichum Г. Вендерота (Wenderoth, 1931) отличается детальностью описаний первых клеточных делений, создающих крестообразную структуру, которые не описаны никем более столь подробно; небольшая часть их показана на рисунке (рис. 1С).
Кроме того, начиная с 1970-х годов, структура перистомов была замечательным образом изучена с помощью СЭМ и опубликована для сотен видов мхов. Относительная легкость пробоподготовки и фотогеничность изображений сделали их прекрасным иллюстративным материалом, а также хорошим подспорьем для систематики на уровне видов и, иногда, родов. Для выяснения же гомологии и выводов, имеющих далеко идущие последствия, этот метод дал на удивление немного. Например, замечательно представленные в работе Й. Левински (Lewinsky, 1989) на СЭМ фото перистомы ортотриховых мало помогли при попытке решить проблемы их строения и объяснить морфоструктурные преобразования в этой группе. Другой пример – морфоструктурные преобразования перистомов энкалипт, великолепно проиллюстрированные Д. Хортон (Horton, 1982–1983) с помощью СЭМ; следует, однако, отметить, что суть этих преобразований, хотя и не столь наглядно, была описана еще Филибером (Philibert, 1889).
3. СТРОЕНИЕ ПЕРИСТОМА В СВЕТЕ ДАННЫХ О МОЛЕКУЛЯРНОЙ ФИЛОГЕНИИ МХОВ
Второй номер американского журнала “The Bryologist” за 2000 год, безо всякого преувеличения, положил начало новой эры в развитии систематики мхов. В нем была опубликована подборка из 6 статей, на сравнительно больших выборках представившая первые крупные результаты молекулярно-филогенетического анализа мхов, как в целом для всей группы, так и для крупных ее ветвей (Newton et al., 2000; Cox et al., 2000). Хотя последующие работы и внесли важные изменения, общая картина, представленная в этих первых статьях, остается неизменной до настоящего времени и, вероятно, сохранится и в будущем. Опубликованная в 2019 году филогения мхов на основе молекулярно-филогенетического анализа огромного массива данных (Liu et al., 2019), в сущности, только повторила работы 2000 года, уточнив детали, не существенные в контексте данного обзора.
Филогенетическое древо мхов (рис. 3) представленo базальной градой, начинающейся с такакиевых и сфагновых. Они, и следующие за ними андреэевые, андреэобриевые и эдиподиевые не имеют и никогда не имели перистома (Shimamura, Deguchi, 2008). Затем следуют представители групп, перистом которых образован целыми клетками, т.е. нематодонтные мхи (Tetraphidaceae, Polytrichaceae); далее следуют Buxbaumiales, перистом которых имеет сложное строение и обычно считается совмещающим структуры и артродонтного, и нематодонтного перистомов, что обсуждается ниже. Следующим в граде находится порядок Diphysciales, с перистомом, во многом сходным с Buxbaumiales, но обычно считающимся уже полностью артродонтным. Далее начинается участок грады, наиболее сильно разнящийся от одного анализа к другому (в зависимости от набора исследуемых образцов и методов анализа). Он включает 5 порядков: Gigaspermales, Funariales, Disceliales, Encalyptales и Timmiales. Первый из этих 5 порядков не имеет представителей с перистомом, а у остальных перистом (если развит) имеет супротивное расположение элементов экзо- и эндостома. В разных вариантах анализа последние четыре порядка оказываются в базальном положении двух терминальных клад, Bryidae и Dicranidae, или часть порядков группируется друг с другом, однако, в любом случае, эти 5 порядков никогда не образуют клады. При том, что перистомы этих групп сходны супротивным расположением элементов экзо- и эндостома, общее их строение отличается настолько сильно, что в “домолекуляную эпоху” их ставили в три разные части системы мхов, не предполагая между ними сколько-нибудь близкого родства.
Две терминальные клады, включающие большую часть (90%) ныне живущих видов мхов, примерно по 5–6 тысяч видов каждая, имеют перистомы, строение которых сходно между собой настолько, что оно вошло в их названия, особенно в английском языке, в котором краткие неологизмы легко приживаются. Таким образом, в обиходный язык бриологической литературы последнего десятилетия прочно вошли “diplolepideous opposite mosses” (с диполепидным перистомом с супротивным расположением элементов экзостома и эндостома, Funariidae), haplolepideous (c гаплолепидным перистомом, Dicranidae) и “diplolepideous alternate” (с диполепидным перистомом с очередным расположением элементов экзостома и эндостома, Bryidae) (Shaw et al., 2011).
Следует отметить, что, с одной стороны, основные типы строения перистома оказались совпадающими с основными линиями эволюции мхов даже лучше, чем можно было ожидать, а с другой – на уровне родов и видов вариабельность и склонность перистомов к редукции и модификации самого разного толка значительно превзошла ожидания. Особенно это коснулось бокоплодных мхов, систематика которых претерпела кардинальные изменения, поскольку выяснилось, что роды с сильно модифицированными перистомами в очень многих случаях помещались не в те семейства, какие надо (Buck et al., 2000; Huttunen et al., 2004; 2012; Ignatov et al., 2007).
4. НОВЫЙ ВЗГЛЯД НА НЕКОТОРЫЕ ПРОБЛЕМЫ В СВЕТЕ РЕЗУЛЬТАТОВ, ПОЛУЧЕННЫХ ЗА ПОСЛЕДНИЕ ГОДЫ
Наиболее значительные изменения взглядов на строение перистомов в последние годы связаны с изучением аномальных типов строения гаплолепидных перистомов, а также Encalyptales и Buxbaumiales. Вообще говоря, наличие остатков экзостома у гаплолепидных мхов подробно обсуждалось Эдвардсом (Edwards, 1979); им даже был предложен специальный термин – Seligeria-тип перистома, поскольку у некоторых представителей этого рода в основании зубцов гаплолепидного перистома (гомологичного эндостому) остаются небольшие фрагменты редуцированного экзостома, который Эдвардс предложил называть простомом. Особенно подробно Эдвардс (Edwards, 1979) рассмотрел пример Hypodontium dragei (Hypodontiaceae, Dicranales), у которого простом развит так, что зубцы его перистома “диплолепидные” почти до середины. Эти примеры не спровоцировали сколько-нибудь серьезной дискуссии и, возможно, показались не слишком существенными самому Эдвардсу, который в своем хрестоматийном и гораздо более широко известном обзоре строения перистома в “Новом учебнике бриологии “ под редакцией Шустера (Edwards, 1984) не счел нужным упомянуть ни термин простом, ни Seligeria-тип перистома.
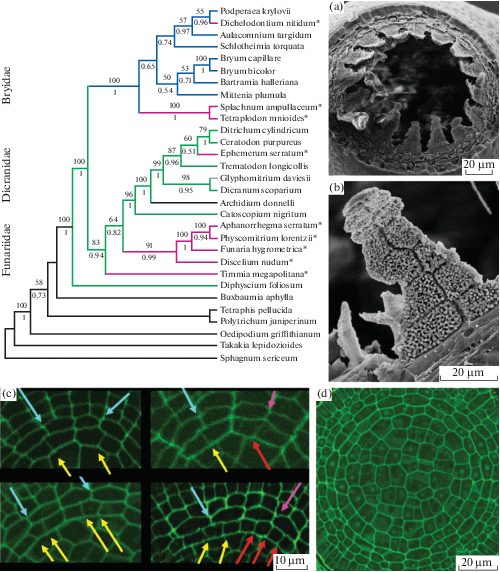
Тем не менее, эти наблюдения Эдвардса оказываются важными в свете того, что впоследствии в Dicranidae (напомним, в англоязычной литературе сейчас это “hapolepideous mosses”) были выявлены представители с еще более развитым экзостомом, чем кто-либо предполагал, а стало быть, в огромной группе основной ее диагностический признак (отсутствие экзостома) оказался не стопроцентно выдержанным. Первым исключением стал монотипный род Catoscopium, исследованный Игнатовым и др. (Ignatov et al., 2015); этот род традиционно относили к меезеевым или к отдельному семейству, близкому к меезеевым и бартрамиевым, то есть Bryidae (“diplolepideous-alternate mosses”). Анализ ранних стадий развития перистома (с применением специального метода выявления характерных паттернов, не очевидных из-за общей сильной редукции) показал наличие у Catoscopium характерной для гаплолепидных мхов формулы перистома 4:2:3, хотя отклонения от нее многочисленны, и на более поздних стадиях распознать ее сложно (рис. 4d). Развернутый филогенетический анализ также подтвердил положение этого рода в основании клады гаплолепидных мхов (рис. 4). Было также выявлено, что при развитии этого частично редуцированного перистома в перистомных слоях происходят дополнительные деления клеток, но, в то же время, “неравные клеточные деления”, обуславливающие образование формулы 4:2:3, оказываются не везде представленными, что приводит к образованию перистомной формулы 4:2:4-супротивного типа (рис. 4с). Авторы высказали предположение, что формула 4:2:4 с супротивным расположением элементов перистома появляется в разных филогенетических линиях, в большинстве случаев просто в связи с редукцией перистома, а не представляет собой отдельного типа строения.
Рис. 4.
Положение в филогенетическом древе мхов с перистомом с супротивным расположением элементов экзостома и эндостома (розовым и отмечено звездочкой), иллюстрирующее возникновения таких групп во всех трех подклассах, а также аномальное положение в “гаплолепидной кладе” Catoscopium с диполепидным перистомом (a–b: СЭМ фото). Поперечный срез молодого спорофита Catoscopium (d) показывает разнообразие расположения клеток в перистомных слоях, а фрагменты этого среза (c) иллюстрируют варианты с образованием формулы 4:2:3 в результате неравных делений в ВПС (желтые и голубые стрелки), или же формулы 4:2:4 в результате нарушения типичных для гаплолепидных мхов неравных делений в ВПС (красные и розовые стрелки) (по Ignatov et al., 2015).
Fig. 4. Phylogenetic tree of mosses showing the independent origin of taxa with opposite exo- and endostome elements (pink color and asterisks), within all three subclasses of Bryopsida, and also anomalous position of Catoscopium in “haplolepideous clade” despite its diplolepideous peristome structure (a–b: SEM). Transverse section of young sporophyte of Catoscopium (d) shows mixture of patterns, partly corresponding to formula 4:2:3, appeared as a result of unequal cell divisions in IPL (c, yellow and blue arrows), or otherwise to formula 4:2:4, formed when unequal divisions in IPL, typical for haplolepideous mosses are not performed (red and pink arrows) (after Ignatov et al., 2015).
Второе исключение оказалось еще более неожиданным и показательным. В. Э. Федосовым с соавторами (Fedosov et al., 2016) с помощью молекулярно-филогенетических методов был впервые изучен род Pseudoditrichum, который также попал в Dicranidae. Если у Catoscopium двойной перистом хоть и имелся, но был отчасти редуцированным, со сросшимися экзо- и эндостомом (рис. 4a, b), то у Pseudoditrichum имеются хорошо развитые, свободные и экзо-, и эндостом (рис. 5). Ранее исследователи не соглашались друг с другом по поводу трактовки устройства этого двойного перистома, которое оказалось довольно необычным – с супротивным расположением элементов наружного и внутреннего круга, но с сегментами эндостома, разделенными на две или три доли. Pseudoditrichum относили к разным подклассам, Funariidae (Steere, Iwatsuki, 1974) или Bryidae (Shaw, 1984). Полученный Федосовым с соавторами результат опроверг обе эти версии. Перистом Pseudoditrichum, как оказалось, представляет ранее не известный тип строения, с формулой 4:2:3, но с хорошо развитым экзостомом и супротивными его зубцам сегментами эндостома, которые очень сходны с простым гаплолепидным перистомом тем, что разделены на две неравные доли. Данное строение оказывается довольно “неудобным” с функциональной точки зрения (эндостом не позволяет экзостому совершать активные движения), что, вероятно, и обуславливает редкость вида (он известен в мире из двух точек). Удивительно то, что Pseudoditrichum в филогенетическом древе располагается далеко от основания “гаплолепидной” клады, все представители которой с хорошо развитым перистомом имеют простой перистом с массивными гигроскопичными зубцами, гомологичными эндостому, в то время как у что Pseudoditrichum сильными и гигроскопичными являются зубцы экзостома, а эндостом становится пленчатым. Онтогенез и факторы такой компенсаторности нуждаются в дальнейшем изучении. Что же касается терминологии, во избежание путаницы или умножения числа типов строения перистомов Федосов с соавторами (Fedosov et al., 2016) предложили считать основным структурным признаком подкласса Dicranidae (“гаплолепидных мхов”) не отсутствие средней линии на зубцах перистома, а соответствие формуле перистома 4:2:3 и разделение зубцов на две неравные доли с чередованием “2:1–1:2”.
Рис. 5.
Перистом Pseudoditrichum mirabile, имеющий уникальное строение (по Fedosov et al., 2015, 2016); a‒b: фото под стереомикроскопом; с–d: СЭМ.
Fig. 5. Peristome of Pseudoditrichum mirabile of unique structure (after Fedosov et al., 2015, 2016); a–b: photos under stereomicroscope; с–d: SEM images.
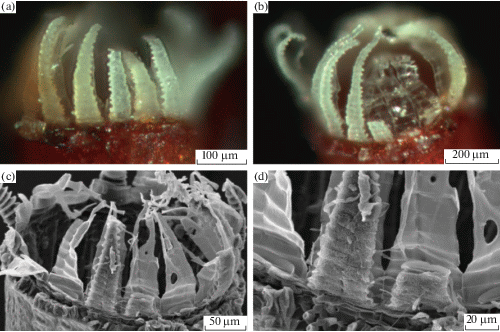
Строение перистомов Encalypta в известной степени продолжает начатую тему, поскольку в этом роде часты случаи, когда эндостом развит значительно лучше экзостома (рис. 6), а экзостом прирастает к нему снаружи в виде “простома” (рис. 6g), который, конечно, проще называть просто экзостомом, не вводя новые сущности (Ignatov et al., 2018с). Разнообразие строения перистома в роде Encalypta столь велико, что неудивительно, что его выделяли в особый, гетеролепидный тип строения. На рис. 6а схематично показаны разные типы строения: Encalypta procera (рис. 6b–d), представляющая наиболее древний подрод (Ignatov et al., 2016), имеет зубцы в двух кругах, сросшиеся в нижней части, будучи, таким образом, вторично нематодонтными; у E. longicolla (рис. 6e) зубцы достигают 5–6 слоев клеток, то есть еще более выраженно “нематодонтные”, что отмечал еще и Филибер; E. brevicolla (рис. 6f) – из двух слоев. Далее вверх по филогенетическому древу у многих видов экзостом развит слабее и прирастает к более сильному эндостому (рис. 6g–h), а у E. ciliata (рис. 6i) перистом образован только эндостомом, материал НПС в процессе формирования перистома утрачивается полностью и, таким образом, перистом полностью удовлетворяет оригинальному определению гаплолепидного типа: наружная его поверхность не разделена срединной линией.
Рис. 6.
Разнообразие строения перистомов Encalypta, показанное на схеме на рис. a и примеры СЭМ фото для Encalypta procera (b–d), E. longicolla (e), E. brevicolla (f), E. rhaptocarpa (g), E. trachymitria (h), E. ciliata (i).
Fig. 6. Diversity of Encalypta peristome structure, schematically shown in a and illustrated by SEM images for Encalypta procera (b–d), E. longicolla (e), E. brevicolla (f), E. rhaptocarpa (g), E. trachymitria (h), E. ciliata (i).
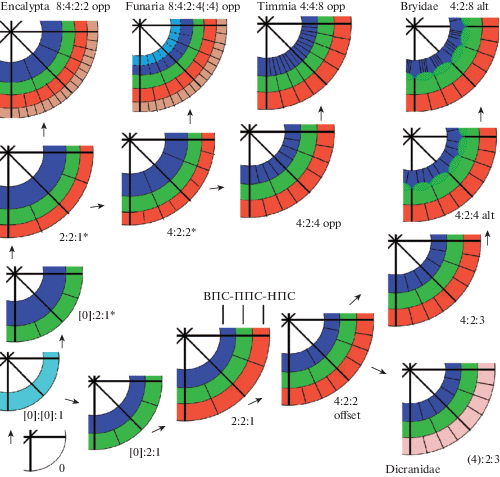
Изучение развития перистомов четырех видов Encalypta (Ignatov et al., 2018c) показало, что все они характеризуются более сильным утолщением клеток внутреннего перистомного слоя (ВПС) (рис. 1A, 7). В процессе развития, как показано на рис. 7, уже на стадии [0]:2:1, у представителей Funariidae заметно более мощное развитие ВПС по сравнению с Bryidae и Dicranidae, что, возможно, и является определяющим и для задержки делений в ВПС на стадии 4:2:2 (в отличие от Bryidae и Dicranidae), и для отсутствия смещения при последующем развитии. Антиклинальные стенки всех перистомных слоев остаются выровненными относительно друг друга, что характерно и для других групп подкласса Funariidae (рис. 7). Более того, в клетках первичного перистомного слоя (ППС) у Encalypta иногда происходят антиклинальные деления, и в таких случаях к одной клетке ВПС будут примыкать две клетки первичного перистомного слоя (ППС). Такие антиклинальные деления в ППС также происходят у Timmia, однако их, по-видимому, совсем нет у Funaria. Выявленные дополнительные деления в ППС ставят вопрос о том, что такое первичный перистомный слой. Критерии, изначально предложенные Бломквистом и Робертсон (Blomquist, Robertson, 1941), звучали следующим образом: “ППС образован 16 клетками, которые уже далее не делятся. Они быстро увеличиваются в размерах и их внутренние стенки становятся выпуклыми. Таким образом, это первый из перистомных слоев, который начинает отличаться от других слоев. Поэтому, а также потому, что количество клеток, его образующих, определяет количество зубцов, представляется целесообразным признать этот слой первичным перистомным слоем”. Полужирным шрифтом я выделил места, по которым ППС Encalypta не удовлетворяет определению этого слоя. Сопоставляя определение Бломквиста и Робертсон с развитием перистома Encalypta, приходится констатировать, что в данном роде ВПС принимает на себя ту роль в регуляции развития перистома, которая обычно принадлежит ППС у большинства мхов с артродонтным перистомом. Эта особенность развития перистома Encalypta может служить объяснением экстраординарного разнообразия перистомов у видов этого рода (рис. 6), которые варьируют от пятислойных до полностью редуцированных, но при редукции всегда редуцируется сначала предперистом, затем экзостом и только потом эндостом, то есть в порядке, зеркальном тому, который свойственен всем артродонтным мхам с двойным перистомом, за редчайшими исключениями.
Рис. 7.
Схема развития перистомов в трех основных эволюционных линиях aртродонтных мхов: Bryidae, Dicranidae и Funariidae (Timmia, Funaria и Encalypta).
Fig. 7. Scheme of peristome development in three main evolutionary lineages of arthrodontous mosses: Bryidae, Dicranidae and Funariidae (Timmia, Funaria, and Encalypta).
Что касается терминологии, не следует что-либо менять в уже устоявшихся названиях трех основных концентрических слоев клеток амфитеция, которые прочно ассоциируются исследователями с их положением: наиболее внутренний ВПС, следующий – ППС и далее НПС. Случаев смещения структуры относительно функции (гетеротопий) у растений множество, так что особая терминология для каждого из них только усложнит понимание сути дела.
В заключение рассмотрим перистом Buxbaumiales, занимающих в филогенетическом древе артродонтных мхов базальное положение (рис. 3). Флейшер, вслед за Филибером, характеризовал их перистом как совмещающий артродонтную часть с нематодонтной. Уникальной конструкции складчатый конус, который авторы единодушно соглашались считать эндостомом, окружен оторочкой из клеток весьма нерегулярного строения, явно целых, и, поскольку выбор того, чем можно было бы их считать, ограничен, не оставалось ничего, кроме того, как назвать эту часть перистома Buxbaumia нематодонтной (рис. 8). Изучение перистомов трех видов буксбаумии (Ignatov et al., 2018d) показало принципиальное отличие развития перистома этого рода от всех остальных мхов: характерное крестообразное расположение клеток в спорофите у Buxbaumia отсутствует. Эндотеций и амфитеций дифференцированы, но в эндотеции клетки располагаются радиально, будучи производными тетраэдрической клетки (рис. 8, 9).
Все клетки внутреннего перистомного слоя (ВПС) располагаются очередно клеткам первичного перистомного слоя (ППС), как это представлено и на поперечных, и на продольных срезах. На ранней стадии развития клетки ВПС оказываются в меньшем количестве (в 1.2–2 раза), чем клетки ППС; затем их число становится одинаковым, а ближе к основанию из-за антиклинальных делений в ВПС клеток в нем становится больше, чем в ППС. В ППС B. aphylla число клеток доходит до 48, однако расчет перистомной формулы путем деления этого числа на 8 в данном случае мало осмыслен. Перистомная формула обычно одна и та же для всего перистома, тогда как здесь отсутствие колонновидной структуры приводит как к значительному увеличению числа клеток в каждом из слоев, так и разному соотношению клеток в перистомных слоях в разных его частях. Строение эндостома Buxbaumia указывает на родство с Diphyscium, что подтверждается и сходным строением части перистома кнаружи от складчатого конуса. Вместе с тем, у Diphyscium развитие перистома на ранних стадиях происходит так же, как и у гаплолепидных мхов, тогда как у Buxbaumia на ранних стадиях развития урночка шире своей длины, и клетки ВПС образуют субэпидермальный слой, покрывающий полушаровидный эндотеций (рис. 9). Такое строение спорофита Buxbaumia вряд ли следует считать примитивным типом, скорее, это высоко специализированное строение: верхняя часть спорофита разрастается и приобретает черты хлоренхимы, что, очевидно, связано с предельной редукцией гаметофита этих безлистных мхов. “Нематодонтную” же часть перистома вряд ли имеет смысл сопоставлять с какими-либо структурами других мхов (за исключением Diphyscium).
Рис. 8.
Buxbaumia aphylla (a–h) и B. minakatae (i). Продольный срез спорофита на ранней стадии развития (a); серия поперечных срезов молодого спорофита, показывает отсутствие крестообразной структуры (b–f на уровнях 2, 32, 46, 58 и 74 мкм от апикальной клетки, на стадии, когда длина перистомных слоев этого спорофита около 150 мкм). Поперечный срез g сделан с более взрослой коробочки, длина перистомных слоев в которой 1300 мкм, и у нее уже б. м. развит складчатый конус (ср. с рис. h и i ).
Fig. 8. Buxbaumia aphylla (a–h) and B. minakatae (i). Longitudinal section of sporophyte at early stage of development (a); transverse section series of young sporophyte, showing lack of cross pattern, characteristic for all other peristomate mosses (sections b–f made at 2, 32, 46, 58 и 74 µm from sporophyte apex, at stage when peristomial layers extend to about 150 µm). Transverse section g shows stage when peristomial layers are as long as 1300 µm and endostome is approaching to pleated cone shape (cf. Figs. h and i ).
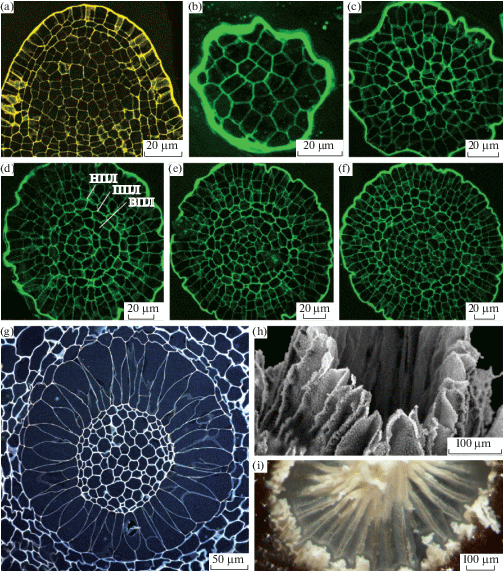
Рис. 9.
Сравнение Buxbaumia с Timmia, представляющей типичный пример мха с артродонтным перистомом. Продольные срезы (d, f) и основанные на них схемы (a, b, c, e) представляющие перистомные слои, правильно выстроенные у Timmia (e, f) и шатровидно расходящиеся у Buxbaumia (Ignatov et al., 2018d). На схемах цветом выделены перистомные слои (НПС, ППС, ВПС и внешний слой эндотеция, ВЭН).
Fig. 9. Comparison of Buxbaumia with Timmia, as a representative of moss with typical arthrodontous peristome. Longitudinal sections (d, f) and schemes based on them (a, b, c, e), showing regular arrangement of peristomial layers in Timmia (e, f) and cone-shaped in Buxbaumia (Ignatov et al., 2018d). Colors denote peristomial layers: НПС – OPL, ППС – PPL, ВПС – IPL, and ВЭН – outer endothecial layer.
ВЫВОДЫ
Прохождение отдельных стадий развития перистома имеет разную степень детерминированности. Накопленные знания и по филогении, и по морфологии мхов позволяют не только отмечать отклонения от типичного варианта развития, но и ранжировать их по сложности морфоструктурных преобразований, а значит и весу, какой они имеют для обсуждения эволюции мхов.
1) Крестообразное расположение клеток в дистальной части спорофита.
Оно свойственно практически всем мхам, кроме буксбаумии, причем формирование такой структуры четко выражено уже у групп, стоящих на эволюционной лестнице заметно ниже уровня, на котором вообще появляется перистом, например у Andreaeobryum (Ignatov et al., 2016). Очевидно, что Buxbaumia – это сильно изолированная боковая ветвь эволюции, и представлять ее как связующее звено между группами с нематодонтным и артродонтным перистомом неправильно (хотя такая точка зрения высказывалась большинством авторов, начиная от Филибера и Флейшера, вплоть до Эдвадрса (Edwards, 1984) и Гоффине и др. (Goffinet et al., 2009).
2) Дифференциация особого слоя клеток в амфитеции.
Клетки амфитеция не отличаются между собой у бесперистомных мхов (Andreaeobryum) и базальных нематодонтных мхов (Tetraphis), увеличиваясь в числе у представителей этих групп от внутренних слоев к наружным. У артродонтных мхов дифференциация есть, кроме случаев предельной редукции, как, например, у Archidium (Snider, 1975). Polytrichaceae обычно рассматривали как группу более базальную по сравнению с Tetraphis (система Бротеруса и, вслед за ней, большинство “Флор…” XX века), хотя в работе Вендерота отмечено, что на поздних стадиях развития спорофита политриховых начинает выделяться меньшим количеством клеток второй, считая изнутри, слой амфитеция (“аналог ППС”). Это указывает на более базальное положение Tetraphis, ближе к Oedipodium, и более продвинутое положение Polytrichaceae, что согласуется и с последними данными молекулярной филогенетики (Liu et al., 2019), и с результатами, полученными при изучении строения плаценты (Ligrone, Ducket, 2011).
3) Определение характера делений ковнутри от ППС.
Анализируя целый ряд примеров супротивного расположения элементов экзо- и эндостома, Игнатов с соавторами (Ignatov et al., 2015) пришли к выводу, что равные деления клеток ВПС у отдельных представителей групп мхов, в которых преобладают неравные деления, свойственны видам с частичной редукцией перистома (рис. 4). Таким образом, возможно, целые группы, в которых утрачена стадия 4:2:3 (= способность к неравным делениям), можно рассматривать как варианты редукции, и перистомы Funariidae – лишь один из таких случаев.
ОСТАЮЩИЕСЯ ПРОБЛЕМЫ И ВОЗМОЖНЫЕ ПУТИ ИХ РЕШЕНИЯ
Как мы видим, в значительной степени изучение перистома, как и изучение цветка, шло двумя параллельными курсами: его исследовали, с одной стороны, в отношении строения и онтогенетического развития на ограниченном количестве объектов, а с другой, систематики, не вникая в строение столь досконально, за счет изучения всех (а не части) видов, и имея в виду также строение не только перистома, но и растений в целом, выстраивали разные типы перистомов в “эволюционные ряды развития”. Разночтения между разными системами оставались значительными до тех пор, пока филогения строилась на основании только морфологических признаков. Возможность доводить до совершенства систематику, используя методы молекулярной филогенетики, закрыла дебаты о положении в системе многих семейств и порядков, хотя среди базальных групп, представленных в современной флоре единичными систематическими реликтами, проблемы, связанные с вымиранием связующих их звеньев, не исчезнут, по-видимому, никогда. Также остается неопределенность роли отдаленной гибридизации, которая у мхов, возможно, более распространена, чем у покрытосеменных (Ignatov et al., 2018b55 ); решение этой проблемы, по-видимому, станет возможным при получении больших объемов данных по последовательностям ДНК. Модусы морфологических преобразований представляют широкое поле для исследований, для которых, впрочем, требуется изучение не единичных срезов, а серий, и исследование объектов, которые ранее не получили достаточного внимания. Учитывая то, что развитие перистомов изучено пока менее чем у 0.5% видов, их изучение, несомненно, позволит выявить еще многие существенные особенности, важные для понимания эволюции мхов.
Список литературы отсутствует.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал