

Ботанический журнал, 2019, T. 104, № 6, стр. 859-887
МЕЗОТРОФНЫЕ КУСТАРНИЧКОВЫЕ СФАГНОВЫЕ И СФАГНОВО-ЗЕЛЕНОМОШНЫЕ ЕЛЬНИКИ ЕВРОПЕЙСКОЙ РОССИИ И УРАЛА
И. Б. Кучеров 1, *, **, С. А. Кутенков 2, ***
1 Ботанический институт им. В.Л. Комарова Российской академии наук
197376 Санкт-Петербург, ул. Проф. Попова, 2, Российская Федерация
2 Институт биологии Карельского научного центра РАН
185910 Республика Карелия, г. Петрозаводск, ул. Пушкинская, 11, Российская Федерация
* E-mail: atragene@mail.ru
** E-mail: IKucherov@binran.ru
*** E-mail: effort@krc.karelia.ru
Поступила в редакцию 18.04.2019
После доработки 01.05.2019
Принята к публикации 04.06.2019
Аннотация
На базе выборки из 321 геоботанического описания, которые выполнены авторами в 1995–2018 гг. или взяты из литературы, проведена доминантно-детерминантная классификация заболоченных мезотрофных ельников (из Picea abies s.l.) сфагновых и сфагново-зеленомошных Европейской России и Урала. В результате выделено 14 синтаксонов разного ранга. Ельники деренно-ерниковые сфагновые растут в северной полосе северотаежной подзоны (крайнесеверной тайге), деренно-черничные и чернично-багульниковые зеленомошно-сфагновые – по всей северной тайге, морошково-черничные сфагновые – в “типичной” северной и в средней тайге. Ельники черничные сфагново-зеленомошные встречаются от северной до южной тайги, черничные и хвощово-черничные сфагновые – в основном в средней и южной тайге. Ельники молиниево-черничные сфагновые отмечены от южной тайги до северной полосы зоны широколиственных лесов, тростниковые сфагновые – однократно отмечены в южной тайге Валдая. В моховом ярусе лесов всех синтаксонов доминирует Sphagnum girgensohnii, часто в сопровождении S. russowii и/или Polytrichum commune.
Настоящая статья продолжает серию публикаций, посвященных заболоченным ельникам (из Picea abies s.l.) Европейской России и Урала. В ней рассматриваются мезотрофные ельники кустарничковые сфагновые и сфагново-зеленомошные, характерной чертой которых является преобладание Sphagnum girgensohnii в моховом ярусе. В отличие от олиготрофных ельников с господством S. angustifolium (Kucherov, Kutenkov, 2019), многим синтаксонам мезотрофных ельников сфагновых свойственны более широкое зональное распространение и наличие замещающих типов в Средней Европе или (реже) в Западной Сибири. У северо- и крайнесеверотаежных синтаксонов мезотрофных ельников кустарничковых сфагновых наблюдаются олиготрофные “синтаксоны-близнецы”, различающиеся преимущественно по видам мохового яруса. О необходимости и методах их разграничения будет сказано в конце статьи.
ИСПОЛЬЗОВАННЫЕ ДАННЫЕ И МЕТОДЫ
Работа основана на выборке из 321 геоботанического описания мезотрофных ельников сфагновых и сфагново-зеленомошных Европейской России и Урала. Из них 269 описаний выполнены авторами в экспедициях 1996–2018 гг. по стандартной методике, подробные ссылки на которую даны в предыдущей статье (Kucherov, Kutenkov, 2019). При описании оценивались проективные покрытия (ПП) видов по ярусам на площади не менее 400 м2, делалась почвенная прикопка или измерялась мощность торфяной залежи.
Как и в предыдущей статье, часть описаний из крайнесеверной и северной тайги Республики Коми (11) взята из рукописи докторской диссертации Ю.П. Юдина (Yudin, 1948), хранящейся в библиотеке БИН РАН. Содержание диссертации опубликовано после смерти ее автора (Yudin, 1954) без публикации исходных данных. Еще 41 описание опубликовано в печати (Regel, 1923; Sambuk, 1932; Salazkin, 1936; Korchagin, 1940, 1956; Katenin, 1972; Kolesnikov, 1985; Morozova, Korotkov, 1999). Классификация растительности проведена доминантно-детерминантным методом, с выделением синтаксонов по доминантам и последующим уточнением их объема по детерминантным группам экологически близких видов (Vasilevich, Bibikova, 2004; Kucherov et al., 2010; Kucherov, Kutenkov, 2019). К одной ассоциации отнесены описания с одинаковым набором доминантов и детерминантов на всем протяжении ее ареала. У субассоциаций могут быть собственные доминанты и детерминанты. Варианты могут выделяться только по детерминантам, модификации – по доминантам (Kucherov, Kutenkov, 2019).
Выделенные при классификации 14 синтаксонов разного ранга сведены в фитоценотическую таблицу (табл. 1), где содержатся также сведения об общем покрытии и высоте ярусов. Названия синтаксонов даны согласно традиции школы В.Н. Сукачева (Sukachev, 1931). Названия в форме, принятой в работах по доминантно-детерминантной классификации (Vasilevich, Bibikova, 2004; Kucherov et al., 2010), приводятся как синонимы. В обоих случаях для краткости наименований использован принцип nomina conservanda, заимствованный из европейских работ (Kucherov, Kutenkov, 2019).
Таблица 1.
Фитоценотическая характеристика мезотрофных ельников кустарничковых сфагновых и сфагново-зеленомошных Европейской России и Урала. Table 1. Phytocoenotic record of mesotrophic dwarfshrub peatmoss and peatmoss-feathermoss spruce forests of European Russia and the Urals
| Вид/Species | Ярус/ Layer | Синтаксоны/Syntaxa | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| Детерминантные виды/Determinants | |||||||||||||||
| Betula nana | b | 9014 | 10014 | 7+ | 7+ | 5+ | |||||||||
| Rubus arcticus | c | 40+ | 26+ | 13+ | 8+ | 3+ | 5+ | 12+ | 15+ | ||||||
| Eriophorum vaginatum | c | 25+ | 531 | 7+ | 7+ | 21+ | 20+ | 3+ | 1 | ||||||
| Bistorta major s.l. | c | 20+ | 211 | 7+ | |||||||||||
| Polytrichum strictum | d | 504 | 633 | 201 | 15+ | 21+ | 23+ | 5+ | 5+ | ||||||
| Cladonia rangiferina s.l. | d | 501 | 32+ | 131 | 401 | 31+ | 7+ | 5+ | 3+ | 4+ | |||||
| Dicranum sp. cf angustum | d | 402 | 421 | ||||||||||||
| D. flexicaule | d | 25+ | 16+ | 8+ | 5+ | 4+ | |||||||||
| Cladonia gracilis subsp. elongata | d | 25+ | 16+ | 8+ | 3+ | ||||||||||
| Betula czerepanovii | a2 + b | 51 | 111 | 7+ | |||||||||||
| Salix myrsinifolia s.l. | a2 + b | 251 | 11+ | 331 | 81 | 5+ | 3+ | 4+ | 3+ | ||||||
| S. glauca | b | 351 | 161 | 272 | 8+ | ||||||||||
| S. lapponum | b | 10+ | 7+ | ||||||||||||
| Chamaepericlymenum suecicum | c | 704 | 633 | 1006 | 7+ | 5+ | 3+ | ||||||||
| Pyrola minor | c | 10+ | 5+ | 13+ | 5+ | 3+ | 4+ | 2+ | |||||||
| Juniperus communis s.l. | b | 553 | 11+ | 672 | 7+ | 152 | 14+ | 18+ | 19+ | 14+ | 15+ | 12+ | 36+ | ||
| Salix phylicifolia | b | 452 | 11+ | 401 | 13+ | 3+ | |||||||||
| Orthilia secunda | c | 601 | 32+ | 731 | 33+ | 381 | 7+ | 25+ | 27+ | 41+ | 19+ | 37+ | 36+ | ||
| Solidago virgaurea s.l. | c | 40+ | 11+ | 731 | 23+ | 3+ | 24+ | 3+ | 271 | 22+ | 18+ | 14+ | |||
| Pedicularis lapponica | c | 10+ | 7+ | ||||||||||||
| Empetrum nigrum s.l. | c | 855 | 844 | 33+ | 9310 | 9210 | 7+ | 331 | 3+ | ||||||
| Vaccinium uliginosum | c | 908 | 893 | 201 | 8712 | 773 | 21+ | 201 | 14+ | 5+ | 2+ | ||||
| Ledum palustre | c | 604 | 582 | 876 | 10026 | 7716 | 14+ | 301 | 11+ | 8+ | 3+ | ||||
| Oxycoccus palustris | c | 25+ | 321 | 8711 | 401 | 381 | 5+ | ||||||||
| Carex vaginata | c | 15+ | 11+ | 6711 | 7+ | 15+ | 2+ | ||||||||
| Nephroma arcticum | d | 401 | 21+ | 27+ | 33+ | 8+ | |||||||||
| Barbilophozia lycopodioides | d | 15+ | 331 | 27+ | 23+ | 11+ | 8+ | 2+ | 9+ | ||||||
| Rubus chamaemorus | c | 905 | 10013 | 604 | 8011 | 464 | 869 | 9516 | 4+ | 18+ | |||||
| Sphagnum russowii | d | 4510 | 6321 | 5312 | 204 | 5411 | 506 | 688 | 274 | 387 | 4+ | 234 | 451 | 14+ | + |
| Gymnocarpium dryopteris | c | 5+ | 5+ | 472 | 7+ | 81 | 291 | 5+ | 16+ | 3+ | 381 | 23+ | |||
| Melampyrum sylvaticum s.l. | c | 10+ | 5+ | 47+ | 211 | 15+ | 24+ | 3+ | 461 | 34+ | 14+ | ||||
| Listera cordata | c | 25+ | 21+ | 47+ | 23+ | 7+ | 10+ | 11+ | 3+ | 8+ | |||||
| Geranium sylvaticum s.l. | c | 5+ | 33+ | 15+ | 3+ | 8+ | 3+ | ||||||||
| Picea abies s.l. | a2 | 403 | 263 | 804 | 674 | 927 | 648 | 437 | 819 | 818 | 9212 | 8310 | 827 | 10013 | |
| Betula pubescens s.l. | a2 | 351 | 212 | 806 | 603 | 856 | 361 | 383 | 433 | 161 | 352 | 653 | 453 | 43+ | |
| Sorbus aucuparia s.l. | a2 + b | 15+ | 932 | 13+ | 382 | 501 | 40+ | 782 | 891 | 922 | 943 | 1003 | 1001 | ||
| Melampyrum pratense s.l. | c | 25+ | 21+ | 601 | 20+ | 691 | 21+ | 35+ | 761 | 651 | 621 | 49+ | 55+ | 431 | |
| Pinus sylvestris | a1 + 2 | 101 | 212 | 271 | 736 | 773 | 576 | 503 | 849 | 817 | 7710 | 666 | 10015 | 10019 | 20 |
| P. sylvestris | b | 10+ | 5+ | 13+ | 471 | 31+ | 14+ | 18+ | 14+ | 14+ | 6+ | ||||
| Cladonia arbuscula s.l. | d | 351 | 16+ | 13+ | 601 | 31+ | 7+ | 8+ | 11+ | 2+ | |||||
| Pinus sibirica | a1 | 81 | 432 | 8+ | 5+ | ||||||||||
| P. sibirica | b | 8+ | 431 | 3+ | 5+ | 2+ | |||||||||
| Maianthemum bifolium | c | 5+ | 7+ | 8+ | 141 | 13+ | 652 | 702 | 10012 | 822 | 821 | 713 | |||
| Luzula pilosa | c | 5+ | 31+ | 29+ | 23+ | 81+ | 57+ | 881 | 63+ | 36+ | 71+ | ||||
| Oxalis acetosella | c | 5+ | 7+ | 8+ | 511 | 321 | 653 | 661 | 181 | 57+ | |||||
| Sorbus aucuparia s.l. | c | 5+ | 7+ | 7+ | 8+ | 7+ | 13+ | 41+ | 54+ | 58+ | 40+ | 36+ | 57+ | ||
| Dicranum polysetum | d | 51 | 13+ | 201 | 311 | 21+ | 381 | 737 | 732 | 812 | 481 | 822 | 1009 | ||
| D. scoparium | d | 15+ | 16+ | 271 | 33+ | 81 | 21+ | 43+ | 594 | 651 | 732 | 521 | 555 | 14+ | |
| Cladonia coniocraea | d + z | 7+ | 13+ | 31+ | 14+ | 28+ | 68+ | 76+ | 69+ | 65+ | 82+ | 71+ | |||
| Plagiothecium laetum | d + z | 5+ | 5+ | 21+ | 18+ | 54+ | 43+ | 57+ | 49+ | 54+ | 100+ | ||||
| Frangula alnus | b | 11+ | 411 | 38+ | 17+ | 733 | 1001 | 10 | |||||||
| Rosa acicularis | b | 301 | 5+ | 7+ | 7+ | 8+ | 3+ | 19+ | 31+ | 35+ | |||||
| Dryopteris expansa s.l. | c | 7+ | 3+ | 19+ | 16+ | 50+ | 32+ | ||||||||
| Anemonoides nemorosa | c | 4+ | 2+ | ||||||||||||
| Dryopteris carthusiana | c | 7+ | 10+ | 24+ | 30+ | 691 | 661 | 27+ | 861 | ||||||
| Salix aurita | b | 51 | 24+ | 6+ | 452 | 3 | |||||||||
| Rubus nessensis | b | 18+ | |||||||||||||
| Betula pendula | a1 | 7+ | 3+ | 243 | 141 | 232 | 8+ | 271 | 572 | ||||||
| Calamagrostis arundinacea | c | 14+ | 5+ | 12+ | 18+ | 27+ | 431 | ||||||||
| Frangula alnus | c | 11+ | 15+ | 9+ | 45+ | 57+ | |||||||||
| Molinia caerulea | c | 5+ | 8+ | 2+ | 1003 | 864 | |||||||||
| Quercus robur | c | 5+ | 36+ | 86+ | |||||||||||
| Q. robur | a1 + 2 | 2+ | 9+ | 29+ | |||||||||||
| Q. robur | b | 5+ | 4+ | 3+ | 9+ | 291 | |||||||||
| Pteridium aquilinum | c | 3+ | 5+ | 15+ | 27+ | 866 | |||||||||
| Carex nigra | c | 3+ | 14+ | 4+ | 6+ | 291 | |||||||||
| C. digitata | c | 3+ | 4+ | 5+ | 29+ | ||||||||||
| Convallaria majalis | c | 3+ | 8+ | 2+ | 291 | ||||||||||
| Phragmites australis | c | 20 | |||||||||||||
| Константные виды / Constants | |||||||||||||||
| Picea abies s.l. | a1 | 10023 | 10028 | 10027 | 10028 | 10030 | 10033 | 9839 | 10044 | 10046 | 9644 | 10046 | 10039 | 10039 | 30 |
| Betula pubescens s.l. | a1 | 656 | 686 | 604 | 737 | 855 | 10011 | 909 | 657 | 594 | 5810 | 788 | 182 | 291 | 10 |
| Picea abies s.l. | b | 552 | 373 | 934 | 874 | 926 | 10013 | 959 | 10016 | 10014 | 10015 | 10015 | 10019 | 10023 | 2 |
| Betula pubescens s.l. | b | 401 | 211 | 1003 | 672 | 771 | 792 | 702 | 461 | 46+ | 27+ | 661 | 55+ | 29+ | |
| Vaccinium myrtillus | c | 908 | 897 | 8721 | 9310 | 10019 | 9313 | 10026 | 10035 | 10027 | 9622 | 10026 | 10030 | 10022 | 3 |
| V. vitis-idaea | c | 1003 | 892 | 935 | 10010 | 10012 | 938 | 1007 | 977 | 1006 | 885 | 987 | 1004 | 862 | 3 |
| Carex globularis | c | 603 | 796 | 404 | 804 | 853 | 10026 | 983 | 462 | 652 | 35+ | 884 | 821 | 712 | 5 |
| Equisetum sylvaticum | c | 703 | 10010 | 733 | 1008 | 461 | 642 | 782 | 652 | 30+ | 651 | 9810 | 9+ | ||
| Pleurozium schreberi | d | 9018 | 795 | 737 | 7311 | 8519 | 936 | 959 | 9724 | 9710 | 10012 | 897 | 1006 | 10028 | + |
| Polytrichum commune | d | 8012 | 8913 | 9311 | 8712 | 928 | 8611 | 835 | 9510 | 848 | 8111 | 9412 | 823 | 861 | 5 |
| Sphagnum girgensohnii | d | 8522 | 7930 | 7326 | 8720 | 10033 | 7943 | 8846 | 8114 | 9563 | 9645 | 8044 | 10074 | 8629 | 95 |
| Hylocomium splendens | d | 757 | 472 | 10020 | 8023 | 9214 | 645 | 857 | 9217 | 653 | 778 | 889 | 452 | 5718 | |
| Прочие виды / Companions | |||||||||||||||
| Populus tremula | a1 + 2 | 5+ | 272 | 13+ | 232 | 71 | 3+ | 3+ | 8+ | 15+ | 11+ | 9+ | 29+ | ||
| Larix sibirica s.l. | a1 + 2 | 5+ | 5+ | 7+ | 231 | 7+ | 10+ | 14+ | |||||||
| Salix caprea s.l. | a2 + b | 15+ | 5+ | 331 | 201 | 312 | 7+ | 22+ | 22+ | 12+ | 25+ | 9+ | 14+ | ||
| Populus tremula | b | 20+ | 20+ | 231 | 3+ | 5+ | 3+ | 12+ | 17+ | 14+ | |||||
| Alnus incana s.l. | a2 + b | 101 | 11+ | 13+ | 13+ | 8+ | 3+ | 18+ | 9+ | ||||||
| Abies sibirica | b | 131 | 8+ | 211 | 14+ | 5+ | 81 | 11+ | |||||||
| Trientalis europaea | c | 451 | 531 | 47+ | 7+ | 15+ | 643 | 33+ | 701 | 73+ | 962 | 952 | 641 | 86+ | 1 |
| Linnaea borealis | c | 601 | 32+ | 933 | 27+ | 463 | 432 | 38+ | 621 | 32+ | 621 | 741 | 27+ | 141 | |
| Avenella flexuosa s.l. | c | 652 | 531 | 734 | 27+ | 691 | 29+ | 23+ | 762 | 701 | 693 | 351 | 73+ | 142 | |
| Lycopodium annotinum s.l. | c | 5+ | 11+ | 40+ | 20+ | 8+ | 431 | 25+ | 38+ | 8+ | 35+ | 34+ | 9+ | 142 | |
| Calamagrostis purpurea s.l. | c | 25+ | 26+ | 20+ | 13+ | 7+ | 13+ | 5+ | 12+ | 22+ | 9+ | ||||
| Carex canescens | c | 101 | 261 | 14+ | 20+ | 5+ | 4+ | 17+ | |||||||
| Rubus saxatilis | c | 5+ | 13+ | 15+ | 3+ | 11+ | 3+ | 19+ | 20+ | 14+ | |||||
| Chamaenerion angustifolium | c | 5+ | 40+ | 13+ | 29+ | 8+ | 3+ | 3+ | 15+ | 9+ | |||||
| Goodyera repens | c | 7+ | 7+ | 5+ | 16+ | 5+ | 12+ | 15+ | 29+ | ||||||
| Chamaedaphne calyculata | c | 5+ | 5+ | 131 | 8+ | 14+ | 33+ | 3+ | 2+ | ||||||
| Aulacomnium palustre | d | 401 | 682 | 27+ | 13+ | 621 | 14+ | 40+ | 27+ | 30+ | 31+ | 37+ | 36+ | 14+ | |
| Ptilium crista-castrensis | d | 5+ | 13+ | 27+ | 46+ | 21+ | 23+ | 411 | 8+ | 191 | 22+ | ||||
| Peltigera aphthosa s.l. | d | 25+ | 21+ | 13+ | 27+ | 23+ | 29+ | 18+ | 11+ | 5+ | 4+ | 11+ | |||
| Sphagnum capillifolium | d | 151 | 75 | 273 | 312 | 144 | 132 | 141 | 5+ | 42 | 233 | 9+ | |||
| Dicranum majus | d | 25+ | 5+ | 13+ | 7+ | 8+ | 18+ | 191 | 5+ | 151 | 14+ | 9+ | |||
| Sphagnum angustifolium | d | 53 | 163 | 73 | 8+ | 296 | 253 | 3+ | 41 | 222 | 9+ | ||||
| S. wulfianum | d | 7+ | 13+ | 73 | 103 | 5+ | 5+ | 191 | 205 | 272 | |||||
| Sciuro-hypnum curtum | d | 8+ | 5+ | 14+ | 11+ | 272 | 14+ | 29+ | |||||||
| Sphagnum magellanicum s.l. | d | 162 | 7+ | 7+ | 301 | 5+ | 8+ | ||||||||
| S. flexuosum | d | 153 | 263 | 7+ | 31+ | 72 | 81 | 8+ | 2+ | ||||||
| Pohlia nutans s.l. | d | 5+ | 7+ | 23+ | 14+ | 18+ | 3+ | 5+ | 2+ | 9+ | 14+ | ||||
| Rhytidiadelphus triquetrus | d | 14+ | 5+ | 5+ | 4+ | 14+ | 29+ | ||||||||
| Sphagnum fallax | d | 211 | 82 | 112 | 4+ | 8+ | |||||||||
| S. riparium | d | 101 | 163 | 7+ | 71 | 81 | 4+ | ||||||||
| S. warnstorfii | d | 5+ | 74 | 133 | 7+ | 5+ | 3+ | 8+ | 5+ | ||||||
| Ptilidium pulcherrimum | d + z | 10+ | 5+ | 13+ | 31+ | 36+ | 25+ | 41+ | 27+ | 23+ | 59+ | 18+ | 57+ | + | |
| Dicranum fuscescens | z | 15+ | 29+ | 43+ | 43+ | 24+ | 31+ | 45+ | 36+ | ||||||
| D. scoparium | z | 5+ | 13+ | 8+ | 7+ | 18+ | 16+ | 46+ | 38+ | 25+ | 18+ | 43+ | |||
| Cladonia deformis | z | 5+ | 11+ | 13+ | 13+ | 31+ | 15+ | 16+ | 22+ | 31+ | 25+ | 18+ | |||
| Dicranum majus | z | 14+ | 13+ | 32+ | 19+ | 31+ | 31+ | 9+ | |||||||
| Pohlia nutans s.l. | z | 5+ | 5+ | 7+ | 8+ | 14+ | 20+ | 24+ | 27+ | 19+ | 20+ | 9+ | 14+ | + | |
| Tetraphis pellucida | z | 14+ | 15+ | 8+ | 24+ | 27+ | 14+ | 27+ | 43+ | + | |||||
| Pleurozium schreberi | z | 5+ | 15+ | 7+ | 8+ | 14+ | 11+ | 15+ | 221 | 9+ | |||||
| Sanionia uncinata | z | 8+ | 19+ | 8+ | 23+ | 14+ | 27+ | 14+ | |||||||
| Средние: сомкнутость крон/проективное покрытие (%) и высота (м) ярусов/ Average: canopy density / cover (%) and height (m) of layers: | |||||||||||||||
| – 1-го яруса древостоя / 1st tree layer | a1 | 0.3/ 11 | 0.4/ 10 | 0.4/ 13 | 0.4/ 11 | 0.4/ 15 | 0.5/ 16 | 0.5/ 19 | 0.6/ 22 | 0.6/ 22 | 0.7/ 23 | 0.6/ 22 | 0.6/ 23 | 0.6/ 24 | 0.6/ ? |
| – 2-го яруса древостоя/2nd tree layer | a2 | <0.1/ 5 | <0.1/ 5 | 0.1/ 5 | 0.1/ 7 | 0.2/ 10 | 0.1/ 9 | 0.1/ 12 | 0.1/ 14 | 0.1/ 15 | 0.2/ 15 | 0.2/ 13 | 0.1/ 15 | 0.2/ 16 | ? |
| – подроста, подлеска / regrowth, shrubs | b | 25/ 1 | 20/ 1 | 15/ 1.5 | 10/ 1.5 | 15/ 2 | 20/ 2 | 10/ 3 | 10/ 2 | 20/ 1.5 | 25/ 2 | 30/ 2 | 25/ 2.5 | 25/ 4 | 15/ ? |
| – травяно-кустарничкового/field layer | c | 55 | 55 | 80 | 85 | 70 | 65 | 60 | 55 | 40 | 55 | 55 | 45 | 45 | 35 |
| – мохового/ground layer | d | 90 | 90 | 95 | 90 | 95 | 90 | 90 | 85 | 90 | 80 | 90 | 95 | 90 | 100 |
| Средний бонитет древостоя / Average stand quality class | V | Va | IV–V | V | IV | IV | III–IV | III | III | III | III | III | II–III | III? | |
| Средняя мощность торфа, см/Average peat thickness, cm | 35 | 40 | 20 | 30 | 25 | 50 | 70 | 15 | 25 | 15 | 25 | 30 | 20 | 30 | |
| Число описаний/Number of relevés (Σ = 321) | 20 | 19 | 15 | 15 | 13 | 14 | 40 | 37 | 37 | 26 | 66 | 11 | 7 | 1 | |
Примечания. Синтаксоны: 1–2 – Piceetum (P.) sphagno girgensohnii-cornoso-nanobetulosum: 1 – var. Juniperus communis, 2 – var. Rubus chamaemorus; 3 – P. sphagno girgensohnii-cornoso-myrtillosum; 4–5 – P. sphagno girgensohnii-ledoso-myrtillosum: 4 – f. Vaccinium uliginosum, 5 – f. typica; 6–7 – P. sphagno girgensohnii-chamaemoroso-myrtillosum: 6 – var. Carex globularis, 7– var. typica; 8–11 – P. sphagno girgensohnii-myrtillosum: 8–9 – subass. typicum: 8 – var. Pleurozium schreberi, 9 – var. typica; 10–11 – subass. equisetosum sylvatici: 10 – f. Maianthemum bifolium, 11 – f. typica; 12–13 – P. sphagno girgensohnii-molinioso-myrtillosum: 12 – subass. typicum, 13 – subass. pleuroziosum schreberi; 14 – P. sphagno girgensohnii-phragmitosum.
Ярусы: a1 и a2 – 1-й и 2-й ярусы древостоя, b – подрост и подлесок, c – травяно-кустарничковый, d – моховой, z – эпифитные и эпиксильные мхи и лишайники. Для видов приводятся постоянство (%) и (в верхнем регистре) среднее проективное покрытие (%). При характеристике древостоя в левой части колонок дана средняя сомкнутость крон, в правой – высота. Детерминантные группы выделены серым фоном; виды в их пределах сгруппированы по ярусам, далее по убыванию встречаемости во всем массиве описаний таблицы. Для доминирующих видов значения покрытия и постоянства даны полужирным шрифтом. Среднее проективное покрытие видов менее 0.5% отмечено плюсом “+”, отсутствие данных – знаком вопроса “?”. Исключены сопутствующие виды со встречаемостью менее 10% хотя бы в одном из синтаксонов (Kucherov et al. 2010; Kucherov, Kutenkov, 2019).
Notes. Syntaxa: 1–2 – Piceetum (P.) sphagno girgensohnii-cornoso-nanobetulosum: 1 – var. Juniperus communis, 2 – var. Rubus chamaemorus; 3 – P. sphagno girgensohnii-cornoso-myrtillosum; 4–5 – P. sphagno girgensohnii-myrtilloso-ledosum: 4 – f. Vaccinium uliginosum, 5 – f. Equisetum sylvaticum; 6–7 – P. sphagno girgensohnii-chamaemoroso-myrtillosum: 6 – var. Carex globularis, 7– var. typica; 8–11 – P. sphagno girgensohnii-myrtillosum: 8–9 – subass. typicum: 8 – var. Pleurozium schreberi, 9 – var. typica; 10–11 – subass. equisetosum sylvatici: 10 – f. Maianthemum bifolium, 11 – f. typica; 12–13 – P. sphagno girgensohnii-molinioso-myrtillosum: 12 – subass. typicum, 13 – subass. pleuroziosum schreberi; 14 – P. sphagno girgensohnii-phragmitosum.
Layers: a1 and a2 – dominant and subordinate tree layers, b – undergrowth and shrubs, c – field layer, d – ground layer, z – epiphytic and epixylic mosses and lichens. Constancy % and (in the upper register) average cover % are given for species. Average canopy density and tree height are given for canopy layers at the left and the right sides of the columns, respectively. The determinant groups are marked by grey backgrounds; species within them are sorted by layer, then by descending occurrence in the whole table relevé set. The cover and constancy values are shown in bold for dominants. Average species cover less than 0.5% is given as “+”, missed data are marked with “?”. Species with occurrence less than 10% at least in one of the syntaxa are excluded (Kucherov et al. 2010; Kucherov, Kutenkov, 2019).
Номенклатура сосудистых растений дана по С.К. Черепанову (Cherepanov, 1995), мхов – по сводке М.С. Игнатова с соавторами (Ignatov et al., 2006), лишайников – по “Списку лихенофлоры России” (A checklist…, 2010). Все европейские таежные популяции ели отнесены к единому комплексу Picea abies s.l. (Popov, 2005). Границы зон и подзон растительности приняты по Т.И. Исаченко и Е.М. Лавренко (Isachenko, Lavrenko, 1980) с последующими уточнениями (Safronova, Yurkovskaya, 2015). В северной части подзоны северной тайги выделяется особая полоса крайнесеверной тайги со специфичным набором синтаксонов (Yudin, 1948, 1954; Bobkova, Patov, 2006). Границы этой полосы, однако, пока не поддаются картированию в масштабах всей Европейской России (Kucherov, Kutenkov, 2019). При картировании синтаксонов (рис. 1–3), кроме данных выборки описаний, учтены сведения из литературных источников.
Рис. 1.
Распространение мезотрофных ельников деренно-ерниковых сфагновых можжевельникового (1) и хвощово-морошкового (2) вариантов, деренно-черничных зеленомошно-сфагновых (3), чернично-багульниковых сфагновых голубичной (4) и типичной (5) модификаций в Европейской России и на Урале (по данным выборки описаний и литературы). Зоны и подзоны: I – тундра, II – лесотундра и подгольцовые редколесья, III – северная, IV – средняя, V – южная тайга. Картооснова (Isachenko, Lavrenko, 1980; Safronova, Yurkovskaya, 2015) объединяет зональные выделы и их высотно-поясные аналоги. Полоса крайнесеверной тайги в северной части подзоны III и (на Северо-Востоке Европейской России) в южной части зоны II не отражена на карте в силу недостатка данных о ее границах (Kucherov, Kutenkov, 2019).
Fig. 1. Distribution of Piceetum (P.) sphagno girgensohnii-cornoso-nanobetulosum var. Juniperus communis (1) and var. Rubus chamaemorus (2), P. sphagno girgensohnii-cornoso-myrtillosum (3), P. sphagno girgensohnii-myrtilloso-ledosum f. Vaccinium uliginosum (4) and f. typica (5) in European Russia and the Urals (mapped according to the available relevé set and the published sources). Zones and subzones: I – tundra, II – forest-tundra and subalpine open woodlands, III – northern-boreal, IV – middle-boreal, V – southern-boreal subzones. The base map (Isachenko, Lavrenko, 1980; Safronova, Yurkovskaya, 2015) is generalized by the association of zonal / subzonal units together with their altitudinal analogs. The northernmost-boreal belt approximately corresponds to the northern part of subzone III and (in the North-East of European Russia) the southern part of zone II; its ranges are known insufficiently (Kucherov, Kutenkov, 2019).
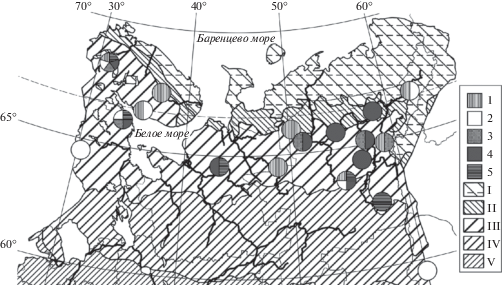
Рис. 2.
Распространение мезотрофных ельников морошково-черничных сфагновых круглоосокового (6) и типичного (7) вариантов, черничных сфагновых зеленомошного (8) и типичного (9) вариантов в Европейской России и на Урале. Зоны и подзоны: VI – подтайга. Прочие обозначения как на рис. 1.
Fig. 2. Distribution of Piceetum (P.) sphagno girgensohnii-chamaemoroso-myrtillosum var. Carex globularis (6) and var. typica (7), and P. sphagno girgensohnii-myrtillosum subass. typicum var. Pleurozium schreberi (8) and var. typica (9) in European Russia and the Urals. Zones and subzones: VI – hemiboreal subzone. For other notes and symbols see Fig. 1.
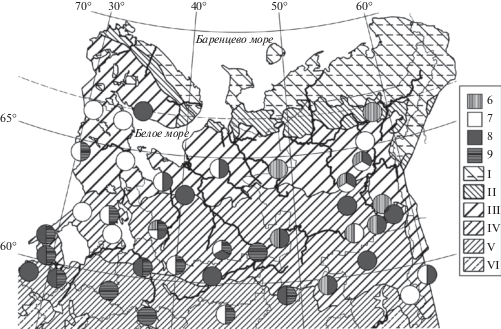
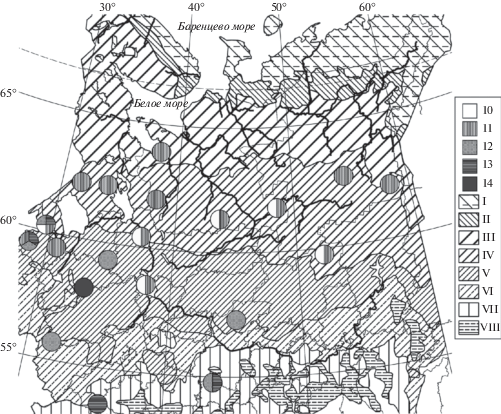
Рис. 3.
Распространение мезотрофных ельников хвощово-черничных сфагновых майниковой (10) и типичной (11) модификаций, молиниево-черничных сфагновых типичной (12) и зеленомошной (13) субассоциаций, тростниковых сфагновых (14) в Европейской России и на Урале. Зоны и подзоны: VII – широколиственные леса, VIII – лесостепь. Прочие обозначения как на рис. 1.
Fig. 3. Distribution of Piceetum (P.) sphagno girgensohnii-myrtillosum subass. equisetosum sylvatici f. Maianthemum bifolium (10) and f. typica (11), P. sphagno girgensohnii-molinioso-myrtillosum subass. typicum (12) and pleuroziosum schreberi (13), and P. sphagno girgensohnii-phragmitosum (14) in European Russia and the Urals. Zones and subzones: VII – broadleaved forests, VIII – forest-steppe. For other notes and symbols see Fig. 1.
Для оценки климатических условий распространения синтаксонов использованы суммы превышений среднесуточных температур над базовой температурой в 10°C (“суммы градусо-дней выше 10°C”; growing degree-days) по данным глобальной сети спутниковой метеосъемки (NASA…, 2006). На основе данных этой же базы рассчитаны индексы континентальности Конрада (Tukhanen, 1980) (табл. 2).
Таблица 2.
Амплитуды среднемноголетних (1983–2004) значений метеопараметров в пределах ареалов синтаксонов мезотрофных ельников кустарничковых сфагновых и сфагново-зеленомошных. Table 2. Amplitudes of averaged (1983–2004) climatic parameter values within the ranges of mesotrophic dwarfshrub-peatmoss / dwarfshrub-peatmoss-feathermoss spruce forest syntaxa
| Синтаксоны/Syntaxa | Метеопараметры/ Climatic parameters | |
|---|---|---|
| GDD > 10, °C | K | |
| 1. Piceetum (P.) sphagno girgensohnii-cornoso-nanobetulosum var. Juniperus communis | 171–410 | 26.6–45.9 |
| 2. Var. Rubus chamaemorus | 171–410 | 26.6–45.9 |
| 3. P. sphagno girgensohnii-cornoso-myrtillosum | 208–360 | 32.2–44.7 |
| 4. P. sphagno girgensohnii-myrtilloso-ledosum f. Vaccinium uliginosum | 208–410 | 32.2–41.9 |
| 5. F. typica | 208–410 | 32.2–44.7 |
| 6. P. sphagno girgensohnii-chamaemoroso-myrtillosum var. Carex globularis | 394–573 | 38.3–44.7 |
| 7. Var. typica | 281–595 | 34.4–43.8 |
| 8. P. sphagno girgensohnii-myrtillosum subass. typicum var. Pleurozium schreberi | 171–686 | 26.6–45.6 |
| 9. Var. typica | 382–686 | 31.2–40.3 |
| 10. Subass. equisetosum sylvatici f. Maianthemum bifolium | 569–658 | 31.2–40.7 |
| 11. F. typica | 471–686 | 31.2–45.6 |
| 12. P. sphagno girgensohnii-molinioso-myrtillosum subass. typicum | 619–1075 | 31.2–42.7 |
| 13. Subass. pleuroziosum schreberi | 619–1075 | 31.2–42.7 |
| 14. P. sphagno girgensohnii-phragmitosum | 727 | 34.9 |
Примечания. GDD > 10 – сумма градусо-дней выше 10°С (NASA, 2006), K – коэффициент континентальности Конрада (Tukhanen, 1980). Амплитуды рассчитаны по данным имеющейся совокупности описаний.
Notes. GDD > 10 – sum of growing degree-days above 10°С (NASA, 2006); K – Conrad continentality index (Tukhanen, 1980). Amplitude calculations are based upon the relevé set in use.
ОБЩАЯ ХАРАКТЕРИСТИКА МЕЗОТРОФНЫХ ЕЛЬНИКОВ КУСТАРНИЧКОВЫХ СФАГНОВЫХ И СФАГНОВО-ЗЕЛЕНОМОШНЫХ
Зональные ареалы рассматриваемых синтаксонов демонстративно различны. Значительная часть последних ограничена в своем распространении северной (в целом) или крайнесеверной тайгой (рис. 1, 2). Именно этим синтаксонам присущи олиготрофные аналоги с господством Sphagnum angustifolium в моховом ярусе (Kucherov, Kutenkov, 2019). Протяженные ареалы других синтаксонов охватывают все подзоны тайги или большую их часть (рис. 2). У этих типов сообществ наблюдаются замещающие их аналоги в горах Средней Европы. Наконец, ряд синтаксонов свойствен только южной тайге и подтайге либо также северной части зоны широколиственных лесов (рис. 3).
Леса “северных” синтаксонов произрастают по долинам рек и ручьев, по окраинам болот в нижней части горно-лесного пояса, в нижней части и при основании склонов увалов, сельг и озовых гряд. По мере продвижения к югу в числе населяемых экотопов возрастает роль плоских низин и ложбин, заболоченных междуречий, окраин равнинных мезотрофных болот и минеральных островов среди них.
Нанорельеф сообществ часто плоский или волнистый, копирующий неровности подстилающей поверхности, при дифференциации на кочки и сфагновые ковры – с низкими кочками высотой 10–20 см. Вдоль ручьев, по окраинам болот, на ветровальных участках дифференциация усиливается. Кочки достигают высоты 50–80 см; появляются вывалы и обводненные западины. Обычно кочки и ковры представлены в равной пропорции, либо доля участков, занятых коврами, незначительно преобладает.
Мезотрофные ельники сфагновые большинства синтаксонов растут на торфяно-подзолисто-глеевых почвах (Smirnova Z., 1928; Yudin, 1954; Medvedeva et al., 1980; Bobkova, 2006a, 2006b) с неглубокой торфяной залежью, что указывает на ускорение биокруговорота сравнительно с олиготрофными ельниками (Kucherov, Kutenkov, 2019). Лишь ельники морошково-черничные приурочены к торфяно-глеевым или торфяно-болотным почвам с более мощной залежью (Sambuk, 1932; Nitsenko, 1960; Bobkova, 2006a). Ельники сфагново-зеленомошные развиты на торфянисто-подзолисто-глееватых почвах (Smirnova Z., 1928; Yudin, 1954; Bobkova, 2006a, 2006b). Кислотность (${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$) торфа в ельниках деренно-ерниковых Терского берега Белого моря составляет 4.3–5.5, возрастая с глубиной отбора образца (Parshevnikov, Chertovskoi, 1961). В ельниках морошково-черничных северной тайги Республики Коми значения pH (3.8–4.3) близки к таковым в олиготрофных лесах, но в сфагново-зеленомошных возрастают до 4.0–4.9, а в черничных сфагновых и сфагново-зеленомошных средней тайги – до 4.1–4.5 (Bobkova, 2006a, 2006b). На карбонатной морене в южной тайге Вологодской обл. ${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ в ельниках черничных сфагновых также не превышает 4.4, но в морошково-черничных достигает 5–6 (Gavrilov, Karpov, 1962). В лесах первого из синтаксонов на кристаллических породах Судет аналогичные цифры равны 4.3–4.8 (Matuszkiewicz, 1977). Значения pHKCl в лесах того же синтаксона в среднетаежной Карелии составляют 4.2–4.7 (Medvedeva et al., 1980).
За вычетом случаев произрастания на торфяно-болотных почвах, корни ели всегда контактируют с минеральными горизонтами профиля. Режим атмосферно-грунтового питания (Kutenkov, Kuznetsov, 2013) способствует увеличению бонитета древостоев до IV и даже III, а сомкнутости их 1-го яруса – до 0.4–0.6. Обычно выражен и 2-й ярус древостоя. В лесах средне- и южнотаежных синтаксонов наблюдается обильный (ПП 15–30%) еловый подрост высотой 2–4 м. Лишь в ерниковых редколесьях крайнесеверной тайги в силу их широтного положения сомкнутость 1-го яруса древостоя не превышает 0.3–0.4, а бонитет – V–Va; 2-й ярус не выражен, а подрост ели малочислен и угнетен (табл. 1), как и в олиготрофных ельниках ерниковых (Kucherov, Kutenkov, 2019).
В кустарничковом ярусе всех синтаксонов, кроме ельников деренно-ерниковых, в число доминантов входит Vaccinium myrtillus. Ее сопровождают V. vitis-idaea и (в зависимости от синтаксона) иные доминанты 1-го или 2-го порядка. Доминант мохового яруса Sphagnum girgensohnii иногда замещается S. russowii, чаще растущим в примеси в сфагновом ковре. Мезогигрофильные виды Sphagnum (S. fallax, S. riparium) редки и тяготеют к западинам; их роль возрастает при прогрессировании заболачивания со сменой ассоциаций. В ельниках сфагново-зеленомошных S. girgensohnii разделяет господство с Polytrichum commune и таежными зелеными мхами (Pleurozium schreberi, Hylocomium splendens, Dicranum polysetum). Polytrichum commune часто растет по ковру Sphagnum girgensohnii, формируя более высокий подъярус. Зеленые мхи тяготеют к микроповышениям, в случае волнистой поверхности почвы – распределены пятнами.
ХАРАКТЕРИСТИКА ВЫДЕЛЕННЫХ СИНТАКСОНОВ
Piceetum (P.) sphagno girgensohnii-cornoso-nanobetulosum (Sphagno girgensohnii-Betulo nanae-P.) – ельник (е.) деренно-ерниковый сфагновый (табл. 1, № 1, 2). Еловые редколесья крайнесеверной тайги, местами проникающие в “типичную” северную тайгу, с характерным ярусом Betula nana высотой до 1 м. Почвы всегда развиты на силикатных породах. Древостой одноярусный, средней высотой 10–11 м, с примесью B. pubescens s.l. к Picea abies s.l. до 2 единиц по составу. Подрост ели малочислен, высотой 1 м. В травяно-кустарничковом ярусе умеренно обильны Vaccinium myrtillus, V. uliginosum, V. vitis-idaea, Chamaepericlymenum suecicum, Empetrum hermaphroditum, Ledum palustre и Carex globularis. В межкочьях растут Eriophorum vaginatum и (единично) мезотрофно-болотные виды (Calamagrostis purpurea s.l., Carex canescens, очень редко Comarum palustre). В моховом ярусе согосподствуют Sphagnum girgensohnii, S. russowii и Polytrichum commune. Детерминанты – гипоарктические (Betula nana, Rubus arcticus, Cladonia gracilis subsp. elongata), гипоарктомонтанные (Dicranum flexicaule) и арктобореальные (Eriophorum vaginatum), в том числе биполярно распространенные (Polytrichum strictum, Cladonia rangiferina s.l.) тундровые и тундрово-болотные виды. К ним примыкает гипоаркто-бореальный болотно-луговой Bistorta major s.l. Другая группа гипоарктических (Chamaepericlymenum suecicum, Betula czerepanovii, Salix borealis) и гипоарктомонтанных (S. glauca, S. lapponum) видов объединяет ельники деренно-ерниковые с деренно-черничными; в кольских сообществах к этой же группе тяготеет Pyrola minor. Выделяются два варианта.
Var. Juniperus communis – можжевельниковый зеленомошно-сфагновый вариант (табл. 1, № 1). Приручейный тип еловых сфагновых редколесий крайнесеверной тайги, на Кольском п-ове обычен также по краю горных болот. Почвы торфянисто- и торфяно-глеевые от песчаных до среднесуглинистых, иногда оподзоленные; мощность торфа 15–60 см. Нанорельеф сглаженный, реже кочковатый; высота кочек 15–40 см. Сомкнутость древостоя 0.3. Подлесок неоднороден по высоте: в верхнем подъярусе – отдельные кусты Juniperus communis s.l., Betula czerepanovii, Salix phylicifolia, S. glauca и S. myrsinifolia s.l., нижний сформирован Betula nana. В травяно-кустарничковом ярусе нет явных доминантов, лишь несколько повышено покрытие Vaccinium uliginosum; Equisetum sylvaticum и Rubus chamaemorus малообильны. В ярусе мхов в число доминантов входит Pleurozium schreberi. Бореальные (Orthilia secunda, Solidago virgaurea s.l.) и гипоаркто-бореальные (Juniperus communis s.l., Salix phylicifolia) лесные и болотно-лесные, в меньшей степени гипоарктические тундровые (Pedicularis lapponica) виды отличают вариант от следующего, одновременно объединяя с ельниками деренно-черничными сфагновыми.
Редколесья данного варианта описаны в крайнесеверной тайге гор Лапландского заповедника, отмечены также среди болот в среднем течении р. Поной (наблюдения С.А. Кутенкова 2018 г.). В литературе имеются описания аналогичных сообществ с Терского берега Белого моря (Betuleto-Piceetum sphagnosum (Regel, 1923)), из междуречья Космы и Тобыша, верховий р. Щугор (P. nano-betuloso-sphagnosum (Yudin, 1948, 1954)) и окрестностей п. Сивая Маска (Katenin, 1972). В “типичную” северную тайгу сообщества проникают в бассейне р. Ижмы (P. sphagnoso-chamaemorosum (Andreyev, 1935)) и на водоразделе Ижмы и Печоры близ п. Нижний Одес (рис. 1).
Var. Rubus chamaemorus – хвощово-морошковый сфагновый вариант (табл. 1, № 2). Сравнительно с предыдущим, сильнее заболочен. Сообщества встречаются не только вдоль ручьев и на речных террасах, но также в нижней трети и при основании склонов увалов; регистрации в горах единичны. Почвы торфяно-глеевые от песчаных до легкосуглинистых; мощность торфа 30–60 см. Нанорельеф бывает как выровненным, так и крупнокочковатым; кочки высотой 30–50 см, занимают половину площади ценоза, между ними стоит вода. Сомкнутость древостоя близка к 0.4, но жизненность его снижена. Верхний подъярус подлеска редуцирован, в нем обильны лишь Salix glauca и Betula czerepanovii. Подъярус B. nana – как в предыдущем варианте. В травяно-кустарничковом ярусе доминируют Rubus chamaemorus и Equisetum sylvaticum; ПП Vaccinium uliginosum снижено. Примесь таежных зеленых мхов к сфагновым невелика. Дифференциация негативная – по отсутствию видов из группы Juniperus communis–Salix phylicifolia.
В крайнесеверной тайге леса этого варианта обычны в районе Сивой Маски (Katenin, 1972), реже встречаются в горах Лапландского заповедника. В “типичной” северной тайге они широко распространены в Беломорье – по Терскому берегу у д. Варзуга (P. sphagnosum (Regel, 1923; Parshevnikov, Chertovskoi, 1961)), в бассейне р. Умбы (P. chamaemoroso-sphagnosum (Salazkin, 1936)) и на Карельском берегу близ п. Чкаловский. Описаны они также в Костомукшском заповеднике (в составе Rubo chamaemori-Piceetum typicum (K.-Lund 1962) Moroz. & Korotk. 1999 (Morozova, Korotkov, 1999)) и на водоразделе Ижмы и Печоры. На юге Северного Урала сходные леса отмечены в горной северной тайге Свердловской обл. по прирусловым участкам речных долин (P. nano-betulosum (Kolesnikov et al., 1973; Rysin, Savelieva, 2002)) (рис. 1). Без деления на варианты мезотрофные ельники ерниковые сфагновые хорошо известны из крайнесеверной тайги бассейна Печоры (P. nanobetuloso-sphagnosum (Sambuk, 1932)).
P. sphagno girgensohnii-cornoso-myrtillosum (Sphagno girgensohnii-Corno suecici-P.) – е. деренно-черничный зеленомошно-сфагновый (табл. 1, № 3). Северотаежные сообщества болотистых речных долин и нижней части склонов гор, реже встречаются на заболоченных водоразделах и вдоль морских берегов. Почвы торфянисто-подзолисто-глеевые или торфяно-глеевые от песчаных до среднесуглинистых, всегда развиты на силикатных породах. Мощность торфа 15–40 см. Иногда торфяная залежь развивается поверх скального водоупора. Слабо выражен двухкомпонентный кочковатый нанорельеф с высотой кочек 10–25, редко 50 см; по площади могут преобладать как кочки, так и ковры. Древостой двухъярусный. Сомкнутость 1-го яруса 0.4 при средней высоте 11 м. Редко высота возрастает до 15–16 м, а бонитет – с V до IV. Фрагментарный 2-й ярус сомкнутостью 0.1, высотой 7 м из Picea abies s.l., Betula pubescens s.l., Sorbus aucuparia s.l. Появление 2-го яруса и возрастание покрытия подроста ели и березы (по 3–5%) указывает на улучшение лесорастительных условий и отличает данный синтаксон (как и все последующие) от ельников деренно-ерниковых. Подлесок редкий из Juniperus communis s.l. и гипоарктических видов Salix, общих с ельниками деренно-ерниковыми var. Juniperus communis. Доминанты травяно-кустарничкового яруса – Vaccinium myrtillus, Carex vaginata и Oxycoccus palustris, ассектаторы – Chamaepericlymenum suecicum, Vaccinium vitis-idaea, Carex globularis, Rubus chamaemorus и Ledum palustre. Полидоминантный моховой ярус сформирован Sphagnum girgensohnii с участием S. russowii, Polytrichum commune и Hylocomium splendens. Детерминанты – мезофильные виды таежно-лесного мелкотравья (Gymnocarpium dryopteris, Melampyrum sylvaticum s.l.) и широкотравья (Geranium sylvaticum s.l.), также Listera cordata. С ельниками деренно-ерниковыми var. Juniperus communis синтаксон объединяют гипоарктические виды из группы Salix glauca–Chamaepericlymenum suecicum. От деренно-ерниковой ассоциации в целом его отличает отсутствие видов из группы Betula nana–Polytrichum strictum.
Ельники этой ассоциации произрастают в обеих полосах северотаежной подзоны. В крайнесеверной тайге они описаны в нижней части лесного пояса гор Лапландского заповедника и в Республике Коми по руч. Седьель и на водоразделе севернее д. Нонбург на р. Цильме. В “типичной” северной тайге сообщества найдены на Карельском берегу Белого моря у п. Чкаловский и в Приполярном Предуралье по р. Большой Аранец (рис. 1). Ельники Коми исходно описаны как P. myrtilloso-sphagnosum (Нонбург) или P. uligini-vaccinioso-sphagnosum (Yudin, 1948, 1954).
P. sphagno girgensohnii-myrtilloso-ledosum (Sphagno girgensohnii-Ledo-P.) – е. чернично-багульниковый зеленомошно-сфагновый (табл. 1, № 4, 5). Сообщества развиты в нижней части “длинных” склонов увалов и сельг, реже на заболоченных водоразделах и минеральных островах среди болот. Почвы торфянисто-подзолисто-глеевые или торфяно-глеевые. В Карелии торфяная залежь часто развивается поверх скальной породы. Нанорельеф кочковатый двух- либо трехкомпонентный, реже выположенный. По площади могут преобладать как кочки высотой 10–50 см, так и ковры. Сомкнутость 1-го яруса древостоя 0.4; примесь Betula pubescens s.l. к Picea abies s.l. до 2, Pinus sylvestris – до 1–2 единиц по составу. Помимо подроста ели и березы высотой 1.5 м есть редкий подрост сосны. Подлесок не выражен. В число доминантов травяно-кустарничкового яруса всегда входят Ledum palustre, Vaccinium myrtillus, V. vitis-idaea и Empetrum nigrum s.l. Carex globularis относится к сопутствующим видам. Моховой ярус тоже полидоминантный с согосподством Sphagnum girgensohnii, Pleurozium schreberi, Hylocomium splendens и Polytrichum commune. Детерминанты – Pinus sylvestris в составе подроста и “спутник” сосны Cladonia arbuscula s.l. От ельников деренно-черничных ассоциацию отличают Pinus sylvestris в древостое, отсутствие гипоарктических видов из группы Salix glauca–Chamaepericlymenum suecicum и таежного мелкотравья из группы Gymnocarpium dryopteris-Melampyrum sylvaticum. От ельников морошково-черничных синтаксон дифференцирует “свита” олиготрофных кустарничков (Empetrum nigrum s.l., Vaccinium uliginosum, Ledum palustre) и сопровождающих их олигомезотрофных видов сосудистых растений (Carex vaginata, Oxycoccus palustris), печеночных мхов (Barbilophozia lycopodioides) и лишайников (Nephroma arcticum).
Ассоциация существует в двух флористически тождественных модификациях.
F. Vaccinium uliginosum – е. чернично-голубично-багульниковый зеленомошно-сфагновый (табл. 1, № 4). Почвы от песчаных до суглинистых, всегда на силикатных породах. Мощность торфа 10–60 см. Нанорельеф всегда кочковатый; по площади могут преобладать и кочки, и ковры. Высота 1-го яруса древостоя 11 м, бонитет V; сомкнутость 2-го яруса 0.1. Покрытие Ledum palustre достигает 30%, Vaccinium myrtillus – лишь 10%. К списку доминантов травяно-кустарничкового яруса добавляются V. uliginosum, Rubus chamaemorus, Equisetum sylvaticum. В ярусе мхов ПП Hylocomium splendens превышает таковое Pleurozium schreberi; Sphagnum russowii относится к сопутствующим видам.
В крайнесеверной тайге Мурманской обл. ельники этой модификации растут в горах Лапландского заповедника, а в Республике Коми – в бассейне р. Усы (P. sphagno-chamaemorosum и P. sphagno-ledosum (Sambuk, 1932)), по рекам Цильме (P. fruticuloso-clado-hypno-sphagnosum) и Чикишне (P. uligini-vaccinioso-sphagnosum (Yudin, 1948, 1954)). В “типичной” северной тайге они известны из Керетской Карелии, со Средней Пинеги (P. uliginosum и P. sphagnosum со Sphagnum russowii и S. warnstorfii (Saburov, 1972)), на Печоро-Ижемском водоразделе, из окрестностей п. Зеленоборск (P. polytrichosum (Bobkova, 2006a)) и д. Кипиево (P. sphagno-ledosum (Sambuk, 1932)) (рис. 1).
Флористически близкие кустарничковые ельники с господством S. compactum развиты на ленточных глинах в средней тайге северо-запада Карельского перешейка (P. fruticuloso-sphagnosum (Dmitrieva, 1973)); их синтаксономический статус пока неясен.
На севере Западной Сибири мезотрофные ельники хвощово-багульниковые сфагновые (P. obovatae ledoso-sphagnosum var. equisetosum) отмечены в верховьях р. Таз (Neshatayev et al., 2002). Ельники с господством Vaccinium uliginosum и Sphagnum girgensohnii (P. uligini-vaccinioso-sphagnosum) также растут по заболоченным долинам ручьев в Приангарье. Однако, судя по сопутствующим видам сосудистых растений (Equisetum pratense, Calamagrostis langsdorffii, Smilacina trifolia, Galium uliginosum (Belov, Ryashin, 1965)), это леса скорее мезоэвтрофных, нежели мезотрофных условий.
F. typica (табл. 1, № 5). Почвы песчаные или супесчаные, могут формироваться и на силикатах, и на карбонатах. Мощность торфа 5–35 см. Нанорельеф выположенный или кочковатый с преобладанием ковров. Высота 1-го яруса древостоя 15 м, бонитет IV; сомкнутость 2-го яруса 0.2. В числе доминантов травяно-кустарничкового яруса остаются лишь Ledum palustre, Vaccinium myrtillus (ПП по 20%) и V. vitis-idaea; прочие виды переходят на роль ассектаторов. В ярусе мхов покрытие Pleurozium schreberi превышает таковое Hylocomium splendens; S. russowii входит в число доминантов.
В крайнесеверной тайге леса этой модификации описаны лишь в горах Лапландского заповедника. В “типичной” северной тайге они известны из Керетской Карелии, с Пинеги (P. ledosum (Saburov, 1972)), Айювы (P. chamaemoroso-sphagnosum (Yudin, 1948, 1954), Печоро-Ижемского водораздела, Средней Печоры (P. sphagno-ledosum (Sambuk, 1932)) и из Илычского Предуралья (P. globulari-caricoso-myrtillosum (Korchagin, 1940)) (рис. 1). Аналогичные ельники тоже отмечены в верховьях р. Таз (P. obovatae ledoso-sphagnosum var. ledosum со Sphagnum girgensohnii (Neshatayev et al., 2002)), равно как и в иных северотаежных районах Западной Сибири (Krylov, 1961).
Замещающий синтаксон в континентальных внутренних районах Норвегии – низкобонитетный ельник с Ledum palustre, Vaccinium uliginosum, Rubus chamaemorus, обильной Carex globularis, также с C. echinata и C. nigra по ковру из Sphagnum girgensohnii, S. flexuosum и S. wulfianum (skog-fattigmyr, granstarr-utforming (Fremstad, 1998)).
P. sphagno girgensohnii-chamaemoroso-myrtillosum (Sphagno girgensohnii-Chamaemoro-P.) – е. морошково-черничный гиргензоново-сфагновый (табл. 1, № 6, 7). Сообщества средней и “типичной” северной тайги, проникающие, однако, как в крайнесеверную, так и в южную тайгу. Произрастают на заболоченных водоразделах, в нижней части пологих склонов, по окраинам болот и ложбинам стока. Сравнительно с лесами прочих обсуждаемых синтаксонов, сильнее всего заболочены. Почвы от песчаных до глинистых. Сомкнутость 1-го яруса древостоя 0.5, 2-го 0.1; примесь Betula pubescens s.l. к Picea abies s.l. до 2, Pinus sylvestris – до 1 единицы по составу. Подрост ели с примесью лишь Betula pubescens, средней высотой 2 м. В подлеске единичные кусты Sorbus gorodkovii и/или S. sibirica. Основные доминанты травяно-кустарничкового яруса – Vaccinium myrtillus и Rubus chamaemorus; их “спутник” – Vaccinium vitis-idaea. Equisetum sylvaticum встречается спорадически, не обилен. Моховой покров с выраженным господством Sphagnum girgensohnii и примесью S. russowii; покрытие Pleurozium schreberi и Hylocomium splendens не превышает 10%. От ельников чернично-багульниковых ассоциацию отличают редкость видов из группы Empetrum nigrum–Vaccinium uliginosum и отсутствие подроста Pinus sylvestris. От ельников черничных сфагновых синтаксон дифференцируют олигомезотрофные тундрово-болотные (Rubus chamaemorus) и болотно-лесные (Sphagnum russowii) гигромезофиты, также отсутствие мезофильного таежного мелкотравья из группы Maianthemum bifolium–Oxalis acetosella. Выделяются два варианта.
Var. Carex globularis – е. круглоосоково-черничный гиргензоново-сфагновый (табл. 1, № 6). Мощность торфяной залежи 30–130 см. Сообщества всегда развиты на силикатных породах. Нанорельеф волнистый, очень редко кочковатый с высотой кочек 30–40 см. Высота древостоя 16 м; бонитет IV. Умеренно (ПП 10%) обилен подрост ели высотой 2 м. Аспектирующий доминант травяно-кустарничкового яруса – Carex globularis (25%); ПП Vaccinium myrtillus составляет 15%, Rubus chamaemorus – 10%. В моховом ярусе Polytrichum commune является доминантом 2-го порядка. Ареал варианта в северной тайге смещен к востоку, его региональным детерминантом cлужит Pinus sibirica.
В крайнесеверной тайге Республики Коми ельники круглоосокового варианта отмечены по Цилемской Печоре и в городском округе Инта (P. sphagno-caricosum (Sambuk, 1932)). В “типичной” северной тайге известны их описания по р. Мезенской Пижме на северо-востоке Архангельской обл. (P. caricoso-sphagnosum (Korchagin, 1956)), а в Коми – на Печоро-Ижемском водоразделе, близ п. Зеленоборск (P. caricoso-sphagnosum (Bobkova, 2006a)) и в Илычском Предуралье (P. globulari-caricoso-sphagnosum с господством Sphagnum russowii (Korchagin, 1940)). Более широко ельники данного варианта распространены в средней тайге. Здесь они произрастают в Южной Карелии (P. globulari-caricosum (Zinserling, 1932)) и на юго-западе Архангельской обл. (Kucherov et al., 2010). Восточнее леса описаны по р. Вычегде от низовий (Урдома) до верховий (P. myrtilli-sphagnosum, P. globulari-caricoso-sphagnosum и P. chamaemori-turfosum (Kolesnikov, 1985)), в Печорском Предуралье и на севере Кировской обл. (Smirnova A., 1954). В южной тайге сообщества отмечены в Ленинградской обл. (Nitsenko, 1960), в том числе в бассейне р. Луги (P. caricoso-sphagnosum (Smirnova Z., 1928)) (рис. 2).
В верховьях р. Таз ельники данного варианта замещаются кедрачами сфагновыми (Pinetum sibiricae sphagnosum girgensohnii) c обилием Rubus chamaemorus, Carex globularis и Ledum palustre при умеренной роли Vaccinium myrtillus (Neshatayev et al., 2002).
Var. typica (табл. 1, № 7). Мощность торфа 20–230 см. Леса могут быть развиты не только на силикатных породах, но и на известняках и гипсах, если изолированы от них залежью глубже 1 м. Нанорельеф с равным соотношением ковров и кочек высотой 20–70 см, реже волнистый. Средняя высота древостоя возрастает до 19 м, бонитет – иногда до III. Еловый подрост гуще (ПП 15%) и выше (3 м), чем в лесах var. Carex globularis. Покрытие Vaccinium myrtillus достигает 25%, Rubus chamaemorus – 15%, Carex globularis – снижается до 3%. Polytrichum commune играет роль ассектатора. Дифференциация негативная по отсутствию Pinus sibirica, в остальном как у ассоциации в целом.
Типичный вариант мезотрофных ельников морошково-черничных обычен и широко распространен, но трудноотличим от аналогичных олиготрофных ельников со Sphagnum angustifolium, особенно в северной тайге, где сообщества обоих типов могут произрастать по соседству, различаясь лишь видами мохового яруса. В крайнесеверной тайге Карелии ельники типичного варианта найдены в национальном парке “Паанаярви”, а в Коми – по Цилемской Печоре (“P. myrtillosum” (Bobkova, Patov, 2006)), Кожве (P. chamaemoroso-sphagnosum (Yudin, 1948, 1954)) и Усе (P. sphagno-chamaemorosum (Sambuk, 1932)). В “типичной” северной тайге Карелии они описаны в Костомукшском заповеднике (Rubo chamaemori-Piceetum typicum (Morozova, Korotkov, 1999)), природном парке “Тулос”, Керетском и Кемском (P. chamaemoro-myrtillosum (Rutkovskii, 1933)) Беломорье, известны и из приграничных районов Финляндии. Восточнее эти леса отмечены вокруг г. Онега, на Средней Пинеге, Среднем Тимане (P. sphagnoso-chamaemorosum (Andreyev, 1935)), Печоро-Ижемском водоразделе, у п. Зеленоборск (P. chamaemoroso-sphagnosum (Bobkova, 2006a)) и по р. Чикишне (Yudin, 1948, 1954).
В средней тайге леса рассматриваемого варианта обычны по всей Южной Финляндии (normale Bruchmoore (Cajander, 1913)) и Карелии. Они найдены также в Кенозерье (Sphagno girgensohnii-P. chamaemoretosum (Kucherov et al., 2010)), близ п. Ротковец (P. sphagnosum (Shimanyuk, 1931)) и в бассейне р. Устьи на юге Архангельской обл., в верховьях р. Вычегды (P. myrtilli-sphagnosum, P. chamaemorosi-turfosum (Kolesnikov, 1985)) и на севере Пермского края (Danilova, 1961). На Урале подобные леса отмечены в горной средней тайге севера Свердловской обл. (Kolesnikov et al., 1973) и при основании склонов хребта Басеги в Пермском крае (P. chamaemoroso-sphagnosum (Balandin, Ladygin, 2002)). В Вологодской и Костромской областях сообщества проникают и в южную тайгу. В первом случае они растут на карбонатной морене водораздела рек Вожега и Сить (Gavrilov, Karpov, 1962), во втором – по болоту в заповеднике “Кологривский лес” (Pismerov et al., 1986) (рис. 2).
Согласно системе болотных местообитаний Финляндии, ельники морошково-черничные сфагновые выделяют как Rubus chamaemorus spruce mires (Eurola et al., 1984); при этом, однако, не различают мезотрофные и олиготрофные сообщества. В Скандинавии ельники морошково-черничные включают в Picea abies-Vaccinium spp.-Sphagnum spp.-typ вместе с черничными сфагновыми (Påhlsson, 1994).
P. sphagno girgensohnii-myrtillosum (Sphagno girgensohnii-Myrtillo-P.) – е. черничный сфагновый (табл. 1, № 8–11). Синтаксономически неоднородная, преимущественно средне- и южнотаежная ассоциация. Сообщества ее различных субассоциаций и вариантов имеют разную зональную и топоэдафическую приуроченность. Нанорельеф волнистый либо двухкомпонентный кочковатый с преобладанием моховых ковров, часто осложнен ветровалом; высота вывалов 0.5–2 м. Сомкнутость 1-го яруса древостоя, как правило, 0.6, второго – 0.1–0.2 при высоте соответственно 22 и 13–15 м; бонитет III. Примесь Pinus sylvestris к Picea abies s.l. – 1–2, Betula pubescens – до 1 единицы по составу древостоя. Средняя высота подроста Picea abies обычно 2 м. В травяно-кустарничковом ярусе всегда господствует Vaccinium myrtillus; в ряде вариантов ей сопутствуют иные доминанты 2-го порядка. Постоянны V. vitis-idaea и (в большинстве вариантов) малообильная Carex globularis; обычен Trientalis europaea. Олиготрофные болотные кустарнички редки и нетипичны. Приствольные повышения нередко без мхов.
От ельников морошково-черничных ассоциацию отличают виды мезофильного таежного мелкотравья (Maianthemum bifolium, Luzula pilosa, Oxalis acetosella), а также бореальные (Dicranum polysetum, Plagiothecium laetum) и бореально-полизональные (Dicranum scoparium, Cladonia coniocraea) лесные мхи и лишайники, кроме того – отсутствие или редкость видов из группы Rubus chamaemorus–Sphagnum russowii.
Выделяются две субассоциации с различным набором доминантов.
Subass. typicum (табл. 1, № 8–9). Единственным доминантом травяно-кустарничкового яруса является Vaccinium myrtillus. Sphagnum russowii эпизодически встречается в примеси к S. girgensohnii. Дифференциация от subass. equisetosum негативная – по отсутствию Dryopteris carthusiana и видов из группы Rosa acicularis–Dryopteris expansa. Выделено два варианта.
Var. Pleurozium schreberi – е. черничный сфагново-зеленомошный (табл. 1, № 8). Широко распространенный вариант, известный как “ельник долгомошный” (Su-kachev, 1931; Sambuk, 1932; Nitsenko, 1960; Rysin, Savelieva, 2002). Его леса представляют собой начальную стадию заболачивания ельников черничных зеленомошных, часто одновременно имея пирогенную природу. Они встречаются от крайнесеверной до южной тайги в понижениях водоразделов и боровых террас и в нижней части пологих склонов, чаще на силикатных песках и супесях, реже на супесях и суглинках поверх известняков и гипсов. Почвы с оторфованной подстилкой толщиной 5–20 см, изредка с маломощной торфяной залежью. Высота кочек 10–30 см. Подрост ели умеренно густой (ПП 10%). Подлесок редкий из единичных кустов Sorbus aucuparia s.l. Ярко выражено господство Vaccinium myrtillus в составе своего яруса (ПП 40%). Equisetum sylvaticum постоянен, но малообилен; обычна Avenella flexuosa. Моховой ярус полидоминантный с преобладанием Pleurozium schreberi и Hylocomium splendens; Sphagnum girgensohnii выступает доминантом 2-го порядка. После пожаров и ветровалов выраженно, но лишь временно возрастает покрытие Polytrichum commune. Дифференциация от var. typica негативная – по отсутствию Frangula alnus и нетипичности Orthilia secunda.
В крайнесеверной тайге ельники этого варианта изучались на Цилемской Печоре (Bobkova, Patov, 2006). В “типичной” северной тайге они описаны по Терскому берегу Белого моря (“P. myrtillosum” (Regel, 1923)) и в Костомукшском заповеднике (в составе Rubo chamaemori-Piceetum pinetosum (K.-Lund 1962) Moroz. & Korotk. 1999 (Morozova, Korotkov, 1999)), далее близ г. Онега, п. Обозерский (P. myrtilloso-polytrichosum (Sokolova, 1937)), на Средней Пинеге (P. empetroso-myrtillosum (Saburov, 1972)), Среднем Тимане (P. polytrichoso-myrtillosum (Andreyev, 1935)), Печоро-Ижемском водоразделе и у п. Зеленоборск (P. polytrichosum (Bobkova, 2006a)). В южной полосе северной тайги Финляндии (Pohjanmaa) сходные ельники известны как Vaccinium-Myrtillus-Typ (Kalela, 1961), а из Финляндии в целом – как gemeine Bruchwälder (Cajander, 1913) и thin-peated spruce heath forests (Eurola et al., 1984). В средней тайге сообщества нередки в Южной Карелии (Zinserling, 1932) и Архангельской обл. от Кенозерья (в составе Sphagno girgensohnii-P. typicum (Kucherov et al., 2010)) до бассейна Устьи и низовий Вычегды. Обычны они и в средней тайге Коми (P. polytrichosum (Bobkova, 2006b)), в том числе в Печорском Предуралье (P. polytricho-myrtillosum (Sambuk, 1932)) и на Верхней Печоре. В Свердловской обл. они отмечены в горной тайге Северного Урала (Kolesnikov et al., 1973).
На границе средней и южной тайги ельники сфагново-зеленомошного варианта покрывают 13% от общей площади еловых лесов на Карельском перешейке (Fedorchuk et al., 2005), описаны на северо-западе последнего (P. myrtilloso-polytrichoso-sphagnosum (Dmitrieva, 1973)) и в заказниках “Линдуловский” и “Озеро Щучье”. На границе подзон сообщества свойственны также северу Вологодской (заказник “Ярбозерский бор”) и Кировской (Smirnova A., 1954) областей и Пермского края (Igoshina, 1930). В южной тайге Ленинградской обл. подобные ельники занимают 11% от площади еловых лесов (Fedorchuk et al., 2005). Они описаны в заказнике “Лисинский” и в бассейне р. Луги (P. polytrichoso-vacciniosum, P. myrtilloso-polytrichosum (Smirnova Z., 1928)), в Вологодской обл. – к югу от р. Кемы и по р. Сухоне (P. sorbosum var. sphagnosum (Korchagin, 1929)), а в Кировской – в Мурашинском районе (Smirnova A., 1954).
Var. typica (табл. 1, № 9). Средне- и южнотаежный вариант, местами проникающий и в северную тайгу, следующая стадия заболачивания ельников черничных. Сообщества встречаются на пониженных участках плоских равнин и по краю сфагновых болот, чаще всего на песках, хотя в Двино-Печорском регионе могут произрастать и на супесях и легких суглинках, в том числе на карбонатной морене. Почвы бедные с оторфованной подстилкой или неглубокой (30–60 см) торфяной залежью. Высота кочек 20–60 см. Подрост ели густой (ПП 20%), но высота его снижена (1.5 м). В подлеске к Sorbus aucuparia s.l. добавляется малообильная Frangula alnus. Покрытие Vaccinium myrtillus снижено до 30%. Equisetum sylvaticum встречается лишь спорадически. В моховом ярусе выраженно господствует Sphagnum girgensohnii (ПП 50%), доминант 2-го порядка на микроповышениях – Pleurozium schreberi. От var. Pleurozium schreberi вариант отличают Frangula alnus и Orthilia secunda.
В северной тайге леса этого варианта лишь однажды отмечены в Костомукшском заповеднике в составе субассоциации Rubo chamaemori-Piceetum pinetosum (Morozova, Korotkov, 1999). Однако в средней тайге они становятся нередкими, будучи известны из заповедника “Кивач” и в целом из Южной Карелии (Kutenkov, Kuznetsov, 2013), а также с юга Архангельской обл., включая Кенозерье (Sphagno girgensohnii-P. typicum (Kucherov et al., 2010)), бассейн Устьи, низовья Вычегды и верховья Северной Двины (Piavchenko, 1957). Часто встречаются они и в средней тайге Республики Коми (P. myrtilloso-sphagnosum (Bobkova, 2006b)). Сообщества обычны также на Северо-Западе России (P. myrtilloso-sphagnosum (Zinserling, 1932; Dmitrieva, 1973); Sphagno girgensohnii-P. (Vasilevich, Bibikova, 2004)), где на них приходится 2% от общей площади еловых лесов на Карельском перешейке, 1% в центре и на юго-западе и 4% на востоке Ленинградской обл. (Fedorchuk et al., 2005). Они описаны в северо-западной части перешейка (Dmitrieva, 1973), на песках в заказниках “Озеро Щучье”, “Комаровский берег”, “Гладышевский” и “Линдуловский”, в центральной части области – в заказнике “Лисинский”, а на юго-востоке – в районе п. Сомино на границе с Вологодской обл. В пределах последней эти леса известны из заказника “Ярбозерский бор” (на песках; средняя тайга) и с водораздела Вожеги и Сити (с господством Sphagnum russowii, на карбонатной морене; южная тайга (Gavrilov, Karpov, 1962)). Восточнее они отмечены также в южной тайге Костромской (заповедник “Кологривский лес” (Pismerov et al., 1986)) и севера Кировской (Smirnova A., 1954) областей. В подтайге ельники данного синтаксона описаны с Молого-Шекснинского междуречья, в дальнейшем затопленного водами Рыбинского водохранилища (P. myrtilloso-sphagnosum (Korchagin, Senyaninova-Korchagina, 1957)).
В финской системе классификации болотных местообитаний ельники черничные сфагновые типичного варианта приводятся как “Vaccinium myrtillus spruce mires” вместе с морошково-черничными (Eurola et al., 1984). В числе доминантов для них указаны и Sphagnum girgensohnii, и S. angustifolium. То же можно сказать и о скандинавском Picea abies-Vaccinium spp.-Sphagnum spp.-typ (Påhlsson, 1994) – сводном типе, объединяющем кустарничковые ельники с доминированием и мезотрофных, и олиготрофных сфагнов.
В Судетах на высоте 1000–1300 м над ур. м. на кристаллических породах описан ельник плауново (Lycopodium annotinum)-бруснично-черничный сфагновый со Sphagnum girgensohnii (Plagiothecio-Piceetum hercynicum sphagnetosum (R.Tx (1932) 1937) Matuszk. 1977). Диагностические виды ассоциации – Calamagrostis villosa, Trientalis europaea, Melampyrum sylvaticum, Galium hercynicum, субассоциации – Listera cordata и Deschampsia cespitosa (Matuszkiewicz, 1977). Сходные леса отмечены и в Высоком Гарце на высоте свыше 600 м над ур. м. (Piceetum excelsum sphagnetosum R.Tx (1932) 1937 (Tüxen, 1937)).
Subass. equisetosum sylvatici – е. хвощово-черничный сфагновый (табл. 1, № 10–11). Субассоциация, переходная к ельникам хвощовым сфагновым; ее сообщества тяготеют к более тяжелым почвам несколько повышенного плодородия. Подрост ели выраженно густой (ПП 30%). Покрытие Vaccinium myrtillus составляет 20–25%, ей сопутствуют доминанты 2-го порядка. Моховой ярус полидоминантный, но Sphagnum girgensohnii остается доминантом 1-го порядка (ПП 40%); Pleurozium schreberi и Polytrichum commune становятся доминантами 2-го порядка. Sphagnum russowii редок. Детерминанты – бореальные (Rosa acicularis, Dryopteris expansa s.l.) и гемибореальные (Anemonoides nemorosa, весенний эфемероид, чье постоянство в массиве описаний таблицы явно занижено) лесные мезофиты. От subass. typicum синтаксон отличает и присутствие Dryopteris carthusiana. Субассоциация представлена двумя модификациями.
F. Maianthemum bifolium – е. майниковый сфагновый (табл. 1, № 10). Леса произрастают в понижениях водоразделов и при основании склонов ледниковых форм рельефа. Почвы от песчаных до легкосуглинистых, иногда щебнистые, всегда на силикатах. Мощность подстилки или торфа не более 30 см. Кочки высотой 10–40 см, их частично имитируют скрытые под подстилкой валуны. Сомкнутость 1-го яруса древостоя возрастает до 0.7, высота – до 23 м, примесь Betula pubescens – до 2 единиц по составу. Подлесок – как в лесах предыдущего варианта. В травяно-кустарничковом ярусе согосподствуют Vaccinium myrtillus и Maianthemum bifolium (ПП 10%), иногда полностью замещающий чернику. Покрытие Oxalis acetosella достигает 3%. Equisetum sylvaticum постоянен, но не обилен; Carex globularis встречается редко. На песчаных и щебнистых почвах с маломощной подстилкой обычна Avenella flexuosa.
Ельники майниковые сфагновые известны из средней тайги Архангельской, Вологодской и севера Кировской (Smirnova A., 1954) областей. В первом из регионов они нередки в бассейне Устьи и в низовьях Вычегды, во втором – отмечены на территории заказника “Ярбозерский бор”. Встречаемость этих лесов возрастает в Северо-Западном регионе России. На них приходится 0.5% от общей площади еловых лесов Карельского перешейка (Fedorchuk et al., 2005), где сообщества описаны в заказниках “Озеро Щучье”, “Комаровский берег”, “Гладышевский” и “Линдуловский”. В южной тайге центра и юго-запада Ленинградской обл. ельники этой модификации занимают 0.5%, а на востоке области – 0.2% от всей площади еловых лесов. Встречаются они также в Новгородской и Псковской областях (Fedorchuk et al., 2005), а в подтайге Ярославской обл. были отмечены на ныне затопленной территории Молого-Шекснинского междуречья (P. vaccinioso-myrtilloso-sphagnosum (Korchagin, Senyaninova-Korchagina, 1957)). В целом, однако, ареал их выявлен не полностью. В северной тайге сообщества не отмечены.
F. typica (табл. 1, № 11). Сообщества заболоченных водоразделов и речных террас, межсельговых понижений, окраин облесенных мезотрофных болот. Почвы чаще от супесчаных до тяжелосуглинистых (на силикатах, иногда на известняках и гипсах), намного реже песчаные. Мощность торфа до 40, редко 60–100 см. Кочки высотой 20‒50 см; иногда встречаются обводненные западины. Характеристики древостоя – как у ассоциации в целом. Постоянство Frangula alnus в подлеске снижено. Доминантом 2-го порядка выступает Equisetum sylvaticum (ПП 10%). Maianthemum bifolium и Oxalis acetosella малообильны; вновь появляется Carex globularis.
В подзоне средней тайги леса этой модификации обычны на юге Архангельской обл. от Кенозерья (в составе Sphagno girgensohnii-P. typicum (Kucherov et al., 2010)) до бассейна Устьи и низовий Вычегды. На северо-западе области они проникают и в северную тайгу близ г. Онега и д. Вонгуда. В средней тайге Карелии подобные ельники описаны в Суоярвском и Кондопожском районах, в Коми – на Южном Тимане (P. equisetoso-sphagnosum (Yudin, 1948, 1954)) и Верхней Печоре, в Вологодской обл. – в заказнике “Ярбозерский бор”, в Кировской – на севере области (Smirnova A., 1954). На Карельском перешейке сходные леса растут на суглинках высокой террасы р. Рощинки в заказнике “Линдуловский”. В южной тайге сообщества отмечены на юго-западе Ленинградской обл. (P. myrtillosum (Smirnova Z., 1928)), а в подтайге – на Молого-Шекснинском междуречье (P. equisetoso-myrtilloso-polytrichosum, P. equisetoso-vaccinioso-myrtilloso-sphagnosum (Korchagin, Senyaninova-Korchagina, 1957)). Возможно, они есть и в Приокско-Террасном заповеднике (наблюдения И.Б. Кучерова 1993 г.).
В горах Средней Европы ельники черничные сфагновые замещает ассоциация Soldanello montanae-Piceetum abietis Volk in Br.-Bl. et al. 1939 (Chytrý et al., 2013), она же Bazzanio-Piceetum Br.-Bl. et Siss. in Br.-Bl. et al. 1939 (Oberdorfer, 1992; Pott. 1995). Это леса прохладного субокеанического климата, развитые на высоте 600–1100 м над ур. м. в Западных Карпатах и Рудных горах (Chytrý et al., 2013), Тюрингском лесу, Шварцвальде и Гарце (Pott, 1995), Баварских (Oberdorfer, 1992) и Австрийских (Willner, Grabherr, 2007) Альпах. Почвы от мелкозалежных подзолисто-глеевых до грубогумусных бурых лесных. Диагностические виды и одновременно доминанты синтаксона – Picea abies, Vaccinium myrtillus, Calamagrostis villosa, Sphagnum girgensohnii, Polytrichum commune, Bazzania trilobata. В древостое встречаются Abies alba и Fagus sylvatica, а в ярусе мхов – Sphagnum quinquefarium, Polytrichum formosum, Rhytidiadelphus loreus вместе с Pleurozium schreberi и Hylocomium splendens (Oberdorfer, 1992; Chytrý et al., 2013). На торфах в субальпийском поясе Швейцарских Альп на высоте 1000–1500 м над ур. м. произрастают ельники флористически близкого синтаксона Sphagno-Piceetum typicum (Richard 1961) Ellenb. & Klötzi 1972. Здесь Vaccinium myrtillus сопутствуют Dryopteris dilatata и малообильный Equisetum sylvaticum, а в ярусе мхов Sphagnum girgensohnii сочетается со S. angustifolium одновременно с Plagiochila asplenioides и Polytrichum formosum (Ellenberg, Klötzi, 1972).
P. sphagno girgensohnii-molinioso-myrtillosum (Sphagno girgensohnii-Molinio-P.) – е. молиниево-черничный сфагновый (табл. 1, № 12–13). Южно- и подтаежная ассоциация, замещающая ельники черничные сфагновые во второй из подзон. Произрастают только на песках, подстилаемых как силикатными породами, так и известняками. Нанорельеф почти всегда кочковатый с выраженным преобладанием ковров, без западин; высота кочек 20–80 см. Сомкнутость 1-го яруса древостоя 0.6, 2-го – 0.1–0.2, высота – соответственно 23–24 и 15–16 м. Примесь Pinus sylvestris к Picea abies – до 2–3 единиц по составу, Betula pubescens – незначительна. Подрост Picea abies достаточно густой (ПП 20%) и высокий. Доминанту травяно-кустарничкового яруса Vaccinium myrtillus сопутствуют Molinia caerulea, малообильные Vaccinium vitis-idaea и Carex globularis. Детерминанты ассоциации – полизональные (Molinia caerulea, Betula pendula) и бореонеморальные (Calamagrostis arundinacea) светолюбивые виды. С ельниками хвощово-черничными синтаксон объединяют Dryopteris carthusiana и виды мезофильного мелкотравья из группы Maianthemum bifolium–Oxalis acetosella. Отличает от названного синтаксона отсутствие таежных видов из группы Rosa acicularis–Dryopteris expansa. Выделены две субассоциации со сходным ареалом.
Subass. typicum (табл. 1, № 12). Сообщества окраин болот и понижений на водоразделах. Почвы с неглубокой залежью мощностью 20–50 см. Бонитет III. Средняя высота подроста 2.5 м. В подлеске Frangula alnus, Sorbus aucuparia и спорадически встречающаяся Salix aurita. В моховом ярусе господствует только Sphagnum girgensohnii. Детерминанты – кустарники подлеска (Salix aurita, Rubus nessensis) и Sphagnum russowii.
Конфигурация ареала субассоциации повторяет таковую Molinia caerulea в южной тайге и подтайге Восточной Европы (Kucherov, 2017). Относящиеся к ней ельники встречаются на границе средней и южной тайги в заказниках “Озеро Щучье” (где очень обычны), “Гладышевский” и “Линдуловский” на юго-западе Карельского перешейка. В южной тайге они отмечены на юго-западе (P. caricoso-sphagnosum с примесью Sphagnum angustifolium к S. girgensohnii (Smirnova Z., 1928)) и юго-востоке Ленинградской обл. В подтайге ельники субассоциации описаны при слиянии рек Межи и Западной Двины и в Нижегородском Приветлужье (Konovalov, Povarnitsyn, 1931; Smirnova A., 1954). Они обычны также в Мордовском заповеднике, где растут в понижениях четвертой (днепровской) террасы р. Мокши на песках, подстилаемых известняками.
В подтайге Северо-Восточной Польши синтаксон описан как Sphagno girgensohnii-Piceetum Polak. 1962. Здесь на торфах поверх песков Vaccinium myrtillus сопровождают Molinia caerulea, Equisetum sylvaticum и таежное мелкотравье, а к Sphagnum girgensohnii примешивается S. palustre. В подлеске обычна Frangula alnus, в древостое – примесь Quercus robur к Picea abies. Диагностические виды – Sphagnum girgensohnii, Listera cordata, Stellaria longifolia, Corallorhiza trifida (Matuszkiewicz, 1977).
Subass. pleuroziosum schreberi – е. молиниево-черничный сфагново-зеленомошный (табл. 1, № 13). Леса этой субассоциации, помимо южной тайги и подтайги, проникают и в северные части зоны широколиственных лесов. Встречаются в нижней части “длинных” склонов и по ложбинам стока. Мощность оторфованной подстилки 5–25 см. Бонитет II–III. Средняя высота подроста 4 м. В подлеске постоянны, но малообильны Frangula alnus и Sorbus aucuparia. К “спутникам” Vaccinium myrtillus добавляются Dryopteris carthusiana и умеренно обильный Pteridium aquilinum. В ярусе мхов согосподствуют Sphagnum girgensohnii, Pleurozium schreberi и Hylocomium splendens. Детерминанты – полизональные (Pteridium aquilinum) и бореонеморальные (Carex digitata, Convallaria majalis) опушечно-лесные мезофиты, также Quercus robur в древостое и в подлеске и Carex nigra.
Ельники этой субассоциации описаны в заказнике “Озеро Щучье” на Карельском перешейке, на склонах Крикковского плато в южной тайге юго-запада Ленинградской обл. (P. myrtilloso-sphagnosum (Smirnova Z., 1928)), в подтайге Мордовского заповедника и в заповеднике “Калужские Засеки” в широколиственно-лесной зоне Калужской обл. Флористически близкие подтаежные ельники II бонитета кустарничково-долгомошные с Molinia caerulea (P. polytrichosum) приводятся из Западно-Двинского и ряда других геоботанических округов Беларуси. Однако для их мохового покрова характерны вкрапления Sphagnum fallax, S. angustifolium и S. magellanicum (Yurkevich et al., 1979).
P. sphagno girgensohnii-phragmitosum (Sphagno girgensohnii-Phragmito-P.) – е. тростниковый сфагновый (табл. 1, № 14). Участок этого синтаксона встречен единственный раз по краю кустарничково-осокового сфагнового болота в южной тайге национального парка “Валдайский” (Новгородская обл.). Торфяная залежь мощностью 30 см поверх песка. Древостой сосново-еловый с примесью Betula pubescens. В подлеске обильна Frangula alnus; подрост ели малочислен. В кустарничково-травяном ярусе доминирует Phragmites australis; на микроповышениях растут Carex globularis, Vaccinium myrtillus и V. vitis-idaea. Единичны Carex echinata, Eriophorum vaginatum. В моховом ярусе к Sphagnum girgensohnii примешиваются Polytrichum commune и (в малом обилии) S. centrale и S. russowii. Указания на аналогичные сообщества в литературе не найдены.
ПРОБЛЕМА РАЗГРАНИЧЕНИЯ “БЛИЗНЕЦОВЫХ” АССОЦИАЦИЙ ОЛИГОТРОФНЫХ И МЕЗОТРОФНЫХ СФАГНОВЫХ ЕЛЬНИКОВ
Почти у каждого синтаксона мезотрофных ельников кустарничковых сфагновых с “центром тяжести” ареала в северной тайге существует “близнецовый” олиготрофный синтаксон со сходным травяно-кустарничковым ярусом, но с моховым покровом из олиготрофных видов сфагновых мхов, прежде всего, Sphagnum angustifolium. Так, с ельниками деренно-ерниковыми (№ 1, 2) сходны олиготрофные ельники ерниковые сфагновые (P. sphagno angustifolii-nanobetulosum), занимающие намного более обширные пространства в полосе крайнесеверной тайги. Нередко приручьевые мезотрофные ельники ерниковые произрастают в окружении олиготрофных. Ельникам чернично-багульниковым зеленомошно-сфагновым голубичной модификации (№ 4) соответствуют багульниковые сфагново-зеленомошные голубичной субассоциации (P. sphagno angustifolii-ledosum subass. uliginosum) и голубичные сфагновые (P. sphagno angustifolii-uliginosum). “Близнецы” ельников чернично-багульниковых типичной модификации (№ 5) – олиготрофные багульниковые типичной субассоциации (P. sphagno angustifolii-ledosum subass. typicum). С ельниками морошково-черничными круглоосокового (№ 6) и типичного (№ 7) вариантов также сходны аналогичные олиготрофные сообщества (соответственно, P. sphagno angustifolii-myrtillosum var. typica f. Carex globularis и f. typica). У ельников чернично-морошково-хвощовых (P. sphagno girgensohnii-chamaemoroso-equisetosum, они же P. sphagno-equisetosum (Sambuk, 1932); P. equisetosum (Rutkovskii, 1933); Equisetum sylvaticum-korpi (Brandt, 1933); Equisetum sylvaticum spruce mires (Eurola et al., 1984); Picea abies-Equisetum sylvaticum-variant (Påhlsson, 1994)) также есть своя олиготрофная пара (P. sphagno angustifolii-myrtillosum var. Equisetum sylvaticum (Kucherov, Kutenkov, 2019)).
Существование столь длинного ряда “близнецовых” пар ельников со Sphagnum angustifolium и S. girgensohnii нельзя объяснить фитоценотической замещаемостью этих видов. Они служат индикаторами совершенно разных режимов минерального богатства и кислотности торфа; замещать же друг друга могут лишь экологически близкие виды сфагнов (Boch, Vasilevich, 1980). Оба названных вида, однако, могут замещаться S. russowii, чья экологическая амплитуда более широка. Ситуация осложняется тем, что в Атлантической и Средней Европе амплитуда толерантности S. girgensohnii также расширяется, и он приобретает способность расти вместе со S. angustifolium (Kielland-Lund, 1981; Oberdorfer, 1992; Dierßen, 1996; Chytrý et al., 2013), что наблюдается и в описанных нами ельниках деренно-морошковых запада Карелии (Kucherov, Kutenkov, 2019). На карбонатных породах случаи совместного произрастания этих видов отмечены и восточнее, в частности, на Среднем Тимане (Andreyev, 1935). К сказанному добавляются возникающие у многих геоботаников трудности при точном распознавании S. angustifolium, S. girgensohnii и зеленоокрашенных форм S. russowii в полевых условиях.
В результате многие авторы при характеристике сфагновых ельников приводят в своих диагнозах как олиготрофные, так и мезотрофные виды сфагновых мхов, благодаря чему появляются описания заведомо гетерогенных типов сообществ (Påhlsson, 1994; Rysin, Savelieva, 2002; Gromtsev, 2008). Менее очевидные примеры из этого же ряда – финские Vaccinium myrtillus spruce mires (Eurola et al., 1984) и Piceetum hylocomioso-sphagnosum из крайнесеверной тайги Усть-Цилемского района Коми (Bobkova, Patov, 2006). В обоих случаях возможно как объединение разнородных сообществ, так и совместное произрастание видов разной экологии.
Предпринимались, однако, и попытки строгого разграничения олиготрофных и мезотрофных сфагновых ельников. Так, в Карелии были раздельно рассмотрены ельники чернично-сфагновые на переходном торфе и багульниково-сфагновые (c господством Ledum palustre, Vaccinium uliginosum и Rubus chamaemorus без участия Equisetum sylvaticum) на двухслойной залежи с верхним слоем верхового и нижним переходного торфа (Medvedeva et al., 1980). Разграничение ельников багульниковых сфагновых Северной Карелии с черничными сфагновыми детально обосновано в работе второго из авторов статьи (Kutenkov, 2015). Есть и примеры раздельного описания олиготрофных и мезотрофных ельников сфагновых, исходя из флористических критериев (Vasilevich, Bibikova, 2004; Kucherov et al., 2010). Однако выделенные в этих работах синтаксоны не являются “близнецовыми”. Возникновение последних часто можно объяснить лишь исходя из забытой ныне теории инкумбации ярусов (Soczava, 1930; Sambuk, 1932). Формирование мезотрофных кустарничковых ельников могло произойти вследствие проникновения синузии болотных кустарничков в черничные и хвощовые сфагновые леса атлантико- и среднеевропейских типов, а олиготрофных – благодаря внедрению в заболоченные мезотрофные ельники (кустарничковые либо травяные) сфагнового ковра верховых болот по мере накопления торфяной залежи (Medvedeva et al., 1980; Kutenkov, 2015).
ЗАКЛЮЧЕНИЕ
Всего в Европейской России и на Урале нами описано 7 ассоциаций мезотрофных ельников кустарничковых сфагновых с четырьмя субассоциациями, шестью вариантами и четырьмя модификациями. Ельникам деренно-ерниковым свойствен “центр тяжести” ареала в крайнесеверной тайге, деренно-черничным и багульниково-черничным – в северной тайге в целом. Ельники морошково-черничные равно характерны для “типичной” северной и для средней тайги, черничные сфагново-зеленомошные – для всех подзон от северной до южной тайги, прочие три синтаксона ельников черничных сфагновых – лишь для средней и южной тайги. Ельники тростниковые сфагновые отмечены лишь в южной тайге, а у молиниево-черничных “центр тяжести” ареалов обеих субассоциаций находится в южной тайге и подтайге. Амплитуды толерантности сообществ всех синтаксонов по отношению к фактору теплообеспеченности вегетационного периода строго соответствуют их зональной приуроченности (табл. 2).
Большая часть синтаксонов равномерно представлена во всех долготных секторах Европейской России. Лишь для ельников морошково-черничных круглоосокового варианта выявлено смещение их ареала на восток в северной тайге, а для молиниево-черничных – напротив, тяготение к западу в северной (южнотаежной) части их ареала. Ни у одного из синтаксонов в пределах изучаемой территории географическое распространение не соответствует изменениям континентальности климата (табл. 2).
Ельники молиниево-черничные, чернично-багульниковые типичной модификации, черничные и хвощово-черничные типичных вариантов могут расти и на силикатных, и на карбонатных породах, морошково-черничные типичного варианта и черничные сфагново-зеленомошные – также на сульфатных. Однако для сообществ всех названных синтаксонов, кроме последнего, необходимо перекрытие этих пород достаточно мощной торфяной залежью или плащом силикатных четвертичных отложений. Леса всех прочих синтаксонов строго приурочены к силикатным породам.
ДОПОЛНЕНИЕ. НЕУЧТЕННЫЕ НАХОДКИ ОЛИГОТРОФНЫХ ЕЛЬНИКОВ СФАГНОВЫХ
При подготовке статьи были выявлены находки ряда синтаксонов олиготрофных ельников сфагновых, расширяющие представления об их ареалах, но неучтенные в предыдущей публикации (Kucherov, Kutenkov, 2019). Так, в средней тайге Пряжинского района Карелии у п. Колатсельга найдены ельники деренно-морошковые сфагновые (P. sphagno angustifolii-cornoso-chamaemorusum), ранее известные лишь в северной тайге Карелии. Их отличают высокое постоянство (V против III в северной тайге) и возросшее покрытие Molinia caerulea. Вырубки на месте подобных лесов с обильным разрастанием M. caerulea, но без Chamaepericlymenum suecicum описаны в бывшем природном парке “Хиисъярви” (средняя тайга Суоярвского района Карелии) как Molinia-korpi (Brandt, 1933).
Неучтенные описания олиготрофных ельников морошково-черничных (P. sphagno angustifolii-myrtillosum var. typica f. typica) были выполнены в южной тайге Ленинградской (заказник “Лисинский”) и Новгородской (национальный парк “Валдайский”) областей, на песчаных почвах. На то, что ельники данного синтаксона встречаются и в южной тайге, не было указано ранее (Kucherov, Kutenkov, 2019).
Список литературы
[A checklist…] Список лихенофлоры России. 2010. СПб. 194 с.
[Andreyev] Андреев В.Н. 1935. Лесная растительность южного Тимана. – Тр. Полярной комиссии СОПС АН СССР. 24: 7–64.
[Balandin, Ladygin] Баландин С.В., Ладыгин И.В. 2002. Флора и растительность хребта Басеги (Средний Урал). Пермь. 191 с.
[Belov, Ryashin] Белов А.Г., Ряшин В.А. 1965. Растительный покров левобережной части Нижнего Приангарья. – В кн.: Растительный покров Красноярского края. Новосибирск. С. 165–178.
[Bobkova] Бобкова К.С. 2006a. Еловые леса северной подзоны тайги. – В кн.: Коренные еловые леса Севера: Биоразнообразие, структура, функции. СПб. С. 52–98.
[Bobkova] Бобкова К.С. 2006b. Еловые леса средней подзоны тайги. – В кн.: Коренные еловые леса Севера: Биоразнообразие, структура, функции. СПб. С. 99–158.
[Bobkova, Patov] Бобкова К.C., Патов А.И. 2006. Еловые леса крайнесеверной подзоны тайги (притундровой зоны). В кн.: Коренные еловые леса Севера: Биоразнообразие, структура, функции. СПб. С. 25–33.
[Boch, Vasilevich] Боч М.С., Василевич В.И. 1980. Состав и структура растительности грядово-мочажинного комплекса. – Экология. 3: 22–30.
Brandt A. 1933. Hiisjarven luonnonpuiston kasvillisuudesta. – Silva Fenn. 32: 1–108.
Cajander A.K. 1913. Studien über die Moore Finnlands. – Acta Forest. Fenn. 2 (3): 1–208.
[Cherepanov] Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб. 991 с.
Chytrý M., Zelený D., Návratilová J., Sádlo J. 2013. Boreokontinentální jehličnaté lesy. – In: Vegetace České republiky. T. 4: Lesní a křovinná vegetace. Praha. S. 380–432.
[Danilova] Данилова М.М. 1961. Еловые леса средней тайги Пермской области. – Учен. записки Пермского гос. ун-та. 18 (3): 11–19.
Dierßen K. 1996. Vegetation Nordeuropas. Stuttgart. 839 S.
[Dmitrieva] Дмитриева Е. В. 1973. Ельники северо-западной части Карельского перешейка. – Лесоведение. 2: 51–64.
Ellenberg H., Klötzi F. Waldgesellschaften und Waldstandorte der Schweiz. – Mitt. schweiz. Anst. forst. Versuchswesen. 48 (4): 587–930.
Eurola S., Hicks S., Kaakinen E. 1984. Key to Finnish mire types. – In: European Mires. L. 117 p.
[Fedorchuk et al.] Федорчук В.Н., Нешатаев В.Ю., Кузнецова М.Л. 2005. Лесные экосистемы северо-западных районов России: Типология, динамика, хозяйственные особенности. СПб. 382 с.
Fremstad E. 1998. Vegetasjonstyper i Norge. 2. oppl. – NINA Temahefte 12: 1–279.
[Gavrilov, Karpov] Гаврилов К.А., Карпов В.Г. 1962. Главнейшие типы леса и почвы Вологодской области в районе распространения карбонатной морены. – Тр. ин-та леса и древесины. 52: 5–119.
[Gromtsev] Громцев А.Н. 2008. Основы ландшафтной экологии европейских таежных лесов России. Петрозаводск. 238 с.
Ignatov M.S., Afonina O.M., Ignatova E.A. 2006. Check-list of mosses of East Europe and North Asia. – Arctoa: Бриологический журнал. 15: 1–130.
[Igoshina] Игошина К.Н. 1930. Растительность северной части Верхне-Камского округа Уральской области. – Тр. биол. ин-та Перм. ун-та. 3 (2): 73–176.
[Isachenko, Lavrenko] Исаченко Т.И., Лавренко Е.М. 1980. Ботанико-географическое районирование. – В кн.: Растительность европейской части СССР. Л. С. 10–22.
Kalela A. 1961. Waldvegetationszonen Finnlands und ihre klimatischen Paralleltypen. – Arch. Soc. Zool. Bot. Fenn. “Vanamo”. 16 (suppl.): 65–83.
[Katenin] Катенин А.Е. 1972. Растительность лесотундрового стационара. – В кн.: Почвы и растительность восточноевропейской лесотундры. Л. С. 118–259.
Kielland-Lund J. 1981. Die Waldgesellschaften SO Norwegens. – Phytocoenologia. 9 (1–2): 53–250.
[Kolesnikov] Колесников Б.П. 1985. Лесная растительность юго-восточной части бассейна Вычегды. Л. 216 с.
[Kolesnikov et al.] Колесников Б.П., Зубарева Р.С., Смологонов Е.П. 1973. Лесорастительные условия и типы леса Свердловской области. Свердловск. 176 с.
[Konovalov, Povarnitsyn] Коновалов Н.А., Поварницын В.А. 1931. Лесные ассоциации Баковского лесничества Бакопытлесхоза Нижегородского края. – В кн.: Природа и хозяйство учебно-опытных лесничеств Ленинградского Лесного института. М. Вып. 2. С. 252–303.
[Korchagin] Корчагин А.А. 1929. К вопросу о типах леса по исследованию в Тотемском уезде Вологодской губернии. – В кн.: Очерки по фитосоциологии и фитогеографии. М. С. 287–327.
[Korchagin] Корчагин А.А. 1940. Растительность северной половины Печорско-Ылычского заповедника. – В кн.: Труды Печорско-Ылычского заповедника. Т. 2. С. 5–415.
[Korchagin] Корчагин А.А. 1956. Еловые леса западного Притиманья в бассейне р. Мезенской Пижмы. – Учен. записки Ленингр. гос. ун-та. Сер. Геогр. 11: 111–239.
[Korchagin, Senyaninova-Korchagina] Корчагин А.А., Сенянинова-Корчагина М.В. 1957. Леса Молого-Шекснинского междуречья (дубняки, черноольшанники и ельники). – Тр. Дарвинского гос. заповедника. 4: 291–402.
[Krylov] Крылов Г.В. 1961. Леса Западной Сибири. М. 255 с.
[Kucherov] Кучеров И.Б. 2017. Изменчивость эколого-ценотической приуроченности Molinia caerulea (Poaceae). – Бот. журн. 102 (11): 1475–1503. https://doi.org/10.1134/S0006813617110011
[Kucherov et al.] Кучеров И.Б., Разумовская А.В., Чуракова Е.Ю. 2010. Еловые леса национального парка “Кенозерский” (Архангельская область). – Бот. журн. 95 (9): 1268–1301.
[Kucherov, Kutenkov] Кучеров И.Б., Кутенков С.А. 2019. Олиготрофные сфагновые и сфагново-зеленомошные ельники Европейской России и Урала. – Бот. журн. 104 (1): 12–40. https://doi.org/10.1134/S000681361901006X
[Kutenkov] Кутенков С.А. 2015. Анализ растительности ельников чернично-сфагновых северотаежной подзоны Восточной Фенноскандии. – Тр. Гос. природн. заповедника “Костомукшский”. 1: 61–68.
[Kutenkov, Kuznetsov] Кутенков С.А., Кузнецов О.Л. 2013. Разнообразие и динамика заболоченных и болотных лесов Европейского Севера России. – В кн.: Разнообразие и динамика лесных экосистем России. Кн. 2. М. С. 152–206.
Matuszkiewicz J. 1977. Przegląd fitosocjologiczny zbiorowisk leśnych Polski. Cz. 4. Bory świerkowe i jodłowe. – Phytocoenosis. 6 (3): 151–226.
[Medvedeva et al.] Медведева В.М., Корнилова Л.И., Вайнблат В.З. 1980. Основные типы заболоченных ельников Карелии. – В кн.: Болотно-лесные системы Карелии и их динамика. Л. С. 78–99.
[Morozova, Korotkov] Морозова О.В., Коротков В.Н. 1999. Классификация лесной растительности Костомукшского заповедника. – Заповедное дело. 5: 56–78.
NASA surface meteorology and solar energy. 2006. http://eosweb.larc.nasa.gov (Accessed 15.05.2006).
[Neshatayev et al.] Нешатаев В.Ю., Потокин А.Ф., Томаева И.Ф., Егоров А.А., Добрыш А.А., Чернядьева И.В., Потемкин А.Д. 2002. Растительность, флора и почвы Верхне-Тазовского государственного заповедника. СПб. 154 с.
[Nitsenko] Ниценко А.А. 1960. Еловые леса Ленинградской области. – Вестн. Ленингр. ун-та. Сер. 3: Биол. 2 (21): 5–16.
Oberdorfer E. 1992. Süddeutsche Pflanzengesellschaften. Teil 4: Wälder und Gebüsche. 2 Aufl. Jena. A. Textband. 282 S. – B. Tabellenband. 580 S.
Påhlsson L. (ed.) 1994. Vegetationstyper i Norden. Köpenhavn. 627 s.
[Parshevnikov, Chertovskoi] Паршевников А.Л., Чертовской В.Г. 1961. Типы еловых лесов и почвы Терского лесхоза. – В кн.: Леса Кольского полуострова и их возобновление. М. С. 40–62.
[Piavchenko] Пьявченко Н.И. 1957. Типы заболачивания лесов в бассейне Северной Двины. – Тр. Ин-та леса АН СССР. 36: 5–55.
[Pismerov et al.] Письмеров А.И., Письмерова Р.С., Воробей Т.М., Тяк А.В. 1986. Лесоводственные особенности древостоев основных групп типов леса. – В кн.: Кологривский лес (Экологические исследования). М. С. 22–33.
[Popov] Попов П.П. 2005. Ель европейская и сибирская: структура, интерградация и дифференциация популяционных систем. Новосибирск. 231 с.
Pott R. 1995. Pflanzengesellschaften Deutschlands. 2 Aufl. Stuttgart. 622 S.
Regel K. 1923. Die Pflanzendecke der Halbinsel Kola. T. I. Lapponia Varsugae. – In: Mémoires de la Faculté des Sciences de L’Université de Lithuanie. Kaunas. 246 S.
[Rutkovskii] Рутковский В.И. 1933. Типы лесов Кемского края АКССР. – Тр. Ин-та изучения леса АН СССР. 1: 1–97.
[Rysin, Savelieva] Рысин Л.П., Савельева Л.И. 2002. Еловые леса России. М. 335 с.
[Saburov] Сабуров Д.Н. 1972. Леса Пинеги. Л. 173 с.
[Safronova, Yurkovskaya] Сафронова И.Н., Юрковская Т.К. 2015. Зональные закономерности растительного покрова равнин Европейской России и их отображение на карте. – Бот. журн. 100 (11): 1121–1141. https://doi.org/10.1134/S0006813615110010
[Salazkin] Салазкин А.С. 1936. Очерк растительности бассейна р. Умбы. – Тр. БИН АН СССР. 3 (3) 69–139.
[Sambuk] Самбук Ф.В. 1932. Печорские леса. – Тр. Бот. музея АН СССР. 24: 63–245.
[Shimanyuk] Шиманюк А.П. 1931. Опыт изучения северных лесов. М.; Л. 104 с.
[Smirnova A.] Смирнова А.Д. 1954. Типы еловых лесов крайнего севера Кировской области. Ч. II. – Учен. записки Горьковск. ун-та. 25: 191–226.
[Smirnova Z.] Смирнова З.Н. 1928. Лесные ассоциации северо-западной части Ленинградской области. – Тр. Петергоф. естеств.-науч. ин-та. 5: 119–258.
[Soczava] Сочава В.Б. 1930. Пределы лесов в горах Ляпинского Урала. – Тр. Бот. музея АН СССР. 22: 1–47.
[Sokolova] Соколова Л.А. 1937. Материалы к геоботаническому районированию Онего-Северодвинского водораздела и Онежского полуострова. – Тр. БИН АН СССР. 3 (2): 9–80.
[Sukachev] Сукачев В.Н. 1931. Руководство к исследованию типов леса. 3-е изд. М. 328 с.
Tukhanen S. 1980. Climatic parameters and indices in plant geography. – Acta Phytogeogr. Suec. 67: 1–105.
Tüxen R. 1937. Die Pflanzengesellschaften Nordwestdeutschlands. – Mitt. flor.-soziol. Arbeitsgem. in Niedersachsen. 3: 1–170.
[Vasilevich, Bibikova] Василевич В.И., Бибикова Т.В. 2004. Сфагновые ельники Европейской России. – Бот. журн. 89 (5): 734–748.
Willner W., Grabherr G. (eds.) 2007. Die Wälder und Gebüsche Österreichs. Ein Bestimmungswerk mit Tabellen. Im 2 Bände. München. 608 S.
[Yudin] Юдин Ю.П. 1948. Темнохвойные леса Коми АССР (геоботаническая характеристика): Дис. … докт. биол. наук. Сыктывкар. 323 с.
[Yudin] Юдин Ю.П. 1954. Темнохвойные леса. – В сб.: Производительные силы Коми АССР. М.; Л. Т. 3. Вып. 1. С. 42–125.
[Yurkevich et al.] Юркевич И.Д., Голод Д.С., Адерихо В.С. 1979. Растительность Белоруссии, ее картографирование, охрана и использование. Минск. 248 с.
[Zinserling] Цинзерлинг Ю.Д. 1932. География растительного покрова северо-запада европейской части СССР. Л. 376 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал