

Ботанический журнал, 2019, T. 104, № 9, стр. 1421-1430
ДИНАМИКА ВИДОВОГО РАЗНООБРАЗИЯ И ПРОДУКТИВНОСТИ ОПУШЕЧНЫХ ФИТОЦЕНОЗОВ АЛАСОВ ЦЕНТРАЛЬНОЙ ЯКУТИИ
М. Х. Николаева 1, *, Р. В. Десяткин 1
1 Институт биологических проблем криолитозоны СО РАН
677980 Якутск, пр. Ленина, 41, Россия
* E-mail: mayan34@yandex.ru
Поступила в редакцию 18.06.2019
После доработки 05.09.2019
Принята к публикации 10.09.2019
Аннотация
Настоящая статья является продолжением серии публикаций по характеристике флуктуаций растительности аласных котловин Центральной Якутии. Исследования проводились в течение 1988–2012 гг. на учетных площадках опушечных фитоценозов у подножий склонов северной экспозиции типичного аласа на стационаре Института биологических проблем криолитозоны СО РАН, растительные сообщества которого формируются на мерзлотных дерново-луговых почвах. Выявлены следующие особенности динамики видового разнообразия и продуктивности (годичной продукции) опушечных фитоценозов: 1) видовой состав растений опушечных фитоценозов не перманентен, межгодовой индекс флористического сходства Жаккара изменяется от 0.27 до 0.82; 2) продуктивность опушечных фитоценозов, в целом, имеет тенденцию к повышению, несмотря на большие межгодовые колебания, составляющие от 2.2 до 27.7 ц/га.
Опушка леса – это переходная полоса между лесом и смежным типом растительности. История изучения опушки как объекта исследований недостаточно богата. В шестидесятые–семидесятые годы прошлого столетия в странах Западной Европы наблюдали повышенный интерес к проблеме опушек. Особенно большой вклад был внесен Тюксеном, Обердорфером, Якучем, которые осветили динамические аспекты и фитотаксономические вопросы изучения опушек (Neshataev et al., 1974). В России отдельные исследования флоры и растительности опушек проведены учеными Красноярского края, Самарского Заволжья, Башкортостана (Maskaev, 1965; Zadulskaya, 1977, 1983, 1990, 1993, 1998; Kukarina, 1997, 1999; Kucherova, Mirkin, 2002; Kukarina et al., 2001).
Наши стационарные исследования динамики видового состава и продуктивности опушечных фитоценозов аласов Центральной Якутии охватывают период 1988–2012 гг. Как правило, склоны северной экспозиции аласных котловин покрыты таежной растительностью, которая простирается до границ с луговой растительности на днищах аласов. Основными лесообразующими породами являются лиственница даурская (Larix dahurica Turcz. ex Trautv.), сосна обыкновенная (Pinus sylvestris L.), береза повислая (Betula pendula Roth). Лиственничники занимают не менее 80% площади лесов (Shcherbakov, 1975).
Результаты исследования закономерностей формирования растительного покрова в аласных котловинах, видового разнообразия и продуктивности влажных, настоящих, остепненных лугов на днищах аласов, а также фитоценозов на склонах котловин южной экспозиции, опубликованы ранее (Nikolaeva, Desyatkin, 2015a, b, 2016a, b). В данной статье мы представляем результаты анализа динамики видового разнообразия и продуктивности опушечных фитоценозов у подножий склонов северной экспозиции аласных котловин в многолетнем цикле.
МАТЕРИАЛ И МЕТОДЫ
Исследования проводили в течение 25 лет на учетных площадках опушки под склоном северной экспозиции типичного аласа Лено-Амгинского междуречья стационара Института биологических проблем криолитозоны СО РАН (ИБПК СО РАН). Склон северной экспозиции покрыт лиственничником багульниковым, по краю днища аласа, и лиственничником брусничным в верхней части склона. На опушке под склоном северной экспозиции аласа на сложение травостоя значительное влияние оказывает затененность, которая защищает растения от сильного иссушения солнцем, а также способствует влагоудержанию. В связи с этим на этом участке создается микроклимат, способствующий развитию настоящих луговых трав.
При проведении исследований использован метод пробных площадей. Описания проводились на площадках 5 м2. Всего сделано 75 описаний. При оценке проективного покрытия использована шкала обилия Браун-Бланке. Величину надземной фитомассы травостоя оценивали методом учетных площадок размером 1 м2 в 4-кратной повторности в воздушно-сухом состоянии.
Опушечные пространства под склоном северной экспозиции модельного аласа занимают небольшую площадь, ежегодная ширина их в зависимости от погодных условий колебалась в пределах 10–20 м по южному периметру аласа. Здесь сформированы дерново-луговые почвы на неоднородных озерно-аласных отложениях. Почвы за лето оттаивают до 0.9–1.3 м. Они достаточно обеспечены влагой в течение всего теплого периода года за счет внутрипочвенной подпитки со стороны склона северной экспозиции заросшего лиственничником багульниковым. Дерново-луговые почвы на неоднородных озерно-аласных отложениях в своем профиле содержат поверхностный и два погребенных горизонта LD озерно-болотного происхождения. Содержание органики в поверхностном горизонте LD составляет 31.3%, в погребенном горизонте LD в средней части профиля почвы – 22.2%. В минеральных горизонтах содержание гумуса колеблется от 0.8 до 9.9% и с глубиной снижается. Реакция почвенной среды в верхнем горизонте щелочная (рН 7.9) и сильнощелочная в нижних минеральных карбонатных горизонтах (рН 8.2–8.8). Емкость обмена в верхних горизонтах средняя (10.3–19.2 мг-экв/100г), в нижних минеральных горизонтах низкая, снижается до 4.0–7.5 мг-экв/100г. Гранулометрический состав верхнего минерального горизонта легкосуглинистый, нижних минеральных – супесчаный, почва не засолена (Desyatkin, 2008).
При проведении исследований видового разнообразия и продуктивности фитоценозов опушечных пространств использованы те же методы, что и при изучении влажных, настоящих, остепненных лугов и фитоценозов склонов южной экспозиции (Nikolaeva, Desyatkin, 2015a, b, 2016a, b). Погодные условия в течение вегетационных сезонов с 1988 по 2012 г. имели существенные колебания. Так, во влажные годы количество осадков за май–сентябрь составляло от 163 до 256 мм, при этом средняя температура воздуха за сезон колебалась от 11.9 до 13.4°С. В экстрасухие годы за сезон выпадало всего 68 и 89 мм осадков (соответственно 2001 и 2002 г.). В засушливые годы количество осадков составляло 103–147 мм, при температурных показателях – 11.9–14.8°С. По метеорологическим условиям летние периоды 11 лет относятся к влажным сезонам (1988, 1989, 1993, 1994, 1996, 1999, 2003, 2005–2007, 2011), 14 – к засушливым (1990–1992, 1995, 1997, 1998, 2000–2002, 2004, 2008–2010, 2012). Изменения погодных условий отражались на видовом составе и продуктивности опушечных фитоценозов аласов.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Состав фитоценозов опушечных пространств аласа за период наблюдений насчитывал 86 видов сосудистых растений. Из них 17 видов относятся к Poaceae, 14 – к Cyperaceae. Разнотравье представлено 55 видами, среди них: 11 – Asteraceae, 5 – Rosaceae, по 4 вида – Fabaceae, Ranunculaceae, Polygonaceae, по 3 вида – Scrophullariaceae, Portulacaceae, Primulaceae, Brassicaceae, по 2 – Rubiaceae, Violaceae, Campanulaceae, Equisetaceae, по 1 виду – Boraginaceae, Geraniaceae, Plantaginaceae, Onagraceae, Juncaceae. Минимальное количество – по 11 видов – в василистниково-гауптобескильницевых (Thalictro-Hordeetum brevisubulati вар. Puccinellia hauptiana) и василистниково-незамечаемовейниковых (Thalictro-Hordeetum brevisubulati вар. Calamorostis neglecta) фитоценозах отмечено в 1988 и 1995 г., максимальное – 43 вида – в василистниково-короткоостистоячменных (Thalictro-Hordeetum brevisubulati вар. typica) фитоценозах в 2002 г. В видовом составе василистниково-короткоостистоячменных фитоценозов доминируют Hordeum brevisubulatum (Trin.) Link11, Poa pratensis L., Alopecurus arundinaceus Poir., Thalictrum simplex L., Potentilla anserina L. В формировании травостоя василистниково-гауптобескильницевых и василистниково-незамечаемовейниковых фитоценозов опушечных пространств также принимают участие Puccinellia hauptiana V. Krecz., Elytrigia repens (L.) Nevski, Calamagrostis neglecta (Ehrh.), Ranunculus propinquus C.A. Mey, Potentilla stipularis L., Mulgedium sibiricum Cass. ex Less., Stellaria dahurica Willd. ex Schlecht, Vicia cracca L.
По встречаемости все виды условно можно разделить на группы: доминирующие – 5 видов; постоянные виды с низким обилием – 6; виды, встречающиеся из 25 лет в течение 10–19 лет – 19; виды, встречающиеся в течение 2–9 лет – 35; единичные виды – 21.
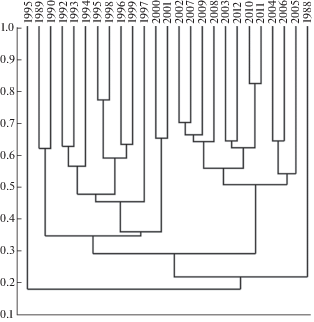
Для сравнения видового состава опушечных фитоценозов по годам применен индекс флористического сходства Жаккара, на основе значений которого построена дендрограмма (рис. 1). На дендрограмме видно, что видовой состав фитоценозов опушечных пространств представлен двумя кластерами, (т.е. годами, объединенными в группы): 1) 1989–1994 и 1996–2001 гг., 2) 2002–2012 гг., при этом 1988 г. и 1995 г. занимают обособленное место, это может быть связано с тем, что 1988 г. относится к влажному, а 1995 год – к засушливому сезону, хотя по количеству видов равны. Индекс сходства видового состава выше 0.8 отмечен во второй группе, между 1997 и 1998, 2002 и 2010 г. В центральной части дендрограммы индекс Жаккара варьировал в пределах 0.6–0.8. В первой группе индекс невысокий, выше 0.4, но ниже 0.6. В третьей группе индекс колебался в широких пределах, от 0.27 до 0.65.
Рис. 1.
Сходство видового состава по годам (индекс Жаккара, метод по средней связи).
По оси абсцисс – годы исследований; по оси ординат – значение индекса Жаккара.
Fig. 1. Similarity of species composition by year (Jaccard index, “paired group”).
X-axis – observation years; Y-axis – values of Jaccard index.
Во влажном 1988 г. видовое разнообразие василистниково-гауптобескильницевых (Thalictro-Hordeetum brevisubulati вар. Puccinellia hauptiana) фитоценозов оказалось невысоким, всего отмечено 11 видов, при этом проективное покрытие Puccinellia hauptiana составила 50–75%, Alopecurus arundinaceus 1–5%, Thalictrum simplex – 5–20%. В этот год видовой состав дополняли Artemisia tanacetifolia L. c проективным покрытием 20–50%, Gallium verum L., Achillea millefolium L., Taraxacum officinale Wigg. с проективным покрытием 1–5% каждый. В следующем влажном 1989 г. видовое разнообразие увеличилось до 22 видов. Появились виды Hordeum brevisubulatum и Potentilla anserina L. с проективным покрытием по 20–50%, Poa pratensis, Elytrigia repens, Potentilla stipularis с проективным покрытием по 5–20%, Stellaria dahurica, Knorringia sibirica (Laxm.) Tzvel, Anemone sylvestris L., Lychnis sibirica L., Chamerion angustifolium (L.) Holub с проективным покрытием 1–5% каждый вид. В последующие более сухие годы c 1990 по 1992 проективное покрытие Hordeum brevisubulatum и Puccinellia hauptiana постепенно снизилось до 5–20%, а проективное покрытие Elytrigia repens и Alopecurus arundinaceus повысилось до 20–50%. В эти же сухие годы проективное покрытие Thalictrum simplex снизилось до 1–5% (1990 г.), в следующий 1991 г. покрытие повысилось до 20–50%, затем в последующий год снизилось до 5–20%. Проективное покрытие Potentilla anserina снизилось до 5–20% и оставалось на одном уровне. В эти же годы появились Vicia cracca L. и Taraxacum ceratophorum (Ledeb.) DC с проективным покрытием 5–20% каждый вид, Carex reptabunda (Trautv.) V. Krecz. с проективным покрытием 1–5%.
Во влажный 1994 г. проективное покрытие доминирующих видов Hordeum brevisu-bulatum, Alopecurus arundinaceus, Potentilla anserina оставалось на уровне 20–50%, Poa pratensis – 1–5%, а Thalictrum simplex увеличилось до 50–75%. Отмечено появление Calamogrostis neglecta (Ehrg.) Gaertn с проективным покрытием 5–20%. В сухие 1995, 1998 годы обилие Potentilla anserina существенно повысилось – до 20–50%, а проективное покрытие Alopecurus arundinaceus, Puccinellia hauptiana снизилось до 15%. Во влажные 1996, 1997, 1999 гг. проективное покрытие Alopecurus arundinaceus и Potentilla anserina составило от 20 до 50%, Thalictrum simplex и Hordeum brevisubulatum от 5 до 50%, Poa pratensis от 1 до 5%. Однако четкой зависимости видового разнообразия опушечных фитоценозов от погодных условий не наблюдалось. Тем не менее в сухие годы отмечено снижение проективного покрытия доминанта Alopecurus arundinaceus, тогда как проективное покрытие Poa pratensis повысилось.
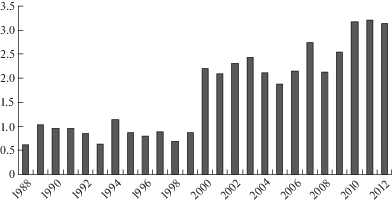
В зависимости от метеорологических условий вегетационного сезона того или иного года, площадь опушечных фитоценозов аласа ежегодно менялась и варьировала от 0.61 (1988 г.) до 3.21 га (2011 г.), при этом 1988 и 2011 г. по метеоусловиям относились к влажным (рис. 2). На изменение параметров площади опушки оказывало влияние и изменение площади водного зеркала озера. Так, за годы наблюдений, в 1988 г. площадь водного зеркала озера была минимальной (0.03 га). Начиная с 2005 г., площадь озера расширилась и достигла максимального значения в 2008 г. (6.89 га). Между площадью опушечной полосы и площадью водного зеркала озера выявлена прямая коррелятивная связь (r = 0.78).
Рис. 2.
Динамика площади опушечных фитоценозов модельного аласа в период 1988–2012 гг.
По оси абсцисс – годы исследований; по оси ординат – площадь, га.
Fig. 2. Dynamics of the area of model alas margin phytocenoses during 1988–2012.
X-axis – observation years; Y-axis – area, ha.
За период наблюдений продуктивность опушечных фитоценозов аласа имела значительные колебания по годам (рис. 3). Диапазон продуктивности варьировал от 2.2 до 27.7 ц/га воздушно-сухого вещества. Продуктивность имела слабую положительную зависимость от количества выпадающих осадков в первой половине летнего сезона этого года. Майские осадки выпадают при еще достаточно мерзлой почве, поэтому пропорционально не влияли на рост растений. Большее влияние оказывали осенние осадки (август–сентябрь) предыдущего теплого сезона. Как правило, влага, аккумулированная в почве в это время, консервируется сезонной мерзлотой до начала следующего вегетационного периода и начинает расходоваться растительностью по мере оттаивания в первой половине вегетационного сезона последующего года.
Рис. 3.
Динамика продуктивности опушечных фитоценозов, ц/га.
По оси абсцисс – годы исследований; по оси ординат – продуктивность, ц/га (воздушно-сухое вещество).
Fig. 3. Dynamics of productivity of margin phytocenoses, centner/ha.
X-axis – observation years; Y-axis – productivity, centner/ha (air-dry matter).
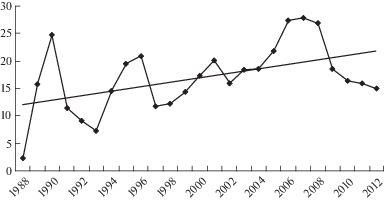
Минимальное значение продуктивности отмечено в 1988 г., которая в основном определялась удельными фитомассами Puccinellia hauptiana (1.5 ц/га), Artemisia tanacetifolia L. (0.52 ц/га), Thalictrum simplex (0.05 ц/га) и Achillea millefolium L. (0.07 ц/га). Максимум продуктивности фитоценозов опушечных пространств приходится на 2007 г. Наибольшую удельную фитомассу имели Poa pratensis (4.2 ц/га), Elytrigia repens (7.14 ц/га), Hordeum brevisubulatum (3.26 ц/га), Alopecurus arundinaceus (2.78 ц/га), Thalictrum simplex (1.53 ц/га).
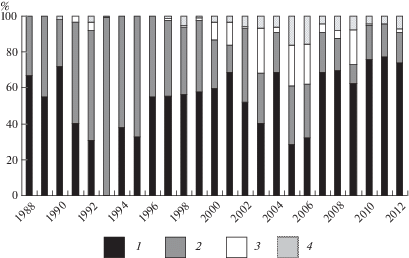
По данным анализа соотношения биологических групп в составе травостоя опушечных фитоценозов по годам видно, что злаки составляли больше 50% от общей фитомассы травостоя, за исключением 1991, 1992, 1994, 1995, 2003, 2005, 2006 гг., когда их доля колебалась в пределах 28.6–40.1%.
В 1993 году присутствие Poaceae не отмечено, так как в этом году была вспышка численности саранчовых, что привело к полному уничтожению их к началу июля. Небольшое снижение доли Poaceae и в 1994 г., возможно, связано с последствиями этой вспышки. На уменьшение доли злаков в 2003, 2005, 2006 гг. оказали влияние засушливые условия предыдущих 2002 и 2004 г. Доля разнотравья выше 50% отмечена в 1991–1995 гг. и составляла 56.4, 61.3, 99.7, 62.0, 67.5% соответственно (рис. 4). Во влажный 1994 г., после вспышки численности саранчовых, вегетация злаков на почвах, удобренных экскрементами саранчовых, была эффективной, поэтому доля разнотравья оказалась на уровне 1992 года. Участие осоковых отмечено за 16 лет, из которых 11 лет относились к сухим, 5 лет – к влажным сезонам. При этом доля осоковых при недостаточном увлажнении достигала 19.5%, во влажные годы – 25.3%. Повышение доли осоковых происходило при сравнительном понижении доли злаков и повышении доли разнотравья из бобовых. В годы с достаточным увлажнением доля разнотравья из бобовых достигала 16.0%, а в засушливые годы снижалась в два раза (до 8.0%).
Рис. 4.
Участие ботанических групп в надземной фитомассе опушечных фитоценозов: злаки (1), разнотравье (2), осоки (3), разнотравье (Fabaceae) (4).
По оси абсцисс – годы исследований; по оси ординат – участие ботанических групп, %.
Fig. 4. Share of agrobotanical groups in above-ground phytomass of margin phytocоenoses: grasses (1), forbs (2), sedges (3), forbs (Fabaceae) (4).
X-axis – observation years; Y-axis – percentage of botanical groups.
ЗАКЛЮЧЕНИЕ
В ходе проведенных исследований выявлены следующие особенности динамики видового разнообразия и продуктивности опушечных фитоценозов аласов Центральной Якутии:
Выявлена существенная межгодовая динамика видового состава опушечных фитоценозов, что обусловлено изменениями погодных условий разных лет и связанное с этим варьирование микроклиматического и гидротермического режимов опушечных местообитаний. Индекс флористического сходства Жаккара варьировал от 0.27 до 0.82.
Обнаружены значительные изменения продуктивности опушечных фитоценозов от 2.2 до 27.7 ц/га. Кроме выше отмеченных причин динамики продуктивности, здесь заметную роль играет и другой экзогенный фактор – массовые вспышки численности саранчовых, которые циклически проявляются в условиях аласных экосистем. В годы максимума численности саранчовых количество белополосой кобылки достигает 1000 и более особей на 1 м2 (Desyatkin, 2008). В такие годы продуктивность опушечных фитоценозов минимальна.
По результатам анализа соотношения биологических групп выявлено, что за 18 лет из 25-ти доля Poaceae составляла выше 50% от общей фитомассы (в основном, это отмечено в сухие годы, за исключением 2 влажных лет). Доля осоковых в годы с недостаточным увлажнением (11 лет) снижалась до 19.5%, достигая во влажные (5 лет) – 25.3%. В годы с достаточным увлажнением доля разнотравья из бобовых достигала 16.0%, а в засушливые годы снижалась в два раза. После перехода от сухих к влажным или наоборот от влажных к сухим годам наблюдалось увеличение доли разнотравья, которая превышала 50% от общей фитомассы 5 раз в течение изученного периода.
Список литературы
[Desyatkin] Десяткин Р.В. 2008. Почвообразование в термокарстовых котловинах – аласах криолитозоны. Новосибирск. 324 с.
[Kuznetsova, Zakharova] Кузнецова Л.В., Захарова В.И. 2012. Конспект флоры Якутии: сосудистые растения. Новосибирск. 272 с.
[Kukarina] Кукарина С.В. 1997. Особенности ксеротермных опушек лесов юга Башкортостана. – В сб.: Леса Башкортостана: современное состояние и перспективы: Материалы науч. – практ. конф. Уфа. С. 49–50.
[Kukarina] Кукарина С.В. 1999. Флора опушек южного берега озера Асликуль. – В сб.: Фауна и флора Республики Башкортостан: Материалы докл. науч. конф. Уфа. С. 210–214.
[Kukarina et al.] Кукарина С.В., Миркин Б.М., Соломещ А.И. 2001. Фитосоциологический градиентный анализ лесных опушек как экотонов. – Бюл. МОИП. Отд. биол. 106(1): 61–69.
[Kucherova, Mirkin] Кучерова С.В., Миркин Б.М. 2002. Влияние экотонного эффекта на флористическое разнообразие лесных опушек. – В сб.: Сохранение и воспроизводство растительного компонента биоразнообразия: Материалы межд. конф., посвященной 75-летию ботанического сада Ростовского гос. ун-та. Ростов-на-Дону. С. 101–104.
[Maskaev] Маскаев Ю.М. 1965. Фитоклиматические особенности на опушках разных экспозиций на восточной окраине Каннской лесостепи. – В сб.: Растительный покров Красноярского края. Новосибирск. Вып. 2. С. 187–212.
[Neshataev et al.] Нешатаев Ю.Р., Витко К.Р. (Рец.) Х. Диршке. 1978. Сообщества лесных опушек по градиентам растительности и местообитаний. – Бот. журн. 63(5): 769–773.
[Nikolaeva, Desyatkin] Николаева М. Х., Десяткин Р. В. 2015а. Динамика видового разнообразия и продуктивности влажных лугов аласов Центральной Якутии. – Раст. ресурсы. 51(1): 70–80.
[Nikolaeva, Desyatkin] Николаева М. Х., Десяткин Р. В. 2015б. Динамика видового разнообразия и продуктивности настоящих лугов аласов Центральной Якутии. – Раст. ресурсы. 51(3): 328–335.
[Nikolaeva, Desyatkin] Николаева М.Х., Десяткин Р.В. 2016а. Динамика видового разнообразия и продуктивности остепненных лугов аласов Центральной Якутии. – Раст. ресурсы. 52(1): 20–27.
[Nikolaeva, Desyatkin] Николаева М.Х., Десяткин Р.В. 2016б. Динамика видового разнообразия и продуктивности фитоценозов степных склонов аласов Центральной Якутии. – Раст. ресурсы. 52(3): 351–360.
[Shcherbakov] Щербаков И.П. 1975. Лесной покров Северо-Востока СССР. Новосибирск. 344 с.
[Zadulskaya] Задульская О.А. 1977. Опыт флороценотического исследования лесных опушек на территории Самарской Луки. – В сб.: Морфология и динамика растительного покрова: Науч. тр. Куйбышев. педин-та. Куйбышев. 207(6): 93–97.
[Zadulskaya] Задульская О.А. 1983. Флора опушек псаммофитных лесов Куйбышевского Заволжья. – В сб.: Сложение и динамика растительного покрова: Науч. тр. Куйбышев. пед. ин-та. Куйбышев. С. 125–128.
[Zadulskaya] Задульская О.А. 1990. Растительность лесных опушек Куйбышевского Заволжья: Автореф. дис. … канд. биол. наук. Ленинград. 17 с.
[Zadulskaya] Задульская О.А. 1993. Анализ флоры лесных опушек Самарского Заволжья. – В сб.: Флористические исследования в Поволжье и на Урале: Межвуз. сб. Самара. С. 66–71.
[Zadulskaya] Задульская О.А. 1998. Классификация лесных опушек. – В сб.: Проблемы ботаники на рубеже XX–XXI веков: Тез. докл., представленных II(X) съезду Русского ботанического общества. Санкт-Петербург. Т. 1. С. 252.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал