

Ботанический журнал, 2020, T. 105, № 4, стр. 348-359
МОРФОЛОГИЧЕСКАЯ И ГЕНЕТИЧЕСКАЯ ИЗМЕНЧИВОСТЬ ЦЕНОПОПУЛЯЦИЙ ANEMONE JENISSEENSIS (RANUNCULACEAE) НА ЮГЕ ПРИЕНИСЕЙСКОЙ СИБИРИ
А. Н. Мариничева 1, *, И. Е. Ямских 1, Н. В. Степанов 1, М. Г. Куцев 1, 2
1 Сибирский федеральный университет
660041 Красноярск, пр. Свободный, 79, Россия
2 Алтайский государственный университет
656049 Барнаул, ул. Ленина, 61, Россия
* E-mail: alexandra.marinicheva@outlook.com
Поступила в редакцию 01.11.2019
После доработки 22.01.2020
Принята к публикации 28.02.2020
Аннотация
Изучена морфологическая и генетическая изменчивость 10 природных ценопопуляций сибирского эндемика Anemone jenisseensis (Korsh.) Krylov, произрастающих в растительных сообществах Красноярской лесостепи, Западного и Восточного Саяна. Установлено, что данный вид обладает относительно широким фитоценотическим ареалом и может встречаться в мелколиственных (березовые, осиновые леса), светлохвойных (сосняки), смешанных лесах, в пойменных ивняках, а также на вырубках. Среднее количество побегов на 1 м2 варьирует от 15 до 31 шт. Максимальные размеры вегетативных органов, показатели семенной продуктивности и генетического разнообразия вида наблюдаются в подтаежных лесах Западного Саяна и в Красноярской лесостепи. На вырубках отмечается достоверное снижение морфологических показателей Anemone jenisseensis, однако уровень внутрипопуляционной модификационной изменчивости и показатели семенной продуктивности варьируют в пределах средних значений. Генетическую изменчивость ценопопуляций Anemone jenisseensis выявляли с помощью ISSR-PCR (Inter Simple Sequence Repeats) метода. Суммарный процент полиморфных локусов составляет 100. Изученные ценопопуляции характеризуются средним уровнем внутрипопуляционного генетического разнообразия (55.86–71.17%) и очень высокой степенью дифференциации (Gst = 0.3316). На дендрограммах сходства, построенных на основе морфологических и генетических данных, достоверно выделяется группа ценопопуляций, характеризующаяся крупными, глубоко расчлененными боковыми сегментами листа. Разделения ценопопуляций по географическому принципу не выявлено.
Одной из наиболее интересных и относительно слабо изученных в Южной Сибири является группа весенних эфемероидов, принадлежащих к роду Аnemone: Аnemone reflexa Steph., A. jenisseensis (Korsh.) Krylov, A. altaica (C.A. Mey) Holub, Anemone саerulea DC. Данные виды относятся к числу эндемиков, третичных неморальных реликтов и занесены в сводки редких и исчезающих видов растений различного уровня.
Объектом нашего исследования является сибирский эндемик A. jenisseensis. Распространена в пределах Красноярского края, Хакасии. Встречается в лесах Томской, Кемеровской, Иркутской областей. Растение включено в “Красную книгу Иркутской области” (Red…, 2010) – категория 2 (V), уязвимый вид.
Для разработки методов сохранения генетического разнообразия редких видов необходимы сведения о полиморфизме и генетической структуре их ценопопуляций (Filippov, Andronova, 2017). Работы по изучению генетического разнообразия представителей рода Аnemone проводились преимущественно за рубежом (Schuettpelz et al., 2002; Cai et al., 2009; Mlinarec et al., 2011; Sun et al., 2012; Zhang et al., 2015; Jiang et al., 2017), и, лишь в некоторых случаях, в Сибири (Marinicheva, Yamskikh, 2017; Yamskikh et al., 2011). Среди сибирских видов ветрениц менее всего изучена Anemone jenisseensis. В настоящее время отсутствуют сведения о морфологической и генетической структуре популяций, внутривидовой дифференциации данного вида.
A. jenisseensis относится к семейству Ranunculaceae и представляет собой многолетнее травянистое растение с членистым, распадающимся, мясистым корневищем (Luferov, Starodubtsev, 1995). В естественных условиях ветреница енисейская размножается, преимущественно, вегетативным способом, путем отмирания более старых участков корневищ и обособления самостоятельных особей (Barykina, 1995).
Цель исследования – изучение морфолого-генетической изменчивости природных ценопопуляций (ЦП) A. jenisseensis.
МАТЕРИАЛЫ И МЕТОДЫ
Сбор материала проводился в период 2007–2018 гг. в Красноярской лесостепи, в горах Западного и Восточного Саяна (рис. 1). Объектом исследования явились 10 ценопопуляций A. jenisseensis. На первом этапе исследования изучали фитоценотическую приуроченность ветреницы. В исследуемом районе описано свыше 50 местообитаний вида. Геоботанические описания проведены по общепринятым методикам (Voronov, 1973). При изучении состояния ценопопуляций ветреницы енисейской определяли проективное покрытие, плотность, оценивали изменчивость вегетативных и генеративных признаков, семенную продуктивность, изучали генетическую вариабельность на основе ISSR-PCR анализа. Характеристика местообитаний вида приведена в табл. 1.
Таблица 1.
Краткая характеристика сообществ ценопопуляций Anemonе jenisseensis Table 1. Brief description of communities with Anemone jenisseensis coenopopulations
| № ц/п Coenopo-pulation number |
Фитоценоз; местоположение Phytocoenosis; location |
Состав древостоя; сомкнутость крон (%) Stand composition; crown density (%) |
Доминирующие виды травяного яруса Dominants of the grass cover |
|---|---|---|---|
| Западный Саян/Western Sayan | |||
| Aj1 | Сосняк орляково-разнотравный (долина р. Уй, окр. пос. Майна, Республика Хакасия) Pine forest with bracken and grass cover (Uy River valley, Mayna village, Republic of Khakassia) |
9С1Б Pinus sylvestris – 90% Betula pendula – 10%, 0.4–0.5 |
Carex macroura (30%) Aconitum barbatum (20%) Anemone jenisseensis (10%) Calamagrostis arundinacea (10%) Pteridium pinetorum (5%) |
| Aj2 | Березово-сосновый лес орляково-разнотравно-злаковый (окр. с. Ермаковское, Ермаковский
р-н, Красноярский край) Birch-pine forest with bracken and mixed herb-grass cover (Yermakovskoye village, Krasnoyarsk Territory) |
4Б6С Pinus sylvestris – 60% Betula pendula – 40%, 0.5 |
Calamagrostis arundinacea (30%) Pteridium pinetorum (25%) Brunnera sibirica (10%) Anemone jenisseensis (1%) |
| Aj3 | Березово-сосновый лес хвощово-разнотравный (д. Григорьевка, Ермаковский р-н, Красноярский
край) Birch-pine forest with horsetail and mixed herb cover (Grigoryevka village, Krasnoyarsk Territory) |
8С2Б Pinus sylvestris – 80% Betula pendula – 20%, 0.7 |
Equisetum sylvaticum (20%) Brachypodium pinnatum (15%) Agrimonia pilosa (8%) Anemone jenisseensis (5%) Brunnera sibirica (4%) |
| Aj4* | Луг злаково-орляковый (окр. д. Осиновка, Ермаковский р-н, Красноярский край) Meadow with grasses and bracken cover (Osinovka village, Krasnoyarsk Territory) |
Древостой отсутствует Forest stand absent |
Pteridium pinetorum (60%) Calamagrostis obtusata (60%) Dactylis glomerata (15%) Anemone jenisseensis (1%) |
| Aj5 | Пойменный ивняк широкотравно-страусниковый (окр. п. Танзыбей (Червизюльский тракт,
3-й км), Ермаковский р-н, Красноярский край) Willow floodplain forest with broad-leaved grasses and ostrich fern cover (Tanzybey village (Chervizyulskiy road, 3rd km), Krasnoyarsk Territory) |
Древостой отсутствует Forest stand absent |
Matteuccia struthiopteris (70%) Brunnera sibirica (20%) Anemone baicalensis (10%) Anemone jenisseensis (2%) |
| Aj6 | Березово-сосновый лес папоротниково-широкотравный (окр. д. Осиновка, Ермаковский р-н,
Красноярский край); Birch-pine forest with fern and broad-leaved herb cover (Osinovka village, Krasnoyarsk Territory) |
6С4Б Pinus sylvestris – 60% Betula pendula – 40%, 0.5 |
Brunnera sibirica (60%) Pteridium pinetorum (40%) Athyrium monomachii (15%) Matteuccia struthiopteris (10%) Viola uniflora (5%) Anemone jenisseensis (1%) |
| Aj7 | Березово-еловый лес разнотравно-осочковый (окр. с. Усинское (пойма р. Иджим), Красноярский
край); Birch-spruce forest with mixed herb and sedge cover (Usinskoye village (Idzhim River valley), Krasnoyarsk Territory) |
8Б2Е Betula pendula – 80% Picea obovata – 20%, 0.5 |
Carex cespitosum (40%) Deschampsia cespitosa (10%) Filipendula ulmaria (8%) Viola uniflora (7%) Anemone jenisseensis (1%) |
| Восточный Саян/Eastern Sayan | |||
| Aj8 | Осиново-березовый лес широкотравно-страусниковый (пойма р. Лалетина,
возле заповедника “Столбы”, окр. г. Красноярск, Красноярcкий край) Aspen-birch forest with broad-leaved herb and ostrich fern cover (Laletina River valley, Stolby Nature Reserve, Krasnoyarsk, Krasnoyarsk Territory) |
8Б2Ос Betula pendula – 80% Populus tremula – 20%, 0.4–0.5 |
Matteuccia struthiopteris (15%) Heracleum dissectum (10%) Corydalis bracteata (10%) Anemone jenisseensis (2%) |
| Красноярская лесостепь/Krasnoyarsk forest-steppe | |||
| Aj9 | Березово-осиновый лес разнотравно-орляковый (Николаевская сопка,
г. Красноярск, Краснояркий край) Birch-aspen forest with mixed herb and bracken fern cover (Nikolaevskaya Sopka, Krasnoyarsk, Krasnoyarsk Territory) |
6Ос4Б Populus tremula – 60% Betula pendula – 40%, 0.5-0.6 |
Pteridium pinetorum (60%) Carex macroura (15%) Brachypodium pinnatum (10%) Anemone jenisseensis (1%) |
| Aj10* | Луг высокотравно-злаковый (окр. ст. Рябинино, окр. г. Красноярск, Красноярский край) Meadow with tall herb and grass cover (Ryabinino railway station, Krasnoyarsk, Krasnoyarsk Territory) |
Древостой отсутствует Forest stand absent |
Poa pratensis (30%) Dactylis glomerata (30%) Trollius asiaticus (25%) Filipendula ulmaria (20%) Anemone jenisseensis (3%) |
Измерения параметров вегетативных органов и показателей семенной продуктивности ветреницы для оценки фенотипической изменчивости проводили на 30 генеративных побегах. Большая часть оцениваемых нами морфометрических признаков приводится в качестве диагностических для определения видов во “Флоре СССР” (Yuzepchuk, 1937), “Флоре Средней Сибири” (Popov, 1957), “Флоре Сибири” (Timokhina, 1993). Кроме метрических, использовался и ряд аллометрических параметров, оценивающих соотношение в развитии разных частей растений. Показатели семенной продуктивности определяли по методике Г.П. Дюрягиной и М.М. Ивановой (Dyuryagina, Ivanova, 1985). У побегов ветреницы енисейской измерялись следующие параметры: длина стебля (×1), длина цветоноса (×2), длина черешка (×3), длина (×4) и ширина (×5) центрального сегмента листа, соотношение длины и ширины листа (×6), соотношение длины стебля и длины листа (×7), глубина выреза на центральном сегменте (×8), соотношение длины выреза и длины листа (×9), количество зубчиков (×10), длина (×11) и ширина (×12) бокового сегмента листа, количество зубчиков на боковом сегменте (×13), длина выреза на боковом сегменте (×14), соотношение длины выреза и длины листа (×15), длина (×16) и ширина (×17) лепестков, количество цветков (×18), реальная (с1) и потенциальная (с2) семенные продуктивности; коэффициент семенификации (с3).
При математической обработке данных рассчитывали пределы варьирования признака (min-max), среднее арифметическое (X) и его ошибку (mx), среднее квадратичное отклонение (σ), коэффициент вариации (Cv). Характер распределения значений признаков ветреницы енисейской оценивали с помощью коэффициентов асимметрии и эксцесса. Выявлено, что для изучаемых нами параметров характерно распределение, близкое к нормальному. Внутрипопуляционную изменчивость признаков оценивали с помощью коэффициента вариации (Cv), поскольку данный параметр позволяет сравнивать признаки, имеющие различные размерности (Schmidt, 1984), кроме того, для коэффициента вариации С.А. Мамаевым (Mamaev, 1972) разработана шкала уровней изменчивости. Для установления достоверных различий между среднепопуляционными значениями одноименных признаков A. jenisseensis использовался однофакторный дисперсионный анализ. Различие считалось достоверным при уровне значимости p < 0.05. Анализ сходства ценопопуляций по исследуемым признакам проводили с помощью кластерного анализа. В обработке применялся метод Уорда (Ward, 1963). В качестве меры сходства использовали Евклидово расстояние.
Генетическую вариабельность ценопопуляций вида изучали с помощью ISSR-PCR (Inter Simple Sequence Repeat) метода, выявляющего полиморфизм межмикросателлитных последовательностей ДНК. Из каждой популяции было проанализировано 10 растений, не являющихся клонами. Геномную ДНК выделяли из 10 мг материала, высушенного в силикагеле, с использованием набора DiamondDNA (ООО “АБТ”, Россия). Для ПЦР использовали смесь БиоМастерHS‑Taq ПЦР-Color 2x(ООО “Биолабмикс”, г. Новосибирск).
ISSR-PCR проводили в 20 мкл смеси (6 мкл Н2О; 10 мкл смеси БиоМастер HS-Taq ПЦР-Color 2x, 2 мкл ДНК; 2 мкл 10 мМ праймера) на амплификаторе C1000 Thermal Cycler BioRad с использованием реактивов производства OOO “БиолабМикс” (г. Новосибирск) и ЗАО “Евроген” (г. Москва). Амплификацию проводили по следующей программе: 95°С – 5 мин; 13 циклов: 95°С – 20 сек, 55°С – 45 сек с последующим понижением температуры на 0.7°С в каждом цикле, 72°С – 90 сек; 25 циклов: 95°С – 20 сек, 44°С – 30 сек, 72°С – 90 сек; завершающая стадия: 72°С – 10 мин, охлаждение при 4°С. Предварительно на 2 образцах ДНК из имеющегося набора праймеров (Mansour et al., 2009; Paterson et al., 2009) опытным путем были выявлены те, которые дают воспроизводимый полиморфный результат. Анализ продуктов амплификации осуществляли в проходящем ультрафиолетовом свете с помощью системы гель-документирования и анализа изображений Bio-Rad Gel Doc XR (Bio-Rad, USA).
Для количественной оценки степени полиморфизма и определения уровня дивергенции между изучаемыми популяциями полученные данные были представлены в виде матрицы бинарных признаков, в которой наличие или отсутствие ISSR-ПЦР фрагментов одинакового размера рассматривалось как состояние 1 или 0 соответственно. При этом учитывали только воспроизводимые в повторных экспериментах фрагменты. Обработку результатов анализа проводили с помощью пакета программ TFPGA version 1.3 (Miller, 1997) (UPGMA-анализ, бутстреп-тест и расчет генетических дистанций Рейнольда) и Popgene version 1.32 (процент полиморфных локусов (P), генетическое разнообразие Нея (I), информационный индекс Шеннона (h), генетические дистанции Нея (D), показатель подразделенности популяций (Gst)).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
При оценке фитоценотической приуроченности выявлено, что ветреница енисейская в Восточном, Западном Саяне и в окр. г. Красноярска распространена в березовых, осиновых, сосновых и смешанных лесах, а также в поймах рек и на вырубках. Проективное покрытие A. jenisseensis в растительных сообществах варьирует от 1 до 10%. Максимальное значение данный показатель имеет в западносаянском сосняке орляково-разнотравном (Aj1, 10%). Также относительно высокие показатели численности ветреницы были зафиксированы в березово-сосновом лесу хвощово-разнотравном (Aj3, 5%, Западный Саян). На вырубке сосняка со следами пожара (Aj4*) отмечено резкое снижение проективного покрытия особей вида до 1%. В растительных сообществах вид выполняет роль ассектатора.
Среднее количество побегов ветреницы енисейской на 1 м2 варьирует от 15 до 31 шт. Максимальная плотность A. jenisseensis зафиксирована для ЦП, произрастающей в окр. г. Красноярска (Аj9) – 30.7 ± 5.7 шт./м2. В осиново-березовом лесу (Аj8) отмечено 24.2 ± 5.8 побегов на 1 м2. Минимальное число побегов было выявлено на вырубке под ЛЭП (Aj10*, 14.6 ± 4.1 шт./м2) и в березово-сосновом лесу орляково-разнотравно-злаковом в окр. с. Ермаковского, Западный Саян (Aj2, 15 ± 2.5 шт./м2).
В результате изучения внутрипопуляционной изменчивости установлено, что большинство вегетативных признаков вида характеризуются средним и высоким уровнями изменчивости, согласно шкале С.А. Мамаева (Mamaev, 1972). Наименее изменчивыми признаками для побегов ветреницы являются длина стебля, длина цветоноса, длина черешка, длина центрального и бокового сегментов листа, форма центрального сегмента листа (рис. 2). Высокой степенью варьирования характеризуются степень и характер рассеченности центрального и боковых сегментов листа – от почти цельных до раздельных (рис. 3). Причем растения с разными типами расчленения листьев могут встречаться в пределах одной ценопопуляции. Три западносаянские ЦП (Aj1, Aj4* и Aj6) характеризуются глубоко расчлененными боковыми сегментами листьев.
Рис. 2.
Внутрипопуляционная изменчивость морфологических признаков Anemone jenisseensis: ×2 – длина цветоноса, ×4 – длина центрального сегмента листа, ×6 – форма центрального сегмента листа, ×9 – степень расчленения центрального сегмента, ×15 – степень расчленения бокового сегмента.
Fig. 2. Intrapopulation variability of morphological characters of Anemone jenisseensis: ×2 – peduncle length, ×4 – central leaf segment length, ×6 – central leaf segment shape, ×9 – degree of incision of the central leaf segment, ×15 – degree of incision of the lateral leaf segment.
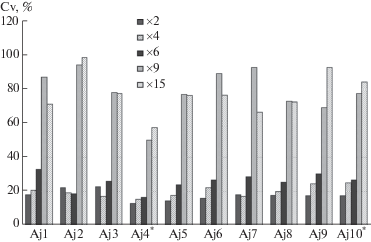
Рис. 3.
Особи ветреницы енисейской с разной степенью расчленения листовых пластинок (фото Н.В. Степанова).
Fig. 3. Plants of Anemone jenisseensis with different degree of leaf blade incision (photo by N.V. Stepanov).

Наибольшими размерами осевых органов (х1-х3) характеризуются особи, произрастающие в сосновых сообществах Западного Саяна (табл. 2): в сосняке орляково-разнотравном (Aj1), березово-сосновом лесу орляково-разнотравно-злаковом (Aj2) и березово-сосновом лесу хвощово-разнотравном (Aj3). Например, длина стебля у особей ЦП Aj2 составляет 23.38 см, а длина цветоноса – 5.49 см. Максимальные размеры листьев зафиксированы для западносаянских ЦП Aj3 и Aj5, а также для Aj9, находящейся в окр. г. Красноярска. Минимальные показатели всех вегетативных признаков характерны для растений ветреницы енисейской, произрастающих на вырубке под ЛЭП (Aj10*) (табл. 2).
Таблица 2.
Среднепопуляционные значения признаков Anemone jenisseensis Table 2. Average population values of Anemone jenisseensis characters
| Aj1 | Aj2 | Aj3 | Aj4* | Aj5 | Aj6 | Aj7 | Aj8 | Aj9 | Aj10* | |
|---|---|---|---|---|---|---|---|---|---|---|
| Длина стебля, см (×1) Stem length, cm (×1) |
23.20 ± 0.6 | 23.38 ± 0.56 | 23.04 ± 0.63 | 18.35 ± 0.53 | 22.75 ± 0.48 | 22.34 ± 0.48 | 21.27 ± 0.58 | 18.29 ± 0.62 | 21.62 ± 0.68 | 20.66 ± 0.58 |
| Длина цветоноса, см (×2) Peduncle length, cm (×2) |
4.32 ± 0.14 | 5.49 ± 0.22 | 5.26 ± 0.22 | 4.94 ± 0.12 | 5.65 ± 0.15 | 5.66 ± 0.16 | 5.92 ± 0.19 | 4.24 ± 0.14 | 4.43 ± 0.14 | 4.29 ± 0.14 |
| Длина черешка, см (×3) Petiole length, cm (×3) |
0.61 ± 0.02 | 0.69 ± 0.03 | 0.6 ± 0.02 | 0.64 ± 0.03 | 0.73 ± 0.03 | 0.75 ± 0.03 | 0.6 ± 0.03 | 0.58 ± 0.02 | 0.5 ± 0.03 | 0.58 ± 0.02 |
| Длина центрального сегмента листа, см (×4) Central leaf segment length, cm (×4) |
5.31 ± 0.2 | 5.02 ± 0.18 | 5.91 ± 0.18 | 4.66 ± 0.13 | 5.41 ± 0.17 | 5.02 ± 0.2 | 4.53 ± 0.14 | 4.91 ± 0.18 | 5.36 ± 0.24 | 3.52 ± 0.16 |
| Ширина центрального сегмента листа, см (×5) Central leaf segment width, cm (×5) |
0.81 ± 0.06 | 0.69 ± 0.03 | 0.88 ± 0.05 | 0.8 ± 0.04 | 0.73 ± 0.03 | 0.78 ± 0.04 | 0.69 ± 0.04 | 0.95 ± 0.07 | 0.94 ± 0.06 | 0.61 ± 0.03 |
| Глубина выреза на центральном сегменте, см (×8) Depth of central segment cut, cm (×8) |
0.46 ± 0.08 | 0.15 ± 0.03 | 0.34 ± 0.05 | 0.37 ± 0.04 | 0.31 ± 0.05 | 0.27 ± 0.05 | 0.34 ± 0.07 | 0.52 ± 0.08 | 0.43 ± 0.06 | 0.27 ± 0.04 |
| Степень расчленения центрального сегмента (×9) Degree of central segment incision (×9) |
0.09 ± 0.01 | 0.03 ± 0.01 | 0.06 ± 0.01 | 0.08 ± 0.01 | 0.06 ± 0.01 | 0.05 ± 0.01 | 0.07 ± 0.01 | 0.10 ± 0.01 | 0.08 ± 0.01 | 0.07 ± 0.01 |
| Количество зубчиков, шт (×10) Number of teeth, pcs (×10) |
7.23 ± 0.37 | 5.10 ± 0.52 | 7.70 ± 0.64 | 7.27 ± 0.46 | 7.47 ± 0.47 | 7.93 ± 0.7 | 6.07 ± 0.33 | 6.57 ± 0.4 | 6.37 ± 0.41 | 5.73 ± 0.34 |
| Длина бокового сегмента листа, см (×11) Lateral leaf segment length, cm (×11) |
4.90 ± 0.2 | 4.66 ± 0.19 | 5.45 ± 0.18 | 4.11 ± 0.11 | 4.95 ± 0.16 | 4.67 ± 0.19 | 3.91 ± 0.14 | 4.47 ± 0.18 | 4.88 ± 0.26 | 3.36 ± 0.11 |
| Глубина выреза на боковом сегменте, см (×14) Depth of lateral segment cut, cm (×14) |
1.2 ± 0.19 | 0.5 ± 0.1 | 1 ± 0.15 | 1.05 ± 0.12 | 0.79 ± 0.11 | 1.15 ± 0.17 | 0.9 ± 0.11 | 0.84 ± 0.11 | 0.55 ± 0.11 | 0.55 ± 0.08 |
| Степень расчленения бокового сегмента листа (×15) Degree of lateral leaf segment incision (×15) |
0.25 ± 0.03 | 0.10 ± 0.02 | 0.18 ± 0.03 | 0.25 ± 0.03 | 0.16 ± 0.02 | 0.24 ± 0.03 | 0.23 ± 0.03 | 0.19 ± 0.02 | 0.11 ± 0.02 | 0.17 ± 0.03 |
| Длина лепестков, см (×16) Petals length, cm (×16) |
0.91 ± 0.03 | 0.73 ± 0.04 | 0.74 ± 0.02 | 0.83 ± 0.05 | – | 0.78 ± 0.04 | 0.69 ± 0.03 | 0.94 ± 0.03 | 0.94 ± 0.04 | 0.68 ± 0.02 |
Высокие показатели реальной семенной продуктивности A. jenisseensis отмечены для ЦП Aj5 (пойменный ивняк широкотравно-страусниковый, Западный Саян), где количество завязавшихся плодов составляет 27.4 шт., а коэффициент семенификации равен 74.6% (табл. 3). Для ЦП Aj10*, произрастающей на вырубке под ЛЭП, сравнительно низкие значения морфологических параметров сочетаются с относительно высокими показателями семенной продуктивности, численности и средним уровнем внутрипопуляционной изменчивости, что свидетельствует о длительно действующих однотипных стрессовых условиях (табл. 2, 3, рис. 2). Минимальные показатели семенной продуктивности (18.9 плодов на генеративный побег), отмечены для ЦП Aj6 (березово-сосновый лес папоротниково-широкотравный, Западный Саян).
Таблица 3.
Среднепопуляционные значения показателей семенной продуктивности Anemone jenisseensis Table 3. Average population values of seed productivity of Anemone jenisseensis
| № ц/п Coenopopula-tion number |
Реальная семенная продуктивность Actual seed productivity |
Потенциальная семенная продуктивность Potential seed productivity |
Коэффициент
семенификации Seed productivity coefficient |
|---|---|---|---|
| Aj1 | 10.40 ± 3.52 | 26.47 ± 2.30 | 39.88 ± 14.06 |
| Aj2 | 17.30 ± 1.57 | 28.37 ± 2.42 | 66.46 ± 4.09 |
| Aj3 | 0 | 20.79 ± 2.12 | 0 |
| Aj4* | 17.36 ± 2.40 | 27.70 ± 2.07 | 56.84 ± 4.18 |
| Aj5 | 27.43 ± 2.13 | 37.73 ± 2.56 | 74.60 ± 3.53 |
| Aj6 | 14.54 ± 1.77 | 18.90 ± 1.55 | 64.91 ± 5.22 |
| Aj7 | 16.45 ± 1.99 | 23.83 ± 2.24 | 68.79 ± 4.71 |
| Aj8 | 17.60 ± 3.36 | 26.70 ± 2.42 | 47.37 ± 7.98 |
| Aj9 | 14.69 ± 3.07 | 25.34 ± 2.51 | 47.02 ± 6.83 |
| Aj10* | 17.38 ± 1.78 | 27.17 ± 2.46 | 61.47 ± 4.90 |
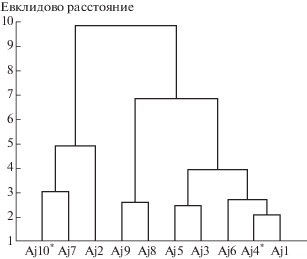
На дендрограмме сходства, построенной с помощью кластерного анализа, наблюдается разделение совокупности ЦП на 2 кластера (рис. 4). В первый кластер объединились подтаежные западносаянские ЦП Aj7 и Aj2, а также примкнувшая к ним Aj10*, произрастающая на вырубке под ЛЭП. Для особей данных ЦП отмечаются небольшие размеры листьев и слабое расчленение боковых сегментов. Второй кластер формируют два подкластера. Первый объединяет красноярские ЦП Aj8 и Aj9 с относительно крупным центральным сегментом листа. Боковой сегмент средних размеров, слабо расчленен. Второй подкластер образуют западносаянские ЦП, особи которых характеризуются крупными размерами вегетативных органов. Внутри данного подкластера наблюдается разделение на две группы по размерам и степени расчленения бокового сегмента листа. Для особей группы ЦП Aj1, Aj4* и Aj6 отмечаются крупные и глубоко расчлененные боковые сегменты.
Рис. 4.
Дендрограмма сходства ценопопуляций Anemone jenisseensis по морфологическим признакам. Ось Y – Евклидово расстояние; ось X – ценопопуляции.
Fig. 4. Dendrogram of similarity of Anemone jenisseensis coenopopulations by morphological characters. Y-axis – Euclidean distance; X-axis – coenopopulations.
Генетический полиморфизм ветреницы енисейской изучен на примере ЦП, произрастающих в лесах Западного Саяна (Aj1, Aj3, Aj5-Aj7), Восточного Саяна (Aj8) и Красноярской лесостепи (Aj9). В ходе анализа выявлено 111 фрагментов ДНК. Суммарный процент полиморфизма составляет 100. Число амплифицированных фрагментов ДНК, в зависимости от праймера (табл. 4), варьировало от 13 (HB8) до 24 (НВ12).
Таблица 4.
Полиморфизм межмикросателлитных фрагментов ДНК, амплифицированных ISSR-PCR методом Table 4. Polymorphism of intermicrosatellite DNA fragments amplified by ISSR-PCR method
| Последова-тельность Sequence |
Общее число фрагментов Total number of fragments |
Число полиморфных фрагментов ДНК (их частота) Number of polymorphic DNA fragments (frequency) |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Всего Total |
Полиморфных Polymorphic |
Aj1 | Aj3 | Aj5 | Aj6 | Aj7 | Aj8 | Aj9 | ||
| ISSR-праймер ISSR primer |
(5' → 3') | |||||||||
| 844B | (CT)8GC | 14 | 14 (100%) | 10 | 13 | 10 | 6 | 5 | 11 | 9 |
| 17898А | (CA)6АС | 19 | 19 (100%) | 13 | 15 | 13 | 10 | 13 | 9 | 12 |
| 17898В | (CA)6GT | 23 | 23 (100%) | 16 | 16 | 16 | 16 | 12 | 13 | 17 |
| 17899B | (CA)6GG | 18 | 17 (94.4%) | 9 | 11 | 7 | 9 | 12 | 10 | 12 |
| HB8 | (GA)6GG | 13 | 13 (100%) | 8 | 3 | 8 | 7 | 3 | 8 | 9 |
| HB12 | (CAС)3GС | 24 | 24 (100%) | 15 | 21 | 16 | 15 | 17 | 21 | 19 |
| Всего Total |
111 | 111 (100%) | 71 (63.96%) | 79 (71.17%) | 70 (63.06%) | 63 (56.76%) | 62 (55.86%) | 69 (62.16%) | 78 (70.27%) | |
Уровень выявляемого внутрипопуляционного генетического разнообразия средний (55.86–71.17%). Показатели генетического полиморфизма максимальны в западносаянской ЦП Aj3 (P = = 71.17%; I = 0.275; h = 0.404) и красноярской Aj9 (P = 70.27%; I = 0.249; h = 0.373). Средними показателями генетического разнообразия характеризуются западносаянские ЦП Aj1 (P = 63.93%; I = 0.209; h = 0.32) и Aj5 (P = 63.06%; I = 0.204; h = 0.313). Минимальные значения отмечены для Aj7 (P = 55.86%; I = 0.189, h = 0.287).
Коэффициент подразделенности ЦП (Gst) составляет 0.3316. Следовательно, на долю межпопуляционного разнообразия приходится 33.16%, а изученные ЦП ветреницы демонстрируют очень высокую степень дифференциации (Wright, 1978). Генетические дистанции М. Нея (Nei, 1972) минимальны между западносаянскими Aj7 и Aj3 (D = 0.0892). Максимальные расстояния зафиксированы между ЦП Aj6 и Aj7 (D = = 0.2788) (табл. 5).
Таблица 5.
Генетические дистанции Нея (1972) между ценопопуляциями Anemone jenisseensis на основе ISSR-PCR Table 5. Nei’s genetic distances between Anemone jenisseensis coenopopulation based on ISSR-PCR
| № ц/п Coenopopulation number |
Aj1 | Aj3 | Aj5 | Aj6 | Aj7 | Aj8 |
|---|---|---|---|---|---|---|
| Aj3 | 0.2269 | |||||
| Aj5 | 0.2024 | 0.1026 | ||||
| Aj6 | 0.1068 | 0.2390 | 0.2397 | |||
| Aj7 | 0.2499 | 0.0892 | 0.1452 | 0.2788 | ||
| Aj8 | 0.2286 | 0.1154 | 0.0963 | 0.2494 | 0.1742 | |
| Aj9 | 0.2113 | 0.1747 | 0.1184 | 0.2630 | 0.1901 | 0.1112 |
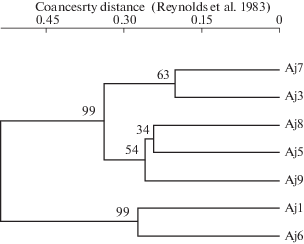
На UPGMA-дендрограмме сходства ЦП с высокой степенью достоверности выделяются два кластера (рис. 5). Первый кластер делится на два подкластера. В первый входят подтаежные западносаянские ЦП Aj3 и Aj7, во второй – красноярские Aj8 и Aj9 и западносаянская Aj5. Второй кластер образуют популяции Aj1 и Aj6, произрастающие в сосняках Западного Саяна и характеризующиеся крупными рассеченными боковыми долями листа. Данные ЦП отличались от других и по морфологическим признакам (рис. 4).
Рис. 5.
Дендрограмма сходства ценопопуляций Anemone jenisseensis на основе данных ISSR-PCR анализа (цифрами указаны значения бутстрепа в %).
Fig. 5. Dendrogram of similarity of Anemone jenisseensis coenopopulations based on ISSR-PCR analysis data (the numbers indicate bootstrap values, %).
ЗАКЛЮЧЕНИЕ
Проведенный анализ состояния ЦП ветреницы енисейской показал, что изучаемый вид обладает относительно широким фитоценотическим ареалом и может встречаться в мелколиственных (березовые, осиновые леса), светлохвойных (сосняки), смешанных лесах, в пойменных ивняках, а также на вырубках.
Максимальные размеры вегетативных органов, показатели семенной продуктивности, генетического разнообразия зафиксированы для ЦП A. jenisseensis, произрастающих в лесах Западного Саяна (Aj3, Aj5) и Красноярской лесостепи (Aj9). На вырубке под ЛЭП (Aj10*) низкие значения морфологических параметров сочетаются с относительно высокими показателями семенной продуктивности, численности и средним уровнем изменчивости, что свидетельствует о длительно действующих стрессовых условиях.
ЦП A. jenisseensis демонстрируют очень высокую степень генетической дифференциации (Gst = 0.3316). Важным диагностическим признаком, позволяющим разделить ЦП ветреницы енисейской, является степень расчленения сегментов листа. ЦП Aj1 и Aj6, характеризующиеся крупными расчлененными боковыми долями листа, формируют морфологически и генетически обособленную группу. Четкого разделения ценопопуляций по географическому принципу не выявлено.
Список литературы
[Barykina] Барыкина Р.П. 1995. Поливариантность способов естественного вегетативного размножения и расселения в семействе Ranunculaceae – Бюлл. МОИП. Отд. биол. 100 (1): 53–64.
Cai Y., Li S.-W., LiuY., Quan S., Chen M., Xie Y.-F., Jiang H.-Z., Wei E.-Z., Yin N.-W., Wang L., Zhang R., Huang C.-L, He X.-H., Jiang M.-F. 2009. Molecular phylogeny of Ranunculaceae based on internal transcribed spacer sequences. – Afr. J. Biotechnol. 8 (20): 5215–5224.
[Dyuryagina, Ivanova] Дюрягина Г.П., Иванова М.М. 1985. Характеристика ценопопуляций редких видов флоры Бурятии. – Бот. журн. 70 (11): 1529–1538.
[Filippov, Andronova] Филиппов Е.Г., Андронова Е.В. 2017. Генетическая структура популяций и естественная гибридизация Dactylorhiza salina и D. incarnata (Orchidaceae). – Генетика. 53 (3): 310–323.
Jiang N., Zhou Z., Yang J.B., Zhang S.D., Guan K.Y., Tan Y.H., Yu W.B. 2017. Phylogenetic reassessment of tribe Anemoneae (Ranunculaceae): Non-monophyly of Anemone s.l. revealed by plastid datasets. – PLoS ONE. 12 (3): e0174792. https://doi.org/10.1371/journal.pone.0174792
[Luferov, Starodubtsev] Луферов А.Н., Стародубцев В.Н. 1995 Семейство лютиковые (Ranunculaceae). – В кн.: Сосудистые растения Советского Дальнего Востока. Т. 7. СПб. С. 9–145.
[Mamaev] Мамаев С.А. 1972. Формы внутривидовой изменчивости древесных растений. М. 284 с.
[Marinicheva, Yamskikh] Мариничева А.Н., Ямских И.Е. 2017. Полиморфизм популяций Anemone reflexa (Ranunculaceae) в горах Южной Сибири. – Вестник КрасГАУ. 7 (130): 126–133.
Mansour A., Ismail H.M., Ramadan M.F., Gyulai G. 2009. Variations in tomato (Lycopersicon esculentum) cultivars grown under heat stress. – Journal fur verbrauchersschutz und lebensmittelsicherheit. 1.4: 118–127.
Miller M.P. 1997. Tools for population genetic analyses (TFPGA) 1.3: A Windows program for the analysis of allozyme and molecular population genetic data.
Mlinarec J., Satovic Z., Malenica N., Ivancic-Bace I., Besendorfer V. 2012. Evolution of the tetraploid Anemone multifidi (2n = 32) and hexaploid A. baldensis (2n = 48) (Ranunculaceae) was accompanied by rDNA loci loss and intergenomic translocation: evidence for their common genome origin. – Ann. Bot. 110 (3): 703–712. https://doi.org/10.1093/aob/mcs128
Nei M. 1972. Genetic Distance between Populations. – Am. Nat. 106: 283–292.
Paterson I.D., Downie D.A., Hill M.P. 2009. Using molecular methods to determine the origin of weed populations of Pereskia aculeata in South Africa and its relevance to biological control. – Biol. Control. l.48: 84–91.
[Popov] Попов М.Г. 1957. Семейство лютиковые (Ranunculaceae). – В кн.: Флора Средней Сибири. Т. 1. М., Л. С. 250–274.
[Red…] Красная книга Иркутской области. 2010. Иркутск. 478 с.
[Schmidt] Шмидт В.М. 1984. Математические методы в ботанике. – Л. 288 с.
Schuettpelz E., Hoot. S.B., Samuel R., Ehrendorfer F. 2001. Multiple origins of Southern Hemosphere Anemone (Ranunculaceae) based on plastid and nuclear sequence data. – Plant Syst. Evol. 231: 143–151.
Sun M., Yin X., Shi F., Li L., Li M., Li L., Xiao H. 2012. Development of Eighteen Microsatellite Markers in Anemone amurensis (Ranunculaceae) and Cross-Amplification in Congeneric Species. – Int. J. Mol. Sci. 13: 4889–4895. https://doi.org/10.3390/ijms13044889
[Timokhina] Тимохина С.А. 1993. Семейство лютиковые (Ranunculaceae). – В кн.: Флора Сибири. Т. 6. Новосибирск. С. 145–149.
[Voronov] Воронов А.Г. 1973. Геоботаника. 384 с.
Ward J.H.Jr. 1963. Hierarchical Grouping to Optimize an Objective Function. – Journal of the American Statistical Association. 58: 236–244.
Wright S. 1978. Evolution and the Genetics of Populations: Variability within and among natural populations. – University of Chicago Press. 4: 590.
[Yamskikh et al.] Ямских И.Е., Куцев М.Г., Уварова О.В. 2011. Полиморфизм популяций Anemonoides altaica (C.A. Mey.) Holub в горах Южной Сибири на основе морфологических данных и ISSR-PCR анализа. – Turczaninowia. 14 (1): 8–16.
[Yuzepchuk] Юзепчук С.В. 1937. Семейство лютиковые (Ranunculaceae). – В кн.: Флора СССР. Т. 7. М., Л. С. 225–282.
Zhang Y., Hong Y., Ren C., Ming T., Hoot S.B., Yang Q.E. 2015. Palynology, cytology, and molecular systematics of Anemone section Begoniifolia (Ranunculaceae). – Plant Syst. Evol. 301 (1): 411–424. https://doi.org/10.1007/s00606-014-1082-0
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал