

Ботанический журнал, 2020, T. 105, № 7, стр. 697-704
ПОБЕГООБРАЗОВАНИЕ У SILENE TATARICA (CARYOPHYLLACEAE) НА ПЕСЧАНЫХ ОТМЕЛЯХ Р. ВЯТКИ (Г. КИРОВ)
Е. В. Лелекова 1, *, М. Н. Шаклеина 1, Н. П. Савиных 1
1 Вятский государственный университет
610000 г. Киров, ул. Московская, д. 36, Россия
* E-mail: LelekovaEV1980@mail.ru
Поступила в редакцию 27.06.2019
После доработки 05.04.2020
Принята к публикации 14.04.2020
Аннотация
С позиций модульной организации выполнен структурно-морфологический анализ побеговых систем Silene tatarica (L.) Pers. Выделены и описаны анизотропные монокарпические побеги возобновления с геофильными участками и без них, а также силлептические побеги и побеги с неполным циклом развития с разными по числу метамеров вегетативными участками. Основу побегового тела растения составляют монокарпические побеги с геофильными участками. У них охарактеризованы фазы развития (почки; геофильного побега; вегетативного ассимилирующего побега; бутонизации, цветения и плодоношения; вторичной деятельности) и структурно-функциональные зоны. Модель побегообразования S. tatarica определена как симподиальная длиннопобеговая. Жизненная форма – поликарпический моноцентрический вегетативно мало подвижный летнезеленый травянистый каудексово‑стержнекорневой многолетник с анизотропными удлиненными монокарпическими побегами, гемикриптофит/геофит. Основные адаптации к существованию этого растения на прибрежьях в условиях занесения песком и периодического затопления: большой резерв спящих почек, способных к ветвлению (однажды занятая территория при этом осваивается неоднократно; растения успешно произрастают в экстремальных условиях), а также длительная связь системы побегов с главным корнем.
В настоящее время прибрежные растения исследуются с различных позиций (Cronk, Fennessy, 2001; Bornette, Puijalon, 2011; Zhmylev et al., 2012; Wang et al., 2016; Bobrov, 2017; и др.). Silene tatarica в этом плане не исключение. Она произрастает по прирусловым валам, песчаным пляжам и берегам рек, пойменным гривам, иногда сухим лесным опушкам, склонам и у дорог. У смолевки татарской изучены эколого-ценотическая характеристика и продуктивность (Volodina et al., 2004; и др.), генетическая структура ценопопуляций (Aspi et al., 2003; Tero et al., 2003, 2005; и др.), стратегии выживания в прибрежных и рудеральных местообитаниях (Jäkäläniemi et al., 2004, 2006; и др.). В свете возрастающего интереса к использованию растений в качестве растительного сырья для получения новых лекарственных препаратов, пищевых добавок и косметических композиций, исследованы и биохимические свойства S. tatarica (Lafont et al., 1993; Chadin et al., 2004; Tero et al., 2006; Gyunter, 2016; и др.). В дополнение к изученному, анализ структурной организации особей S. tatarica, равно как и остальных прибрежно-водных растений, в различных местообитаниях позволит наметить и определить основные тенденции их морфологических адаптаций к существованию в условиях переменного увлажнения и обводнения (Savinykh et al., 2015; Savinykh, Shabalkina, 2017; и др.).
Цель работы: изучить биоморфологические особенности S. tatarica на песчаных отмелях для выявления структурно-функциональных адаптаций к условиям среды обитания.
МАТЕРИАЛ И МЕТОДЫ
Материал для исследования собран на песчаных затопляемых отмелях р. Вятка в окрестностях г. Кирова. Биоморфологический анализ побеговых систем S. tatarica проведен с учетом представлений о растении как целостном организме (Shafranova, Gattsuk, 1994). Жизненная форма растения охарактеризована с позиций синтетической множественной классификации с учетом подходов И.Г. Серебрякова (Serebryakov, 1949, 1954, 1964), О.В. Смирновой (Smirnova et al, 1976) и С. Raunkiaer (1934). Структура особей рассмотрена с позиций модульной организации с использованием трех категорий модулей: элементарного, универсального и основного (Savinykh, Maltseva, 2008).
Основные методы исследования – разработанный И.Г. Серебряковым (Serebryakov, 1952; 1964) на базе сравнительно-морфологического анализа способ описания конкретных структур; ритмологический (Serebryakov, 1949; 1954) и биоморфологический (Sovremennyye podkhody…, 2008). Строение монокарпических побегов и их структурно-функциональную зональность оценивали согласно подходам W. Troll (1964), И.В. Борисовой и Т.А. Поповой (Borisova, Popova, 1990); развитие их – в соответствии с представлениями Серебрякова (Serebryakov, 1954) и Т.И. Серебряковой (Serebryakova, 1971).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Жизненная форма S. tatarica – поликарпический моноцентрический вегетативно мало подвижный летнезеленый травянистый каудексово-стержнекорневой многолетник с анизотропными удлиненными монокарпическими побегами, гемикриптофит/геофит.
Тело растения формируется по симподиальной длиннопобеговой модели побегообразования и состоит из удлиненных моно- и дициклических анизотропных монокарпических побегов возобновления с геофильной частью и без нее, небольших силлептических побегов и побегов с неполным циклом развития – универсальных модулей побеговой системы этого растения. Монокарпические побеги возобновления с геофильным участком формируются из почек возобновления (а также спящих при повреждении надземной части особи) такого же побега предыдущего порядка ветвления. Побег первого порядка развивается из семени (рис. 1, 2).
Рис. 1.
Формирование побеговой системы Silene tatarica: особь семенного происхождения в первый год жизни (а – лето; b – осень); развитие особи во второй год жизни (с – лето; d – осень); 1 – уровень грунта текущего года; 2 – уровень грунта прошлого года; 3 – вегетативный побег; 4 – вегетативно-генеративный побег; 5 – узел с листьями и побегом; 6 – ветвящиеся почки; 7 – неветвящиеся почки; 8 – семя; 9 – главный корень; 10 – боковой корень; 11 – шнуровидные придаточные корни; 12 – тонкие ветвящиеся придаточные корни; 13 – монокарпический побег возобновления в фазу цветения, плодоношения и бутонизации; 14 – монокарпический побег возобновления в фазу вегетативного ассимилирующего побега; одинарной линией обозначены однолетние части побегов, двойной – многолетние, пунктирной – отмершие.
Fig. 1. Forming shoots of Silene tatarica: individual of seed origin in the first year of life (а – summer; b – autumn); plant development in the second year of life (с – summer; d – autumn); 1 – current ground level; 2 – ground level of the last year; 3 – vegetative shoot; 4 – vegetative-generative shoot; 5 – node with leaves and shoots; 6 – branching buds; 7 – unbranched buds; 8 – seed; 9 – main root; 10 – lateral root; 11 – cord-like adventitious roots; 12 – thin branched adventitious roots; 13 – monocarpic renewal shoot in the flowering, fruiting and budding phase; 14 – monocarpic renewal shoot in the vegetative assimilation shoot phase; simple line indicates annual areas of shoots, double line – perennial part of shoots, dotted line – dead shoots.

Рис. 2.
Формирование побеговой системы Silene tatarica на третий год развития особи: а – в условиях повышения уровня грунта в результате занесения песком при весеннем половодье; b – без значительного изменения уровня грунта.
Fig. 2. Forming shoots of Silene tatarica in the third year of life: а – under conditions of rising ground level as a result of the influx of sand during the spring flood; b – without significant change of ground level.
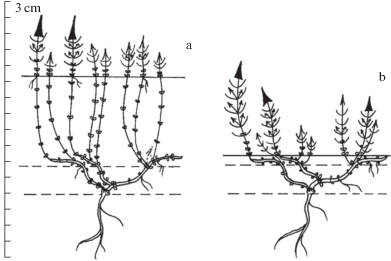
Главный корень сохраняется в течение всей жизни особи. На нем развиваются боковые слабоветвящиеся шнуровидные корни, диаметром 1–5 мм, которые располагаются зачастую параллельно поверхности почвы (рис. 3a). Наряду с питанием растения, они несут функцию закрепления особи, что особенно необходимо в условиях переменного обводнения. На геофильных участках некоторых побегов образуются слабоветвистые, длинные, шнуровидные придаточные корни, которые, как и боковые, питают и закрепляют растение. Кроме того, для побегов S. tatarica, располагающихся в поверхностном слое грунта и стелющихся по нему, характерно образование немногочисленных коротких тонких ветвящихся придаточных корней. За счет них происходит закрепление и дополнительное питание побегов, пространственно удаленных от главного корня (рис. 2).
Рис. 3.
Спящие почки на главах каудекса Silene tatarica: а – внешний вид многоглавого каудекса со спящими почками (линиями отмечены уровни грунта разных лет); b–d – ветвление спящих почек.
Fig. 3. Dormant buds on the heads of Silene tatarica caudex: а – multi-headed caudex with dormant buds (lines mark ground levels in different years); b–d – branching dormant buds.

В строении монокарпического побега S. tatarica выделяется несколько структурно-функциональных зон (рис. 4d), которые формируются в соответствующие фазы его развития.
Рис. 4.
Универсальные модули побеговой системы Silene tatarica: а–b – побеги возобновления без геофильных участков; c – силлептический побег; d – монокарпический побег с геофильным участком; 1 – главное соцветие; 2 – верхняя зона торможения; 3 – зона обогащения; 4 – средняя зона торможения; 5 – зона возобновления; 6 – нижняя зона торможения.
Fig. 4. Universal modules of the shoot system of Silene tatarica: а–b – renewal shoots without geophilic areas; c – sylleptic shoot; d – monocarpic renewal shoot with geophilic areas; 1 – main inflorescence; 2 – upper zone of inhibition; 3 – enrichment zone; 4 – middle inhibition zone; 5 – zone of renewal; 6 – lower inhibition zone.
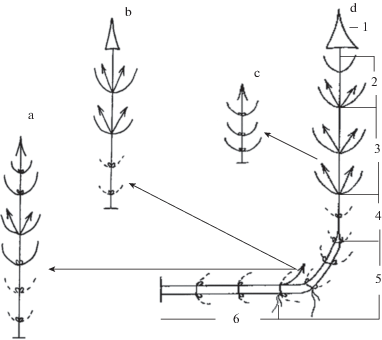
Фаза почки продолжается от года (почки возобновления) до трех и более лет (спящие почки). У одного монокарпического побега трогаются в рост 3–5 (редко более) почек возобновления в основании надземного участка побега. Остальные переходят в разряд спящих и сохраняют жизнеспособность в течение ряда лет. Спящие почки базальных метамеров геофильных участков побегов ветвятся (рис. 3b–d) подобно таковым у древесных растений, например, у березы (Serebryakov, 1952). Чем старше геофильный участок, тем более ветвится спящая почка: наибольшее число дочерних структур образуется у почек стеблевой части каудекса исходного побега (рис. 3b–c). На участках побегов, близко расположенных к поверхности грунта и над ним, почки, в большинстве своем, не ветвятся (рис. 2а, 3а). Спящие почки S. tatarica – это резерв возобновления особи.
Фаза геофильного побега непродолжительна: чаще до одного месяца (в начале вегетационного периода с момента реализации пазушной почки и выхода побега на поверхность грунта). Длина геофильного участка зависит от степени занесения песком исходного побега (рис. 2) и достигает 10–20 (40) см при толщине 0.2–0.3 мм. На верхушке располагается почка небольшой емкости, покрытая почечными чешуями. Эта часть монокарпического побега сложена однотипными элементарными модулями: все они с длинным междоузлием и рано отмирающими небольшими чешуевидными листьями, располагающимися супротивно. Различия этих модулей лишь в периоде покоя пазушных почек. Как правило, у большинства метамеров геофильного участка побега они остаются спящими. Последний соответствует нижней зоне торможения (рис. 4d).
Почки возобновления закладываются у метамеров приземного участка побега; их междоузлия значительно короче. Именно из них на следующий год будут развиваться побеги, обеспечивающие возобновление целостной особи. В случае занесения песком материнского побега при половодье в следующем году, вновь сформируются побеги с длинными геофильными участками. Если уровень грунта не изменится, из данных почек образуются монокарпические побеги без геофильных участков, которые своим основанием полегают на грунт. Целостная особь при этом приобретает вид небольшого куста. Этот участок побега принимает на себя функции зоны возобновления. У него сохраняются также функции поглощения и закрепления на поверхности субстрата за счет имеющихся придаточных корней.
Фаза вегетативного ассимилирующего побега составляет 2–3 месяца для моноциклических и 14–15 месяцев – для дициклических монокарпических побегов. Высота надземного участка монокарпического побега в среднем составляет 10–40 см. Главная его функция – ассимиляционная. Все его элементарные модули с длинными междоузлиями. В базальной части надземного участка монокарпического побега листья небольшие, ланцетные, рано отмирающие и разрушающиеся. Стебель одревесневающий, нередко с красновато-бурым оттенком. Пазушные почки могут реализовываться в побеги последующего порядка, стелющиеся по поверхности грунта или приподнимающиеся над ним. Этот участок также выполняет функции зоны возобновления. Следующие несколько метамеров, почки которых не развиваются, входят в состав средней зоны торможения. Их листья, как правило, также отмирают рано. Выше по стеблю расположены метамеры с ланцетными, острыми, сидячими, стеблеобъемлющими листьями. Из пазушных почек этого участка образуются небольшие вегетативные силлептические побеги, формирующие дополнительную ассимилирующую поверхность. Это зона обогащения монокарпического побега S. tatarica. Верхняя зона торможения включает несколько метамеров, почки которых не реализуются.
В конце осени надземные ортотропные участки этих побегов полегают. Листья срединной формации отмирают; верхушечная и пазушные (рис. 5) почки базальных метамеров остаются живыми. В следующем вегетационном периоде эти побеги, при успешной перезимовке, а также выходе на поверхность при занесении песком во время половодья, продолжат развитие и перейдут в фазу бутонизации, цветения и плодоношения. В некоторых случаях, на отдельных монокарпических побегах текущего года в конце осени можно наблюдать заложение соцветий, которые разовьются летом следующего года.
Рис. 5.
Пазушные почки в основаниях полегающих надземных участков монокарпических побегов Silene tatarica в конце вегетационного периода: а – часть почек реализовалась в вегетативные побеги; b – ветвление пазушных почек; c – незначительное увеличение почек в размерах.
Fig. 5. Axillary buds at the base of decumbent above-ground parts of monocarpic shoots of Silene tatarica in the end of the vegetation period: а – some buds have realized into vegetative shoots; b – branching of axillary buds; c – slight increase in buds size.

Образованием главного соцветия – кистевидной или пирамидальной однобокой метелки – завершается нарастание монокарпических побегов; не перешедшие в генеративную фазу, становятся побегами с неполным циклом развития.
Фаза вторичной деятельности монокарпического побега достаточно продолжительна. Апикальная и средняя части его отмирают. Базальный участок одревесневает и становится частью многоглавого каудекса со спящими почками. На нем можно различить своеобразные ежегодные “этажи” – утолщения стебля в месте формирования побегов следующего порядка ветвления. Их “высота” зависит от степени поднятия грунта в результате занесения песком и может достигать 40–50 см (рис. 3а).
Специализированная морфологическая дезинтеграция возможна, но дочерние особи мало жизнеспособны. Поэтому их нельзя считать вегетативными диаспорами, из которых развиваются полноценные особи вегетативного происхождения.
Ветвление S. tatarica обеспечивается деятельностью пазушных почек приземных участков побегов (рис. 5), а также спящих почек глав каудекса (рис. 3а).
При анализе побеговых систем выявлены каудексы с главами до восьмого порядка ветвления с жизнеспособными спящими почками. Так, в опыте по их реализации, мы изымали особи этого растения из естественной среды в конце вегетационного периода и удаляли большую часть побегов до их базальных участков. Каудексы помещали в грунт, который увлажняли по мере необходимости. При этом, большинство почек, проведших в состоянии покоя более пяти лет, тронулись в рост (рис. 6), а часть почек незначительно увеличилась в размерах. Эта особенность S. tatarica позволяет особям восстанавливаться после травматических повреждений.
Рис. 6.
Развитие побегов из спящих почек каудекса Silene tatarica: 1 – побеги возобновления; 2 – жизнеспособные спящие почки.
Fig. 6. Development of shoots from sleeping buds of Silene tatarica caudex: 1 – shoots of renewal; 2 – viable dormant buds.

Таким образом, в результате образования монокарпических побегов и повторного освоения занятой территории в случае повреждения особи, поддерживается центр закрепления, подобный первичному кусту многолетних наземных трав. Надземные участки побегов разных порядков ветвления отмирают; базальные их участки становятся главами ветвящегося каудекса – основного модуля побеговой системы S. tatarica.
ЗАКЛЮЧЕНИЕ
Основу побегового тела S. tatarica составляют: анизотропные моно- и дициклические монокарпические побеги с геофильными участками и без них, а также силлептические побеги и побеги с неполным циклом развития с разными по числу метамеров вегетативными участками. Побеги всех типов – универсальные модули (рис. 4). Поддержание ценопопуляций, в основном, семенное.
Основные адаптации к существованию в условиях переменного обводнения, регулярного занесения песком и периодического полного затопления особей следующие:
1. Большой резерв жизнеспособных спящих почек, способных к ветвлению. Однажды занятая территория при таком побегообразовании может осваиваться неоднократно, обеспечивая успешное существование особей в экстремальных условиях прибрежий и пляжей. При значительном повреждении особей, возобновление осуществляется за счет спящих почек каудекса, расположенных зачастую глубоко в грунте.
Генеративные побеги в конце вегетационного периода отмирают до основания надземных одревесневающих участков. Почки на них сохраняются живыми и обеспечивают возобновление особей. Это сближает S. tatarica с полукустарничками. Сочетание признаков полудревесных и травянистых растений у S. tatarica также является адаптацией к существованию в меняющихся условиях динамичного песчаного почвогрунта.
2. Длительная связь системы побегов с главным корнем. Геофильные участки побегов последовательных порядков становятся главами ветвящегося каудекса. Побеги, находящиеся на поверхности грунта, порой значительно удалены от главного корня, но остаются жизнеспособными и переходят в вегетативно-генеративную фазу, получая достаточно влаги именно за счет его деятельности.
Список литературы
Aspi J., Jakalaniemi A., Tuomi J., Siikamaki P. 2003. Multilevel phenotypic selection on morphological characters in a metapopulation of Silene tatarica. – Evolution. 57 (3): 509–517.
[Bobrov] Бобров Ю.А. 2017. Жизненные формы водных трав Северо-Востока Европейской России. – Arctic Environmental Research. 17 (2): 104–112.
[Borisova, Popova] Борисова И.В., Попова Т.А. 1990. Разнообразие функционально-зональной структуры побегов многолетних трав. – Бот. журн. 75 (10): 1420–1425.
Bornette G., Puijalon S. 2011. Response of aquatic plants to abiotic factors: a review. – Aquatic Sciences. 73: 1–14.
[Chadin et. al.] Чадин И.Ф., Володина С.О., Володин В.В. 2004. Индивидуальная изменчивость растений смолевки татарской по уровню накопления 20-гидроксиэкдизона. – Вестник Института биологии Коми научного центра Уральского отделения РАН. 1 (75): 13–15.
Cronk J.K., Fennessy M.S. 2001. Wetland plants: biology and ecology. London, N.Y., Washington. 462 p.
Gyunter E.A. 2016. Pectinic substances from Silene tatarica. Callus culture and native plant. – Chemistry of Natural Compounds. 52 (3): 458–462.
Jäkäläniemi A., Kauppi A., Pramila A., Vähätaini K. 2004. Survival strategies of Silene tatarica (Caryophyllaceae) in riparian and ruderal habitats. – Can. J. Bot. 82: 491–502.
Jäkäläniemi A., Tuomi J., Siikamaki P. 2006. Conservation of Species in Dynamic Landscapes: Divergent Fates of Silene tatarica Populations in Riparian Habitats. – Conserv. Biol. 20 (3): 844–852.
Lafont R., Parter C.J., Williams E., Read H., Morgan E.D., Wilson J.D. 1993. The application of off-line HPTLC-MS-MS to the identification of ecdysteroids in plant and arthropod samples. – Planar Chromatogr.-Mod. TLC. 6 (5): 421–424.
Raunkiaer C. 1934. The life form of plants and statistical plant geography. Oxford. 632 p.
[Savinykh, Maltseva] Савиных Н.П., Мальцева Т.А. 2008. Модуль у растений как структура и категория. – Вестник Тверского государственного университета. Сер. Биология и экология. 9: 227–234.
[Savinykh et al.] Савиных Н.П., Шабалкина С.В., Лелекова Е.В. 2015. Биоморфологические адаптации гелофитов. – Сибирский экологический журнал. 22 (5): 671–681.
[Savinykh, Shabalkina] Савиных Н.П., Шабалкина С.В. 2017. Основные преадаптации цветковых растений к вторичному заселению водоемов. – В сб.: Биоморфологические исследования на современном этапе: матер. конф. с междунар. участием “Современные проблемы биоморфологии”. Владивосток. С. 164–166.
[Serebryakov] Серебряков И.Г. 1949. Структура и ритм в жизни цветковых растений. Ч. 2. – Бюлл. МОИП. Отд. биол. 54 (2): 47–62.
[Serebryakov] Серебряков И.Г. 1952. Морфология вегетативных органов высших растений. М. 390 с.
[Serebryakov] Серебряков И.Г. 1954. О методах изучения ритмики сезонного развития растений в геоботанических стационарах. – В кн.: Докл. совещ. по стационарным геобот. исслед. Л. С. 145–159.
[Serebryakov] Серебряков И.Г. 1964. Жизненные формы высших растений и их изучение. – Полевая геоботаника. 3: 148–208.
[Serebryakova] Серебрякова Т.И. 1971. Морфогенез побегов и эволюция жизненных форм злаков. М. 360 с.
[Smirnova et al.] Смирнова О.В., Заугольнова Л.Б., Торопова Н.А., Фаликов Л.Д. 1976. Критерии выделения возрастных состояний и особенности хода онтогенеза у растений различных биоморф. – В кн.: Ценопопуляции растений: Основные понятия и структура. М. С. 14–43.
[Shafranova, Gattsuk] Шафранова Л.М., Гатцук Л.Е. 1994. Растение как пространственно временная метамерная (модульная) система. – В сб.: Успехи экологической морфологии и ее влияние на смежные науки: Межвузовский сб. научных трудов. М. С. 6–7.
[Sovremennyye podkhody…] Современные подходы к описанию структуры растений. 2008. Киров. 355 с.
Tero N., Aspi J., Siikamaki P., Jakalaniemi A., Tuomi J. 2003. Genetic structure and gene flow in a metapopulation of an endangered plant species Silene tatarica. – Mol. Ecol. 12: 2073–2085.
Tero N., Aspi J., Siikamäki P., Jäkäläniemi A. 2005. Local genetic population structure in an endangered plant species Silene tatarica (Caryophyllaceae). – Heredity. 94: 1–10.
Tero N., Neumeier H., Gudavalli R., Schlötterer C. 2006. Silene tatarica microsatellites are frequently located in repetitive DNA. – Journal of evolutionary biology. 19: 1612–1619.
Troll W. 1964. Die Infloreszenzen. Jena. 615 s.
[Volodina et. al.] Володина С.О., Чадин И.Ф., Мартыненко В.А., Володин В.В. 2004. Эколого-ценотическая характеристика и продуктивность надземной части смолевки татарской в природных популяциях (Европейский северо-восток России). – Вестник института биологии Коми научного центра Уральского отделения РАН. 6 (80): 2–5.
Wang H., Wang Q., Bowler P, Xiong W. 2016. Invasive aquatic plants in China. – Aquatic Invasions. 11 (1): 1–9.
[Zhmylev et al.] Жмылёв П.Ю., Леднёв С.А., Щербаков А.В. 2012. Биоморфология водных растений: проблемы и подходы к классификации жизненных форм. – В сб.: Леонид Васильевич Кудряшов. Ad Memoriam: Сб. статей. М. С. 101–128.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал