

Ботанический журнал, 2020, T. 105, № 7, стр. 687-696
ХАРАКТЕРИСТИКА ПОПУЛЯЦИИ THYMUS REVERDATTOANUS (LAMIACEAE) НА ЮГЕ АРЕАЛА (НАДЫМСКИЙ РАЙОН, ЯМАЛО-НЕНЕЦКИЙ АВТОНОМНЫЙ ОКРУГ)
Н. Н. Лащинский 1, Е. Б. Таловская 1, *
1 Центральный сибирский ботанический сад СО РАН
630090 г. Новосибирск, ул. Золотодолинская, 101, Россия
* E-mail: kolegova_e@mail.ru
Поступила в редакцию 07.09.2019
После доработки 05.03.2020
Принята к публикации 17.03.2020
Аннотация
Приводится новое местонахождение Thymus reverdattoanus – редкого вида на территории Ямало-Ненецкого автономного округа. Анализируются особенности морфологии особей и эколого-фитоценотические условия нового местообитания. Небольшая популяция обнаружена в подзоне северной тайги Западно-Сибирской равнины на песках в долине реки. Описаны растительные сообщества, в которых обитает тимьян и проанализирована структура генеративных особей. По сравнению с основной частью ареала выявленные местообитания отличаются временным характером в активной пойме реки. Высказаны предположения о происхождении этой изолированной популяции и о возможных путях и способах миграции вида.
Важным направлением при изучении биоразнообразия является выявление характера распространения и условий местообитаний эндемичных видов растений. Они представляют большой интерес для объяснения происхождения, особенностей и путей миграции, как отдельных таксонов, так и флоры конкретного региона в целом. Немаловажное значение имеют сведения об эндемиках и для организации природоохранной деятельности (Tolmachov, 1974; Bykov, 1979; Endemichnye…, 2013; Brodskiy, 2016).
Одним из замечательных эндемиков Сибири является Thymus reverdattoanus Serg. (Lamiaceae). Сибирский гипоарктический вид, распространенный в Западной (Тюменская область) и Средней Сибири (Иркутская область, Красноярский край), Якутии (Doron’kin, 1997). На территории Ямало-Ненецкого автономного округа T. reverdattoanus является редким, внесен в Красную книгу ЯНАО (Krasnaya…, 2010).
Согласно литературным данным T. reverdattoanus11 обычен в сообществах тундровой зоны по южным склонам песчаных холмов и террас, развеянным пескам. В лесном поясе и высокогорьях вид участвует в сообществах, расположенных на щебнистых и песчаных склонах гор и скалах. В полосе редколесий встречается спорадично на сухих пойменных лугах и по щебнистым южным остепненным склонам (Menitskii, Yurtsev, 1980; Kuvaev, 2006; Pospelova, Pospelov, 2007). Согласно высотному распределению видов во флоре субарктических гор И.Б. Куваева (Kuvaev, 2006), T. reverdattoanus встречается в диапазоне от 200 до 1100 м над ур. м. Все местообитания вида приурочены к участкам с глубоким протаиванием мерзлоты в летнее время. Во флорах и Красных книгах для вида указывается жизненная форма полукустарничка (Klokov, 1954; Krasnaya…, 2010). Как отмечает Е.Е. Гогина (Gogina, 1990), в тундровой и лесной зонах популяции тимьянов интразональны и представляют собой вкрапления, приуроченные к специфическим условиям. Численность популяций T. reverdattoanus на протяжении ареала невелика, особи располагаются единично и небольшими группами, образующими пятна (Krasnaya…, 2010).
При проведении экспедиционных работ по описанию растительного покрова Западно-Сибирской равнины популяция T. reverdattoanus была обнаружена в окрестностях п. Ягельный Надымского р-на Ямало-Ненецкого автономного округа значительно южнее от известных местонахождений вида на данной территории.
МАТЕРИАЛЫ И МЕТОДЫ
Определение видовой принадлежности собранного нами тимьяна проведено с учетом консультации д-ра биол. наук М. C. Князева и типовых образцов T. reverdattoanus (LE, TK). Снимки гербарных образцов выполнены с использованием сканера ObjectScan 1600 (Microtek, Тайвань) и стереомикроскопа Carl Zeiss SteREO Discovery.V12 с камерой AxioCam HRc (Германия).
Эколого-ценотическая характеристика дана на основе десяти оригинальных геоботанических описаний, выполненных по стандартной методике на пробных площадях размером 20×20 м для лесных сообществ и 10×10 м для травянистых, с географической привязкой по 12-канальному GPS в системе координат WGS-84. При описании лесных сообществ эпифитные мхи и лишайники не учитывались. Все геоботанические описания внесены в единую базу данных с использованием программного пакета IBIS 7.2 (Zverev, 2007). Оценка проективного покрытия видов в таблице приводится по шкале Браун-Бланке (Becking, 1957): r – единично; + – менее 1%; 1 – 1–5%; 2 – 5–25%; 3 – 25–50%; 4 – 50–75%; 5 – 75–100%. Постоянство видов дано в классах от I до V с величиной класса в 20%. Гербарный материал T. reverdattoanus для определения и морфологического анализа в количестве 5 гербарных листов отбирался из разных локусов популяции.
Определение жизненной формы T. reverdattoanus проведено с использованием эколого-морфологической классификации жизненных форм И.Г. Серебрякова (Serebryakov, 1964). Тип биоморфы установлен в соответствии с фитоценотической классификацией О.В. Смирновой (Smirnova, 1987), построенной на особенностях пространственного распределения побегов, почек возобновления и корней растения. При характеристике побеговой системы мы опирались на классификацию М.Т. Мазуренко и А.П. Хохрякова (Mazurenko, Khokhryakov, 1977). Были выделены побеги: обогащения, ветвления, составная скелетная ось. В работе использованы термины: безрозеточный побег (Serebryakov, 1959); верхнерозеточный побег (Nukhimovskii, 1997); среднерозеточный побег (Bobrov, 2009). Для сравнения особенностей побеговой структуры был проанализирован гербарный материал из типичных мест обитания T. reverdattoanus (LE, TK).
РЕЗУЛЬТАТЫ
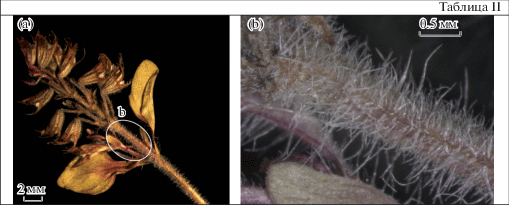
Несмотря на установленную видовую принадлежность собранных нами образцов, хотелось бы отметить некоторые морфологические отличия наших экземпляров от типовых: удлиненные ортотропные побеги, голые листовые пластинки, разнообразное опушение побегов под соцветием (из длинных волосков или из длинных и коротких) (табл. I, IIa, b). Указанные различия могут объясняться спецификой экологии местообитаний, а также отражать высокую модификационную изменчивость популяций вида, присущую всем тимьянам арктического распространения (Gogina, 1990).
Таблица I. Гербарный образец T. reverdattoanus.
Plate I . Herbarium specimen of Thymus reverdattoanus.

Таблица II. Опушение побега под соцветием.
Plate II . Pubescence of the shoot under the inflorescence.
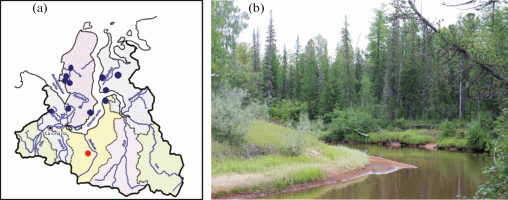
Обнаруженная популяция T. reverdattoanus является на данный момент крайней южной точкой распространения вида на территории Ямало-Ненецкого автономного округа , отстоящей от ранее известных местонахождений на 350 км к югу (рис. 1a). Ценопопуляции тимьяна были обнаружены в северной подзоне таежной зоны равнинной части Западной Сибири в долине небольшой реки в окрестностях п. Ягельный Надымского р-на Ямало-Ненецкого автономного округа (рис. 1b). Территория с поверхности перекрыта мощным чехлом рыхлых четвертичных отложений, верхняя часть которых представлена аллювиальными песками. Зональные почвы – иллювиально-железистые подзолы. Территория расположена в области сплошного распространения многолетнемерзлых грунтов. Мощность активного слоя в конце августа составила 1.7 м. Фоновая растительность по долине реки представлена северо-таежными кедрово-лиственничными бруснично-зеленомошными лесами (табл. 1, оп 1–6). Древостой сомкнутостью 0.5–0.6 отчетливо делится на два подъяруса. Первый подъярус высотой 24–28 м образован лиственницей (Larix sibirica Ledeb.) со значительной (до 4–5 единиц состава) примесью кедра (Pinus sibirica Du Tour). Стволы ровные, диаметром 26–32 см на высоте 1.3 м; очищение от сучьев хорошее, живая крона начинается на высоте 6–8 м от поверхности почвы. Второй подъярус сомкнутостью 0.4–0.5 сформирован преимущественно елью (Picea obovata Ledeb.) при единичном участии лиственницы и кедра. Высота второго яруса 14–18 м при диаметре стволов 12–14 см. Распределение деревьев случайное. Встречается единичный подрост березы (Betula pubescens Ehrh.), ели и кедра. В подлеске единичные разновысокие кусты ольховника (Duschekia fruticosa (Rupr.) Pouzar), можжевельника (Juniperus communis L.) и рябины (Sorbus sibirica Hedl.). Травяно-кустарничковый ярус 25 см высотой равномерный с отчетливым доминированием брусники (Vaccinium vitis-idaea L.) образован мелкими таежными и таежно-болотными травами и кустарничками; общее проективное покрытие составляет 75–90%. Моховой покров сплошной из типичных бореально-таежных мхов (табл. 1). Видовое разнообразие составляет 12–14 видов сосудистых растений на 400 м2. В рамках эколого-флористической классификации эти леса относятся к порядку Ledo palustris-Laricetalia cajanderi Ermakov in Ermakov et Alsynbayev 2004 класса Vaccinio-Piceetea Br.-Bl. in Br.-Bl. et al. 1939.
Рис. 1.
Распространение (a) и изученное местообитание (b) Thymus reverdattoanus в Ямало-Ненецком автономном округе. Синими точками обозначены известные местонахождения вида (Krasnaya…, 2010), красной – новое местонахождение.
Fig. 1. Distribution (a) and habitat (b) of Thymus reverdattoanus in Yamal-Nenets Autonomous Area. Blue dots indicate known localities of the species (Krasnaya…, 2010), red dot indicate a new locality.
Сообщества с участием T. reverdattoanus представлены в виде небольших обособленных участков площадью не более 200–300 м2 каждый, расположенных на аккумулятивных частях высокой поймы, примыкающих к первой надпойменной террасе, покрытой сомкнутым лесом. Расстояние между соседними сообществами составляет 150–200 м. Субстрат образован свежими песчаными аллювиальными наносами. По структуре и видовому составу сообщества резко контрастируют с зональными лесами (табл. 1, оп 7–10). Это открытые травяные сообщества с доминированием длиннокорневищного злака Calamagrostis epigeios (L.) Roth. Общее проективное покрытие 55–75%. Травостой 45–55 см высотой с хорошо выраженной злаковой основой при единичном участии таежных кустарничков (табл. 1 ). В составе травостоя преобладают виды опушечных (Calamagrostis epigeios) и пойменных (T. bipinnatum (L.) Sch. Bip.) местообитаний. В моховом покрове редкие дернинки пионерных видов, характерных для песчаных субстратов (P. juniperinum Hedw. и P. piliferum Hedw.). В рамках эколого-флористической классификации данные сообщества могут рассматриваться в составе класса Epilobietea angustifolii Tüxen et Preising ex von Rochow 1951 – пионерных сообществ начальных стадий посткатастрофических сукцессий. Проективное покрытие T. reverdattoanus меняется от единичного участия до 10–15%. На момент сбора (28 августа) растения тимьяна были в фазе окончания цветения. В ценопопуляции T. reverdattoanus выделяются локусы, образованные группой из нескольких взрослых особей, вблизи и под пологом которых развивается подрост.
Для T. reverdattoanus в этих условиях характерна жизненная форма моноцентрического полукустарничка. Взрослое растение представляет собой куст с полегающими и слабо укореняющимися составными скелетными осями (рис. 2). Многолетнюю основу куста образуют побеги, которые отличаются по положению в пространстве и роли. Основное значение среди них имеют восходящие и ортотропные вегетативные побеги ветвления, которые развиваются из зимующих почек. Они моноподиально нарастают два года, каждый их годичный прирост по структуре верхнерозеточный. В зимний период отмирает большая часть побега ветвления, сохраняются 2–4 базальных метамера первого годичного прироста с почками возобновления. В дальнейшем, в результате базисимподиального нарастания формируется составная скелетная ось, состоящая из последовательно сменяющих друг друга базальных частей побегов ветвления. Составная скелетная ось может быть ортотропной (состоит из ортотропных базальных частей побегов ветвления) или восходящей (состоит из плагиотропных базальных частей побегов ветвления). Первая принимает участие в увеличении запаса почек возобновления, вторая способствует разрастанию куста. В структуре особей T. reverdattoanus выявлено разнообразие генеративных побегов. Выделены двулетние генеративные побеги, по структуре они среднерозеточные восходящие (их вегетативный прирост плагиотропный верхнерозеточный, а годичный генеративный прирост ортотропный безрозеточный), развиваются из зимующих почек. Для них характерна синфлоресценция, которая образована главным соцветием и 2–3 паракладиями первого порядка. Соцветие, заканчивающее главный побег, и соцветие паракладий – колосовидный тирс, состоящий из супротивно расположенных дихазиев, нередко с одной отодвинутой парой. После плодоношения большая часть генеративного побега отмирает, сохраняется базальная часть с почками возобновления, которая также, как у двулетних вегетативных побегов, принимает участие в образовании многолетней основы куста. Однолетние генеративные побеги развиваются из зимующих и силлептических почек. По структуре однолетние генеративные побеги ортотропные безрозеточные. Они не ветвятся, соцветие представляет собой головчатый тирс. После плодоношения, как правило, полностью отмирают. Однолетние генеративные побеги выполняют функцию обогащения куста ассимиляционными и репродуктивными органами и не участвуют в построении многолетней основы. Корневая система смешанная, образована системой главного и придаточных корней. Главный корень сохраняется до отмирания всей особи. Придаточные корни нитевидные, развиваются в узлах и на междоузлиях того участка побега, который соприкасается с субстратом. Преобладающий способ размножения – семенной. Вегетативное размножение слабое, происходит в основном в результате механического повреждения оси. В связи со слабым развитием придаточных корней, образовавшиеся раметы (парциальные побеги и кусты) нежизнеспособны, они быстро погибают.
Рис. 2.
Полукустарничек Thymus reverdattoanus.
1 – составная скелетная ось, 2 – двулетний вегетативный побег, 3 – генеративный побег, 4 – придаточный корень, 5 – главный корень.
Fig. 2. Dwarf subshrub Thymus reverdattoanus.
1 – composite bough axis, 2 – biannual vegetative shoot, 3 – generative shoot, 4 – adventitious root, 5 – main root.
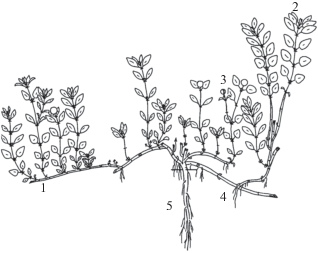
Таблица 1.
Геоботаническая характеристика сообществ с участием Thymus reverdattoanus Table 1. Geobotanical description of the plant communities with Thymus reverdattoanus
| Ярус Layer |
Полевой номер описания/Field relevé number | Постоянство Constancy |
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L13-550 | L13-551 | L13-555 | L13-556 | L13-557 | L13-558 | L13-553 | L13-554 | L13-552 | L13-559 | |||
| Табличный номер описания/Table relevé number | ||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |||
| Средняя высота древесного яруса, м Average height of tree layer, m |
26 | 24 | 28 | 26 | 24 | 26 | – | – | – | – | ||
| Сомкнутость крон древесного яруса Crown closure of tree layer |
0.7 | 0.5 | 0.6 | 0.5 | 0.6 | 0.5 | – | – | – | – | ||
| Проективное покрытие кустарникового яруса, % Shrub layer projective coverage, % |
+ | 2 | + | + | + | + | + | + | + | + | ||
| Проективное покрытие травяно-кустарничкового яруса, % Herb-dwarf shrub layer projective coverage, % |
90 | 75 | 90 | 85 | 85 | 80 | 55 | 75 | 65 | 70 | ||
| Проективное покрытие мохово-лишайникового яруса, % Moss-lichen layer projective coverage, % |
40 | 100 | 100 | 100 | 100 | 100 | – | – | – | – | ||
| Число видов сосудистых растений Number of vascular plant species |
14 | 12 | 13 | 13 | 14 | 11 | 11 | 10 | 12 | 13 | ||
| Диагностические виды порядка Ledo palustris-Laricetalia/Diagnostic species of order Ledo palustris-Laricetalia | ||||||||||||
| Ledum palustre | c | + | + | + | . | + | . | . | . | . | . | II |
| Vaccinium uliginosum | c | + | + | + | . | . | . | . | . | . | . | II |
| Sphagnum girgensohnii | d | 1 | + | + | + | . | + | . | . | . | . | III |
| Polytrichum commune | d | + | + | + | + | + | + | . | . | . | . | III |
| Диагностические виды класса Vaccinio-Piceetea/Diagnostic species of class Vaccinio-Piceetea | ||||||||||||
| Larix sibirica | a1 | 2 | 3 | 2 | 3 | 4 | 2 | . | . | . | . | III |
| Pinus sibirica | a1 | 3 | + | 1 | 2 | + | 3 | . | . | . | . | III |
| Picea obovata | a2 | 3 | 3 | . | 3 | 3 | 3 | . | . | . | . | III |
| Pinus sibirica | a2 | . | . | . | + | + | 2 | . | . | . | . | II |
| Larix sibirica | a2 | 1 | 2 | . | . | . | 1 | . | . | . | . | II |
| Picea obovata | a3 | . | + | 2 | + | + | + | . | . | . | . | III |
| Pinus sibirica | a3 | + | + | . | . | + | + | . | . | . | . | II |
| Vaccinium vitis-idaea | c | 5 | 5 | 5 | 5 | 5 | 5 | + | . | 1 | . | IV |
| Orthilia secunda | c | + | + | + | + | + | + | . | . | . | + | IV |
| Linnaea borealis | c | + | + | + | + | + | + | . | . | . | . | III |
| Pleurozium schreberi | d | 2 | 4 | 4 | 4 | 4 | 4 | . | . | . | . | III |
| Hylocomium splendens | d | 2 | 3 | 2 | 2 | 2 | 2 | . | . | . | . | III |
| Ptilium crista-castrensis | d | 1 | 1 | 2 | 2 | 2 | 2 | . | . | . | . | III |
| Прочие виды/Other species | ||||||||||||
| Betula pubescens | a2 | 1 | . | . | + | + | . | . | . | . | . | II |
| Betula pubescens | a3 | + | + | 1 | + | + | + | . | + | . | . | IV |
| Duschekia fruticosa | b | + | . | + | + | + | + | . | . | . | . | III |
| Sorbus sibirica | b | + | + | . | + | + | + | . | . | . | . | III |
| Calamagrostis lapponica | c | + | . | + | . | + | . | . | . | . | . | II |
| Peltigera membranacea | d | + | + | + | . | . | + | . | . | . | . | II |
| Диагностические виды класса Epilobietea angustifolii/Diagnostic species of class Epilobietea angustifolii | ||||||||||||
| Calamagrostis epigeios | c | . | . | . | . | . | . | 3 | 4 | 2 | 4 | II |
| Прочие виды/Other species | ||||||||||||
| Tanacetum bipinnatum | c | . | . | . | . | . | . | + | 2 | + | 1 | II |
| Thymus reverdattoanus | c | . | . | . | . | . | . | 1 | 1 | 1 | + | II |
| Rosa acicularis | b | . | + | . | . | . | . | + | + | + | + | III |
| Hieracium umbellatum | c | . | . | . | . | . | . | + | + | + | + | II |
| Festuca ovina | c | . | . | . | . | . | . | + | + | + | + | 4 |
| Solidago virgaurea | c | . | . | . | . | . | . | + | + | + | + | II |
| Equisetum arvense | c | . | . | . | . | . | . | + | + | + | + | II |
| Polytrichum juniperinum | d | . | . | . | . | . | . | . | + | + | + | II |
| Calamagrostis langsdorffii | c | + | . | . | + | . | . | 1 | . | + | + | III |
Примечание. Точкой в ячейках таблицы обозначено отсутствие вида. Ярус: a — древесный (a1, a2, a3 — подъярусы), b — кустарниковый, c — травяно-кустарничковый, d — мохово-лишайниковый.
Note. Dots in the table cells indicate absence of species. Layers: a – tree layer (a1, a2, a3 — sublayers), b — shrub layer, c — herb-dwarf shrub layer, d — moss-lichen layer.
Встречены 1–2 раза/Recorded 1–2 times: Aconogonon ochreatum [c] (10 +), Carex globularis [c] (4 +, 5 +), Chamaenerion angustifolium [c] (7 +, 9 1), Dicranum sp. [d] (2 +), Empetrum nigrum [c] (1 +, 3 +), Goodyera repens [c] (6 +), Juniperus communis [b] (1 +, 3 +), Nephroma arcticum [d] (2 +, 3 +), Peltigera aphthosa [d] (1 +, 3 +), Picea obovata [a1] (1 +, 3 3), Poa alpigena [c] (10 +), Pogonatum sp. [d] (7 +, 8 +), Polytrichum piliferum [d] (9 +), Ptilidium ciliare [d] (1 +, 2 +), Rubus chamaemorus [c] (4 +, 5 +), Rumex thyrsiflorus [c] (8 +, 10 +), Trientalis europaea [c] (2 +, 4 +), Vaccinium myrtillus [c] (5 +, 6 +).
Координаты описаний/Coordinates of relevés: 1 – N 64.78759° – E 72.53608°, 2 – N 64.78629° – E 72.53687°, 3 – N 64.78722° – E 72.53846°, 4 – N 64.78681° – E 72.54121°, 5 – N 64.78547° – E 72.54185°, 6 – N 64.78461° – E 72.53785°, 7 – N 64.78575° – E 72.53698°, 8 – N 64.78575° – E 72.53764°, 9 – N 64.78580° – E 72.53639°, 10 – N 64.78461° – E 72.53689°.
Автор описаний: Н.Н. Лащинский.
Author of the relevés: N.N. Lashchinskiy.
Анализ структуры особей T. reverdattoanus позволил выявить ряд специфических особенностей, связанных с условиями произрастания. Так, сомкнутый травостой, высота которого достигает 55 см, способствует значительному удлинению ортотропных побегов до 20–30 см. Песчаный субстрат и его высокая влажность являются благоприятными условиями для вегетативного разрастания T. reverdattoanus, но вместе с этим наличие многолетнемерзлых грунтов и длительное сезонное промерзание субстрата препятствуют развитию мощной придаточной корневой системы, и как следствие выживанию рамет. У особей вида на протяжении всей жизни сохраняется главный корень и моноцентрическая биоморфа.
ОБСУЖДЕНИЕ
Отличительными особенностями нового местонахождения T. reverdattoanus являются эколого-ценотические условия и удаленность от основной части ареала. Поскольку обнаруженная популяция отделена широкой долиной Оби, представляющей существенную преграду для миграции видов, невозможно предположить непрерывный ареал между этим и ранее известными местонахождениями. В литературе представлены разные точки зрения о распространении тимьянов в северных районах (Klokov, 1973; Menitskii, Yurtsev, 1980; Gogina, 1990). Согласно Гогиной при расселении из Средиземноморской области на восток тимьяны пережили эпоху оледенения в горах Восточной Сибири, а затем в благоприятные эпохи расширили ареал к северу и достигли своего климатического предела распространения (Gogina, 1990). Новое местонахождение T. reverdattoanus в отрыве от основного ареала позволяет предположить, что вид попал на эту территорию по речным долинам, которые стали одним из коридоров миграции тимьянов с юга на север. В пределах ареала вид проявляет себя не только как петрофит, приуроченный преимущественно к горным породам, но также произрастает и на песчаном субстрате. Разнообразие типов субстрата отмечается в местообитаниях и других видов тимьянов: T. baicalensis Serg. (Talovskaya (Kolegova), 2015), T. mongolicus (Ronn.) Ronn. (Kolegova, Cheryomushkina, 2015), Thymus roseus Schipcz. (Talovskaya et al., 2017). Способность вида произрастать как на каменистом, так и на песчаном субстрате и отсутствие приуроченности к горным породам определенного химизма существенно расширяет возможности его расселения и миграций. Все изученные нами местообитания вида связаны с аккумулятивными песчаными наносами в речной пойме. Практически полное отсутствие вегетативного размножения и поддержание ценопопуляции за счет семенного возобновления указывает на пионерную стратегию вида, осваивающего вновь возникающие песчаные косы в речной долине и существующего в течение определенной сукцессионной стадии. Повторяемость подобных местообитаний, обусловленная закономерностями развития речной долины, обеспечивает неопределенно долгое существование вида на данной территории и его распространение в пределах речной сети. Анализ структуры особей T. reverdattoanus позволил выявить ряд специфических особенностей (значительное удлинение побегов, слабое развитие придаточных корней, практически полное отсутствие рамет, моноцентрическая биоморфа), обусловленных особенностями эколого-ценотических условий произрастания. Вместе с этим, стоит отметить, что полукустарничковая жизненная форма у T. reverdattoanus сохраняется на протяжении всего ареала. Такая стабильность не случайна, анализ литературы о жизненных формах тимьянов показал, что жизненная форма полукустарничка является наиболее прогрессивной, ее морфологическая пластичность увеличивает выживаемость особей в неоднородных и неблагоприятных условиях обитания (Klokov, 1973; Berko, 1988; Gogina, 1990).
ЗАКЛЮЧЕНИЕ
Новое местонахождение T. reverdattoanus и анализ условий его произрастания существенно расширяют представления об экологии вида. Вид обладает экологической пластичностью, осваивая не только каменистые, но и песчаные субстраты различного химизма. Пионерный характер сообществ, в составе которых тимьян произрастает в Западной Сибири, позволяет предположить возможность распространения вида по речным долинам, что существенно дополняет сведения о путях и способах миграции рода в целом.
Анализ структуры особей в новых условиях обитания выявил значительную морфологическую пластичность вида при сохранении базовой жизненной формы. Пластичность носит адаптивный характер и позволяет виду реализовывать пионерную стратегию в условиях динамичных местообитаний и сукцессивных сообществ речных пойм.
Список литературы
Andreev M.P., Kotlov Yu.V., Makarova I.I. 1996. Checklist of the lichens and lichenicolous fungi of the Russian Arctic. – The Bryologist. 99 (2): 137–169. https://doi.org/10.2307/3244545
Becking R. 1957. The Zürich-Montpellier school of phytosociology. – Bot. Rev. 23 (7): 411–488.
[Berko] Берко Й.М. 1988. Типи пагонових систем i життεвi форми видiв секцiϊ Verticillati (Klok. et Shost.) Klok. роду Thymus L. флори Украϊни. – Укр. ботан. журн. 45 (1): 27–32.
[Bobrov] Бобров Ю.А. 2009. Грушанковые России. Киров. 130 с.
[Brodskii] Бродский А.К. 2016. Биоразнообразие: структура, проблемы и перспективы сохранения. – Сборник трудов Зоологического музея МГУ им М.В. Ломоносова. 54: 380–396.
[Bykov] Быков Б.А. 1979. К количественной оценке эндемизма. – Бот. мат. Герб. Ин-та ботаники АН КазССР. 11: 3–8.
[Czerepanov] Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб. 992 с.
[Doron‘kin] Доронькин В.М. 1997. 27. Thymus L. – тимьян, богородская трава – В кн.: Флора Сибири. Pyrolaceae-Lamiaceae (Labiatae). Т. 11. Новосибирск. С. 205–220.
[Endemichnye…] Эндемичные растения Урала во флоре Свердловской области. 2013. Екатеринбург. 612 с.
[Gogina] Гогина Е.Е. 1990. Изменчивость и формообразование в роде Тимьян. М. 208 с.
Ignatov M.S., Afonina O.M., Ignatova E.A. 2006. With contributions on regional floras from: Abolina A., Akatova T.V., Baisheva E.Z., Bardunov L.V., Baryakina E.A., Belkina O.A., Bezgodov A.G., Boychuk M.A., Cherdantseva V.Ya., Czernyadjeva I.V., Doroshina G.Ya., Dyachenko A.P., Fedosov V.E., Goldberg I.L., Ivanova E.I., Jukoniene I., Kannukene L., Kazanovsky S.G., Khar-zinov Z.Kh., Kurbatova L.E., Maksimov A.I., Mamatkulov U.K., Manakyan V.A., Maslovsky O.M., Napreenko M.G., Otnyukova T.N., Partyka L.Ya., Pisarenko O.Yu., Popova N.N., Rykovsky G.F., Tubanova D.Ya., Zheleznova G.V., Zolotov V.I. Checklist of mosses of East Europe and North Asia. – Arctoa. 15: 1–130. https://doi.org/10.15298/arc-toa.15.01
[Klokov] Клоков М.В. 1954. Род Thymus L. – Тимьян. – В кн.: Флора СССР. Т. 21. М., Л. С. 470–591.
[Klokov] Клоков В.М. 1973. Расообразование в роде тимьянов – Thymus L. на территории Советского Союза. Киев. 190 с.
Kolegova E.B., Cheryomushkina V.A. 2015. Ontogenetic structure and assessment of state of Thymus mongolicus (Lamiaceae) coenopopulations in Southern Siberia. – Contemporary problems of ecology. 8 (2): 155–161. https://doi.org/10.15372/SEJ20150203
[Krasnaya…] Красная книга Ямало-Ненецкого автономного округа: животные, растения, грибы. 2010. Екатеринбург. 308 с.
[Kuvaev] Куваев В.Б. 2006. Флора субарктических гор Евразии и высотное распределение ее видов. М. 568 с.
[Mazurenko, Khokhryakov] Мазуренко М.Т., Хохряков А.П. 1977. Структура и морфогенез кустарников. М. 158 с.
[Menitskii, Yurtsev] Меницкий Ю.Л., Юрцев Б.А. 1980. Род Thymus L. – Тимьян, чабрец. – В кн.: Арктическая флора СССР. Вып. 8. Л. С. 255–262.
[Nukhimovskii] Нухимовский Е.Л. 1997. Основы биоморфологии семенных растений. Т. 1. М. 630 с.
[Pospelova, Pospelov] Поспелова Е.Б., Поспелов И.Н. 2007. Флора сосудистых растений Таймыра и сопредельных территорий. Часть 1. Аннотированный список флоры и ее общий анализ. М. 457 с.
[Serebryakov] Серебряков И.Г. 1959. Типы развития побегов у травянистых многолетников и факторы их формирования. – Ученые записки МГПИ им. В.П. Потёмкина. Вопросы биологии растений. 100 (5): 3–38.
[Serebryakov] Серебряков И.Г. 1964. Жизненные формы высших растений и их изучение. – Полевая геоботаника. 3: 146–208.
[Smirnova] Смирнова О.В. 1987. Структура травяного покрова широколиственных лесов. М. 205 с.
Talovskaya (Kolegova) E.B. 2015. Thymus baicalensis (Lamiaceae) morphological transformation under different environmental conditions. – Contemporary Problems of Ecology. 6: 736–749. https://doi.org/10.1134/S1995425515050133
[Talovskaya et al.] Таловская Е.Б., Черёмушкина В.А., Асташенков А.Ю., Гусева А.А. 2017. Mорфологическая адаптация Thymus roseus (Lamiaceae) в Восточном Казахстане. – Бот. журн. 102 (9): 1232–1248.
[Tolmachov] Толмачёв А.И. 1974. Эндемизм. – В кн.: Введение в географию растений. Л. С. 147–160.
[Zverev] Зверев А.А. 2007. Информационные технологии в исследованиях растительного покрова. Томск. 303 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал