

Ботанический журнал, 2020, T. 105, № 7, стр. 672-686
ИЗМЕНЕНИЯ РАСТИТЕЛЬНОСТИ ДУБРАВЫ “ЛЕС НА ВОРСКЛЕ” ЗАПОВЕДНИКА БЕЛОГОРЬЕ ЗА 60 ЛЕТ
Д. М. Мирин 1, *, М. Ю. Тиходеева 1
1 Санкт-Петербургский государственный университет
199034 Санкт-Петербург, Университетская наб, 7/9, Россия
* E-mail: mirin_denis@mail.ru
Поступила в редакцию 10.02.2020
После доработки 10.04.2020
Принята к публикации 14.04.2020
Аннотация
На участке “Лес на Ворскле” заповедника “Белогорье” сохранились лесостепные дубравы с незначительными признаками антропогенной трансформации. Эти растительные сообщества считались устойчивыми, коренными, однако за последние чуть более 60 лет в них произошли существенные изменения, зафиксированные геоботаническими описаниями в 1954, 1984 и 2018 годах. В течение всего времени наблюдений в древостое уменьшалось участие дуба Quercus robur и увеличивалось участие липы Tilia cordata, ясеня Fraxinus excelsior и, особенно, клена Acer platanoides. С 1954 года древостой стал более тенистым с лучше выраженными нижними его ярусами. Нарастающими темпами идет вывал старых деревьев первого яруса. Практически полностью исчез светолюбивый подрост дуба, появился подрост ильма. Произошло полное зарастание полян, которые раньше поддерживались кошением. Сильно изменился состав подпологовой растительности. Из подлеска исчезли относительно светолюбивые ксеромезофильные виды, такие как терн Prunus spinosa, появились гигромезофильные виды, такие как черемуха Prunus padus. Видовое разнообразие и насыщенность и общее проективное покрытие травостоя резко уменьшилось в первую половину срока наблюдений и увеличилось за вторую половину срока. Из травостоя за первые 30 лет наблюдений исчезли ксеромезофитные и относительно светолюбивые виды (Agrimonia eupatoria, Astragalus glycyphyllos, Galium verum и др.). За последние 30 лет разрослись в первую очередь теневыносливые нитрофилы (Lamium maculatum, Sambucus nigra), появились инвазионные виды (Acer negundo, Erigeron annuum). Эти изменения носят необратимый характер, т.е. являются отражением идущей сукцессии. Основные причины сукцессии: (1) естественная смена лесов с резким преобладанием дуба на полидоминантные широколиственные леса с малым участием дуба, предопределенная эколого-биологическими особенностями возобновления древесных видов и достижением основным поколением дуба возраста средней продолжительности его жизни, (2) увеличение влажности климата, (3) прекращение выпаса в послевоенные годы, (4) увеличение численности популяции кабанов.
Подавляющее большинство современных европейских лесов сформировались в результате длительного антропогенного воздействия и далеки от их естественного облика (Küster, 2018). Что касается территории Восточной Европы, то под максимальным антропогенным прессом оказались степи, лесостепи и широколиственные леса. Сохранившиеся на сегодняшний день малые островки этих зональных сообществ сильно трансформированы (Antipina, Maslov, 1994; Bobrovsky, 2010; Ponomarenko et al., 2015). Их заповедование из-за нарушения природной динамики, обусловленной малыми площадями и отсутствием ключевых видов, не приводит к формированию коренных сообществ. В ряде случаев прекращение всех видов пользования, имитирующих в некоторой степени воздействие зоогенного компонента естественных биоценозов, провоцируют полную их деструкцию. Мониторинговое изучение растительного покрова и оптимизация возможных режимов охраны остаются приоритетными задачами при организации особо охраняемых природных территорий.
Участок “Лес на Ворскле” государственного природного заповедника “Белогорье” (Белгородская область) занят, в основном, лесостепной нагорной дубравой, расположен в непосредственной близости от п. Борисовка и до 1999 года имел статус отдельного государственного заповедника. Этот лесной массив площадью 1038 га разделен на 10 кварталов, занимает высокий правый берег реки Ворсклы (левый приток Днепра). В начале XXI века большая часть площадей приходится на 100–150-летние древостои, возраст которых в 1965 году был соответственно 50–80 и 80–100 лет (Neshataev, 1967). Коренными лесными формациями здесь считаются дубняки из дуба черешчатого: ясене-дубняки и липо-дубняки. Наиболее ценными признаются участки условно коренных высокоствольных дубрав, возраст которых к концу 1990-х гг. по оценкам Ю.Н. Нешатаева, В.Н. Ухачевой (Neshataev, Ukhacheva, 2001) составил около 300 лет. В конце 1950-х гг. площадь этих высокоствольных дубрав в заповеднике составляла 160 га, но к середине 1990-х гг. из-за естественной гибели старых деревьев сократилась до 130 га (Neshataev, Ukhacheva, 2001). Их место оказалось занято липняками и кленовниками. На сегодняшний день остатки уникальных 300‑летних лесов сохранились в 1, 5, 8 и 10 кварталах заповедника (Ryabtsev et al, 2009).
“Лес на Ворскле” – один из старейших заповедных лесных массивов РФ. В XVII в. здесь проходила “Белгородская засечная черта” – 800-километровое уникальное фортификационное сооружение, защищающее южные границы Русского государства от набегов кочевников. С 1706 г. (со времен начала владения этими землями графом Б.П. Шереметевым) до конца XIX века этот массив использовался как охотничье хозяйство и имел статус “Заказной Рощи”. Лес на этой территории охранялся весьма строго, и лишь в отдельных его кварталах в начале XIX века для хозяйственных нужд были разрешены выборочные рубки (Goryshina, 1986). В конце ХIХ века и в революционные и послереволюционные годы рубка леса в дубраве приняла стихийный характер. Но в 1923 году, благодаря усилиям известного энтомолога С.И. Малышева, на территории бывшего графского леса был образован заповедник “Лес на Ворскле” (Shapovalov, 2005). В 1934 году заповедник в качестве учебной и научной базы был передан под юрисдикцию Ленинградского университета. Основателем и первым директором базы стал выдающийся геоботаник академик Владимир Николаевич Сукачев. Первостепенными задачами базы было осуществление практической части образовательного процесса для студентов биолого-почвенного и географического факультетов Ленинградского университета и проведение комплексных исследований природы с целью разработки мер рационального природопользования. Развернувшиеся и плодотворно реализуемые научные и образовательные работы были прерваны Великой Отечественной войной. На этих землях велись активные боевые действия. На территории заповедника были размещены многочисленные схроны боеприпасов, а по периметру лесного массива были установлены артиллерийские батареи. Лес вырубался для хозяйственных нужд и использовался под выпас скота, о чем свидетельствуют сохранившиеся единичные геоботанические описания 1947–1949 гг. Но после войны научные работы и летняя практика студентов-биологов вновь возобновились. С тех пор благодаря многолетним научным изысканиям преподавателей, аспирантов и студентов Ленинградского (ныне Санкт-Петербургского) государственного университета накоплены уникальные научные данные.
Материалом для этой работы послужили серии геоботанических описаний пробных площадей (20×20 м), проведенные в южной половине 8 квартала заповедника в 1954 (23 описания), 1984 (23 описания) и 2018 (34 описания) годах преподавателями и студентами кафедры геоботаники ЛГУ (СПбГУ). Геоботанические описания 1954 и 1984 годов выполнены под руководством Ю.Н. Нешатаева, в 2018 г. работы проведены под руководством Д.М. Мирина. Описания содержат подробную географическую и топографическую привязку, характеристику гидрологических и почвенных условий. Детальное описание древостоя включает пересчет деревьев каждого вида по ярусам с указанием максимальной и господствующей высоты, диаметра ствола и класса возраста. Дано описание подроста и подлеска с указанием жизненности и количества по высотным рангам для каждого вида. Характеристика напочвенного покрова включает общее проективное покрытие (%) и полный список видов сосудистых растений с указанием обилия. Обилие в разные годы описаний указано в разных единицах, поэтому для анализа переведено в баллы шкалы Браун-Бланке. При исследовании изменений состава и строения напочвенного покрова в данной статье использованы данные только по константности видов. Моховой напочвенный покров на этом участке дубравы отсутствовал. Также отмечена интенсивность и характер зоогенного (порои, лежки и чесалки кабанов, кротовины, муравейники, следы деятельности насекомых-фитофагов) и антропогенного (следы и возраст рубок, тропинки) воздействия (Ipatov, Mirin, 2008). Названия видов в данной статье приведены в соответствии с актуальной информацией на ресурсе ThePlantList.org.
Восьмой квартал (объект исследования) занимает восточную часть лесного массива и захватывает оконечность пологого склона верхней террасы, высота которой достигает на территории участка “Лес на Ворскле” 217 м над ур. м. Грунтовые воды залегают здесь на глубине 25–30 м и недоступны растениям (Neshataev, 1967).
Характер растительного покрова в этом квартале типичен для всего лесного массива и представлен широколиственными лесными сообществами из Quercus robur L., Fraxinus excelsior L., Tilia cordata Mill., Acer platanoides L. Древостой, как правило, двухъярусный с выраженным подлеском из Euonymus europaeus L., E. verrucosus Scop., Crataegus rhipidophylla Gand. и др. В первом ярусе преимущественно доминирует Quercus robur. Второй ярус в основном образуют те же породы, что и первый (перечисленные выше) с единичной примесью Ulmus glabra Huds., Betula pendula Roth, Pyrus pyraster (L.) Burgsd.
В напочвенном покрове в роли доминантов обычно выступают Aegopodium podagraria, Carex pilosa, C. spicata, Geranium robertianum, Lamium maculatum, Lysimachia nummularia, Poa nemoralis, Urtica dioica, высокое постоянство в настоящее время демонстрируют Asarum europaeum, Galium aparine, Geum urbanum, Glechoma hederacea, Moehringia trinervia, Polygonatum multiflorum, Pulmonaria obscura, Scrophularia nodosa, Stellaria holostea, Viola mirabilis, Viola suavis. Общий список встреченных на пробных площадях видов трав с указанием их константности приведен в табл. 1.
Таблица 1.
Константность видов трав в дубраве в три периода описаний, % Table 1. Constancy of herb species in the oak-wood in three terms of descriptions, %
| Виды/Species | 1954 | 1984 | 2018 |
|---|---|---|---|
| Achillea millefolium L. | 30 | 0 | 0 |
| Adoxa moschatellina L. | 9 | 13 | 0 |
| Aegopodium podagraria L. | 30 | 26 | 18 |
| Agrimonia eupatoria L. | 35 | 0 | 0 |
| Ajuga genevensis L. | 17 | 0 | 0 |
| Alliaria petiolata (M. Bieb.) Cavara & Grande | 0 | 0 | 12 |
| Artemisia vulgaris L. | 9 | 0 | 9 |
| Asarum europaeum L. | 9 | 35 | 38 |
| Astragalus glycyphyllos L. | 70 | 0 | 3 |
| Athyrium filix-femina (L.) Roth | 0 | 0 | 6 |
| Brachypodium sylvaticum (Huds.) P. Beauv. | 17 | 0 | 0 |
| Campanula rapunculoides L. | 87 | 9 | 3 |
| Campanula trachelium L. | 13 | 9 | 3 |
| Cardamine impatiens L. | 0 | 4 | 6 |
| Carex pilosa Scop. | 70 | 52 | 15 |
| Carex praecox Schreb. | 26 | 0 | 0 |
| Carex spicata Huds. | 17 | 0 | 15 |
| Chaerophyllum temulum L. | 0 | 0 | 6 |
| Chelidonium majus L. | 0 | 0 | 9 |
| Chenopodium album L. | 9 | 0 | 0 |
| Cirsium vulgare (Savi) Ten. | 4 | 0 | 6 |
| Clinopodium vulgare L. | 48 | 0 | 0 |
| Cystopteris fragilis (L.) Bernh. | 0 | 0 | 6 |
| Dactylis glomerata L. | 43 | 4 | 0 |
| Dryopteris carthusiana (Vill.) H.P. Fuchs | 17 | 0 | 6 |
| Elymus repens (L.) Gould | 9 | 0 | 0 |
| Erigeron annuus (L.) Desf. | 0 | 0 | 9 |
| Fallopia dumetorum (L.) Holub | 26 | 0 | 9 |
| Fragaria moschata Weston | 26 | 0 | 0 |
| Fragaria vesca L. | 83 | 9 | 0 |
| Galium aparine L. | 26 | 13 | 38 |
| Galium odoratum (L.) Scop. | 9 | 4 | 15 |
| Galium verum L. | 43 | 0 | 0 |
| Geranium robertianum L. | 39 | 35 | 56 |
| Geum urbanum L. | 83 | 78 | 41 |
| Glechoma hederacea L. | 78 | 57 | 50 |
| Hypericum perforatum L. | 74 | 0 | 9 |
| Impatiens noli-tangere L. | 0 | 13 | 12 |
| Lactuca quercina L. | 17 | 0 | 0 |
| Lamium maculatum (L.) L. | 0 | 30 | 62 |
| Lapsana communis L. | 22 | 0 | 6 |
| Lysimachia nummularia L. | 83 | 9 | 6 |
| Melilotus officinalis (L.) Pall. | 9 | 0 | 0 |
| Moehringia trinervia (L.) Clairv. | 0 | 4 | 56 |
| Myosotis ramosissima Rochel | 17 | 0 | 0 |
| Origanum vulgare L. | 26 | 0 | 0 |
| Plantago lanceolata L. | 22 | 0 | 0 |
| Plantago media L. | 22 | 0 | 0 |
| Poa angustifolia L. | 13 | 0 | 0 |
| Poa nemoralis L. | 87 | 13 | 3 |
| Polygonatum multiflorum (L.) All. | 0 | 57 | 47 |
| Prunella vulgaris L. | 22 | 0 | 0 |
| Pulmonaria obscura Dumort. | 30 | 17 | 21 |
| Scrophularia nodosa L. | 35 | 13 | 47 |
| Securigera varia (L.) Lassen | 17 | 0 | 0 |
| Silene viscaria (L.) Jess. | 17 | 0 | 0 |
| Silene vulgaris (Moench) Garcke | 9 | 0 | 0 |
| Stachys sylvatica L. | 4 | 9 | 12 |
| Stellaria holostea L. | 52 | 30 | 18 |
| Taraxacum officinalis (L.) Weber ex F.H. Wigg. | 26 | 0 | 6 |
| Torilis japonica (Houtt.) DC. | 74 | 0 | 0 |
| Trifolium pratense L. | 9 | 0 | 0 |
| Trifolium repens L. | 4 | 0 | 6 |
| Urtica dioica L. | 35 | 17 | 53 |
| Veronica chamaedrys L. | 70 | 0 | 12 |
| Vicia sepium L. | 22 | 17 | 0 |
| Viola arvensis Murray | 17 | 4 | 0 |
| Viola hirta L. | 48 | 9 | 0 |
| Viola mirabilis L. | 17 | 0 | 38 |
| Viola odorata L. | 26 | 0 | 0 |
| Viola suavis M. Bieb. | 43 | 70 | 65 |
С константностью менее 5% были также отмечены следующие виды / Also, the following species were found with constancy less than 5%: Actaea spicata L. (2018), Agrostis capillaris L. (1954), Anemone ranunculoides L. (1954), Anthriscus sylvestris (L.) Hoffm. (1984), Artemisia campestris L. (1954), Campanula persicifolia L. (1984), Capsella bursa-pastoris (L.) Medik. (1954), Carduus acanthoides L. (2018), Carduus crispus Guirão ex Nyman (2018), Chenopodium hybridum L. (1954), Epilobium montanum L. (1954, 2018), Galeopsis tetrahit L. (2018), Gypsophila muralis L. (1954), Hepatica nobilis Mill. (1954), Hypericum hirsutum L. (1954), Leucanthemum vulgare Lam. (1954), Mercurialis perennis L. (1984), Pastinaca sativa L. (1954), Plantago major L. (1954, 1984), Ranunculus acris L. (1954), Ranunculus cassubicus L. (1984), Silene latifolia Poir. (1954), Stellaria graminea L. (1954), Stellaria media (L.) Vill. (1954, 2018), Trifolium arvense L. (1954), Verbascum nigrum L. (1954), Verbascum thapsus L. (1954), Veronica officinalis L. (1954).
По флористической классификации описанные сообщества относятся к ассоциации Fraxino excelsioris–Quercetum roboris Bulokhov et Solomeshch 2003, субассоциации F.e.–Q.r. crataegetosum curvisepalae Semenishchenkov 2015 (Bulokhov, Semenishchenkov, 2015; Semenishchenkov et al, 2018). Состав и строение древесного полога разных участков в пределах исследуемого квартала имеет ряд различий. Это с одной стороны предопределено неоднородностью рельефа: здесь выделяются плакорные участки, дно оврага и его склоны, а с другой – характером многовековой хозяйственной деятельности, а именно: на разных участках проходили разные типы рубок, с разной интенсивностью и в разное время, что, привело к формированию лесных сообществ разного возрастного состава древостоя.
Проводя общую оценку растительного покрова, следует сразу отметить, что за 60-летний период исследований и древостой, и напочвенный покров в условиях заповедного режима претерпели значительные изменения. Анализу этих изменений и посвящена данная работа.
АНАЛИЗ ИЗМЕНЕНИЙ В ДРЕВЕСНОМ ПОЛОГЕ
Анализ всех 80 описаний за 1954, 1984 и 2018 гг. показал, что в 90% описаний дубравы основную роль в древостое играет Quercus robur, в 4% – Acer platanoides, по 3% – Fraxinus excelsior и Tilia cordata. Лидирующая роль дуба в сложении древесного яруса отражена в диаграммах, построенных на основании анализа коэффициентов участия, рассчитанных по формуле древостоя (рис. 1). За 60-летний период наблюдений дуб, сохраняя лидирующие позиции, тем не менее, снизил степень участия в древостое: если в 1954 году в формуле древостоя преобладают у дуба коэффициенты 8–9–10 (например, 8Д2Я+Л, 9Д1Л+Кл, 10Д), то в 2018 году они уменьшились до 5–6–7 (5Д3Л2Кл, 6Д3Кл1Л, 7Д2Кл1Я). Роль клена в сложении древостоя, наоборот, усилилась: в 1954 г. на 74% пробных площадей клен в древостое отсутствовал, в 2018 г. лишь 4% пробных площадей были без клена, а коэффициенты участия клена в формуле древостоя достигли 4–5–6 (4Д4Кл2Я, 5Кл4Д1Л, 6Кл3Д1Л). Увеличение роли в древостое продемонстрировали ясень и липа, но по сравнению с кленом не так сильно. Выявленные закономерности проявились и в ярусном строении древостоя: за 60 лет в первом ярусе дуб сохранился почти на всех пробных площадях, ясень, липа и клен усилили свое присутствие в первом ярусе (рис. 2). Во втором ярусе значительно возросла роль клена (рис. 3).
Рис. 1.
Участие древесных пород по формуле древостоя. По оси абсцисс: значение коэффициента по формуле древостоя; по оси ординат: частота сообществ, %.
Fig. 1. Participance of tree species by tree stand index. X-axis: value of tree stand index; Y-axis: frequency of communities, %.
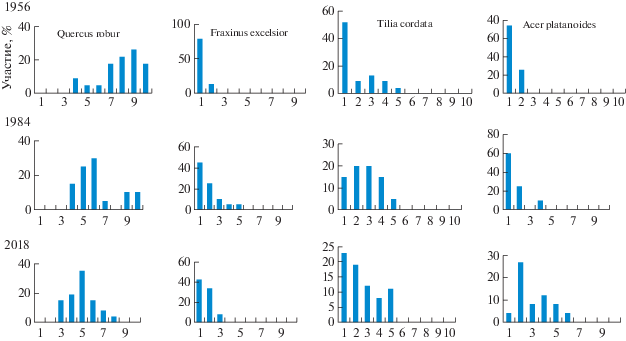
Рис. 2.
Встречаемость видов в 1 ярусе древостоя, %.
Fig. 2. Constancy of species in the 1st layer of tree stand, %.
По оси абсцисс/X-axis: 1 – Quercus robur, 2 – Fraxinus excelsior, 3 – Tilia cordata, 4 – Acer platanoides, 5 – Ulmus glabra.
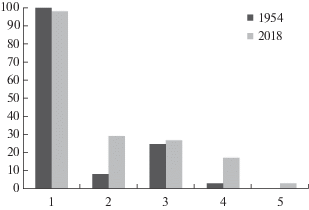
Рис. 3.
Встречаемость видов во 2 ярусе древостоя, %.
Fig. 3. Constancy of species in the 2nd layer of tree stand, %.
По оси абсцисс/X-axis: 1 – Quercus robur, 2 – Fraxinus excelsior, 3 – Tilia cordata, 4 – Acer platanoides, 5 – Ulmus glabra, 6 – Pyrus pyraster, 7 – Betula pendula.
Толщина стволов косвенно отражает возраст дерева. Распределение пробных площадей, описанных в 1954 и 2018 гг., по ступеням толщин стволов дуба первого яруса (рис. 4) свидетельствует о том, что исследованные лесные сообщества относятся к двум возрастным группам. Возраст древостоев первой группы (ступени толщин 20–30, 31–40, 41–50 см в диаметре) в 1954 году составил 40–70 лет (в 2018, соответственно, 100–130 лет) – эти леса сформировались на месте сплошных рубок конца XIX – начала XX веков. Возраст древостоев второй группы (ступени толщин от 70 до более 100 см в диаметре) в 1954 году был 200–300 лет (в 2018, соответственно, 260–360 лет) – это перестойные древостои, не подвергавшиеся сплошным рубкам, близкие к коренным лесным сообществам.
Рис. 4.
Распределение пробных площадей по ступеням толщины дуба в 1 ярусе древостоя.
По оси абсцисс: ступени диаметров толщин дуба, см. По оси ординат: число пробных площадей.
Fig. 4. Distribution of sample plots by classes of 1st layer’s oak stem diameter.
X-axis: ranges of oak stem diameter, cm. Y-axis: number of sample plots.
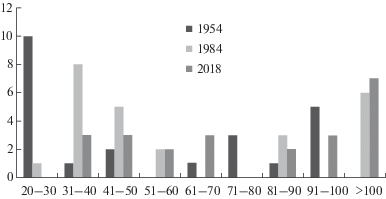
За исследуемый период заметно возросла сомкнутость древесного полога: средняя сомкнутость на древостой пробных площадей в 1954 году составила 0.48 (от 0.2 до 0.8), в 1984 – 0.54 (от 0.3 до 0.9), а в 2018 – 0.88 (от 0.6 до 0.9).
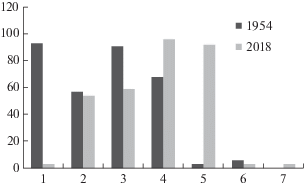
За 60 лет изменился и состав подроста: резко упало присутствие подроста дуба (рис. 5). Если в 1954 году дуб в подросте присутствовал почти на всех пробных площадях и был представлен особями 3–20-летнего возраста в удовлетворительном состоянии плотностью 1200–2500 экз./га, то в 2018 году только на одной пробной площади были зафиксированы проростки дуба до 3 лет с плотностью 250 экз./га. Почти на всех площадях в 2018 году был зафиксирован подрост ильма, практически отсутствовавший в описаниях 1954 года. Возросла встречаемость подроста клена, и если в 1954 году его плотность местами достигала 1300 экз./га, то в 2018 на многих участках уже доходила до 10000 экз./га. Также в подросте в 2018 году были отмечены адвентивные виды Juglans regia L. (на 27% пробных площадей), Acer negundo L. (на 12% пробных площадей), Phellodendron amurense Rupr. (на 6% пробных площадей).
Рис. 5.
Встречаемость видов в подросте, %.
Fig. 5. Constancy of species in saplings pool, %.
По оси абсцисс/X-axis: 1 – Quercus robur, 2 – Fraxinus excelsior, 3 – Tilia cordata, 4 – Acer platanoides, 5 – Ulmus glabra, 6 – Pyrus pyraster, 7 – Betula pendula.
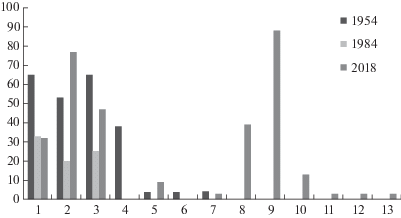
За 60 лет произошли изменения в составе подлеска (рис. 6). На протяжении этого срока в нем стабильно присутствуют типичные представители подлеска среднерусских дубрав Euonymus europaeus, E. verrucosus, Crataegus rhipidophylla. Уже к 1984 году из подлеска исчезли характерные для дубравы в 1954 г. светолюбивые ксеромезофиты Prunus spinosa L. и Caragana arborescens Lam. – адвентивный сибирский вид, используемый в озеленении с XVIII века. В 2018 г. активную позицию стали занимать теневыносливые мезо- и гигромезофиты Prunus padus L. и Sambucus nigra L. Стоит обратить внимание на появление в 2018 году в подлеске адвентивных видов Cornus sanguinea L., Ligustrum vulgare L., Morus nigra L. Также под пологом дубравы иногда встречается Partenocissus quinquefolia (L.) Planch., но не как лиана, а как почвопокровное растение.
Рис. 6.
Встречаемость видов в подлеске, %.
Fig. 6. Constancy of species in shrub layer, %.
1 – Euonymus verrucosus, 2 – Euonymus europaeus, 3 – Crataegus sp., 4 – Prunus spinosa, 5 – Lonicera tatarica L., 6 – Caragana arborescens, 7 – Rosa sp., 8 – Prunus padus, 9 – Sambucus nigra, 10 – Acer negundo, 11 – Ligustrum vulgare, 12 – Prunus cerasifera Ehrh., 13 – Morus nigra.
АНАЛИЗ СОСТОЯНИЯ НАПОЧВЕННОГО ПОКРОВА
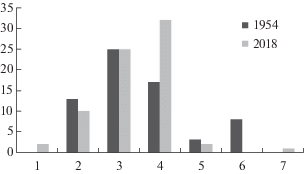
В геоботанических описаниях анализируемых лесных сообществ в 1954, 1984 и 2018 годах отмечено всего 99 видов трав: в 1954 году – 80 видов, в 1984 – всего лишь 34 вида, в 2018 году – 47 видов (табл. 1). Синхронно с видовым богатством менялась за эти годы видовая насыщенность – число видов трав на пробную площадь 400 м2: в 1954 году – в среднем 20 видов (от 13 до 34), в 1984 – всего лишь 7 видов (от 2 до 14), в 2018 году – 13 видов (от 4 до 20).
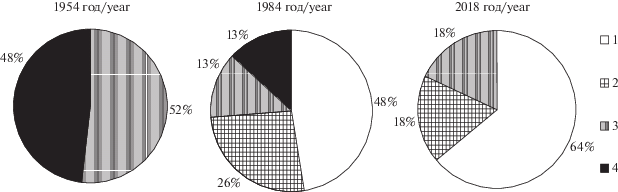
За исследованный чуть более чем 60-летний период степень развития травостоя в лесах обследованного участка заповедной дубравы снизилась. Распределение растительных сообществ по общему покрытию травостоя кардинально изменилось с 1954 по 1984 год. В середине XX века все леса имели травостой с покрытием более 30%, причем территории, занятые фитоценозами с травостоем покрытием от 30 до 50% и более 50% занимали примерно равную площадь. За 30 лет к 1984 году резко сократились площади фитоценозов с покрытием травостоя выше 50%, появились в большом количестве мертвопокровные (общее покрытие травостоя менее 15%) леса и леса с разреженным (от 15 до 30% общего покрытия) травостоем. Тенденция к увеличению числа участков с очень низким проективным покрытием травостоя и исчезновению лесов с высоким покрытием травостоя сохранилась в заповедной дубраве и в следующие десятилетия (рис. 7).
Рис. 7.
Соотношение долей участков с разной густотой травостоя в 8 квартале заповедной дубравы “Лес на Ворскле” в 1954, 1984 и 2018 годах.
1 – общее проективное покрытие травостоя (ОПП) <15%; 2 – ОПП от 15 до 30%; 3 – ОПП от 30 до 50%, 4 – ОПП > 50%.
Fig. 7. Ratio of plots with different density of herb layer in the reserved oak-wood “Les na Vorskle” (“Forest on the Vorskla River”) in 1954, 1984 and 2018.
1 – Total projective cover of herb layer (TPC) <15%; 2 – TPC 15 to 30%; 3 – TPC 30 to 50%, 4 – TPC > 50%.
Видовой состав и константность видов в напочвенном покрове дубравы “Лес на Ворскле” с 1954 по 2018 год значительно поменялись. Только 18 видов отмечены на геоботанических пробных площадях во все три срока описаний. Из них 7 видов не изменили свою константность: Adoxa moschatellina, Aegopodium podagraria, Galium odoratum, Geum urbanum, Glechoma hederacea, Pulmonaria obscura, Stachys sylvatica; все они во все 3 срока описаний имели константность ниже 30%, но более 5%.
Изменения коснулись не только видового состава травяного яруса леса, но и его экологического состава (табл. 2). Оценка по шкалам Элленберга усредненного травостоя дубравы показала существенное повышение теневыносливости и требовательности к влажности почвы с 1954 по 1984 годы, постепенное увеличение нитрофильности травостоя в течение всего периода наблюдений и отсутствие заметных изменений в отношении кислотности почвы (табл. 2). Такие сдвиги экологической структуры нижнего яруса растительности могут быть вызваны различными причинами. Рассмотрим более подробно, какие травы с встречаемостью не менее 5% появились или исчезли из дубравы с 1954 по 2018 годы. Виды трав, изменившие свою константность в дубраве за исследованный период, объединены в 6 групп: 1 – нередкие виды в 1954 году, но отсутствующие в описаниях конца XX – начала XXI века; 2 – нередкие виды в 1954 году, отмеченные в описаниях в 1984 году и отсутствующие в дубраве в 2018 году; 3 – присутствовали во все сроки описаний, но монотонно снижали константность в дубраве; 4 – виды, нередко отмечавшиеся в дубраве в 1954 и 2018 годах, но не встреченные ни разу в 1984 году; 5 – нередкие виды в описаниях 2018 года, отмеченные в описаниях в 1984 году и отсутствовавшие в дубраве в 1954 году; 6 – нередкие виды в описаниях 2018 года, отсутствовавшие в дубраве до конца XX века.
Таблица 2.
Оценка экологических условий в дубраве “Лес на Ворскле” по травостою в три срока описаний (шкала Элленберга, баллы) Table 2. Ecological conditions in the oak-wood “Les na Vorskle” (“Forest on the Vorskla River”) estimated by herb layer in three terms of descriptions (values of Ellenberg scales)
| Шкала (Элленберг) Scale (Ellenberg) |
1954 | 1984 | 2018 |
| Освещенность Light |
5.7 | 4.5 | 4.7 |
| Влажность Moisture |
4.8 | 5.4 | 5.7 |
| Кислотность Acidity |
6.4 | 6.4 | 6.7 |
| Богатство почвы азотом Soil richness with nitrogen |
5.1 | 6.4 | 7.0 |
В описаниях только 1954 года отмечено 44 вида (группа 1). Из них 26 видов имели константность более 5%, поэтому их присутствие в составе ценофлоры дубравы мы не можем считать случайностью, а 6 видов (Agrimonia eupatoria, Astragalus glycyphyllos, Clinopodium vulgare, Galium verum, Lysimachia nummularia и Torilis japonica) имели константность выше 30%. В целом эта группа показывает высокое светолюбие, низкую требовательность к содержанию доступного азота в почве, наиболее высокую устойчивость к засухе среди всех групп встречающихся в лесу видов (табл. 3). Также 4 вида с константностью выше 5%, в т.ч. 1 вид (Viola hirta) с константностью в описаниях 1954 года выше 30%, отсутствовали только в описаниях 2018 года (группа 2). Присутствовали в описаниях растительности во все годы наблюдений, но показали существенное монотонное снижение встречаемости 8 видов (группа 3), в т.ч. 6 видов с константностью в первый срок описаний более 30% (Campanula rapunculoides, C. trachelium, Carex pilosa, Dactylis glomerata, Fragaria vesca, Poa nemoralis). Эти группы (2 и 3) в целом экологически очень близки к группе 1 (табл. 3).
Таблица 3.
Экологические характеристики групп видов трав, различающихся константностью и присутствием в разные годы наблюдений (шкалы Элленберга, баллы) Table 3. Ecological features of herb groups differed by presence and constancy in different terms of observations (values of Ellenberg scales)
| Шкала Scale |
Группы видов, с дифференцированной встречаемостью по годам, годы встреч Species groups with differed constancy in years of descriptions, years when it was present |
|||||
|---|---|---|---|---|---|---|
| Группа 1 Group 1 1954 |
Группа 2 Group 2 1954 & 1984 |
Группа 3 убывание монотонное Group 3 monotonic decreasing |
Группа 4 Group 4 1954 & 2018 |
Группа 5 Group 5 1984 & 2018 |
Группа 6 Group 6 2018 |
|
| L | 6.7 | 6.7 | 5.3 | 6.0 | 4.0 | 5.1 |
| M | 4.0 | 4.3 | 5.1 | 5.1 | 6.0 | 5.5 |
| R | 7.1 | 7.0 | 6.2 | 6.6 | 6.8 | 6.5 |
| N | 4.2 | 4.3 | 5.4 | 7.1 | 6.8 | 6.8 |
Примечание. Шкалы Элленберга: L – освещенность, М – влажность, R – кислотность почвы, N – содержание доступного азота в почве. Название групп – годы описаний, в которых виды были отмечены, или значительное снижение встречаемости для видов, присутствовавших во все сроки описаний, нумерация групп соответствует тексту.
Note. Ellenberg scales: L – light, М – moisture, R – acidity, N – soil richness with available nitrogen. The group’s names are the years of descriptions when species were noted, or significant decrease of frequency for the species presented in all years of description; numbering of the groups corresponds to the text.
Встречены в описаниях 1954 и 2018 годов с константностью выше 5%, но отсутствовали в этих же сообществах в 1984 году 15 видов (группа 4), в том числе с константностью более 30% 4 вида (Carex spicata, Hypericum perforatum, Veronica chamaedrys, Viola mirabilis). Это группа видов выделяется высоким азотолюбием (табл. 3).
Только в 2018 году отмечено 9 видов (группа 6) с константностью выше 5%, в т.ч. 1 вид с константностью выше 30% (Moehringia trinervia). Также 4 вида с константностью выше 5%, в т.ч. 2 вида (Lamium maculatum, Polygonatum multiflorum) с константностью в 2018 году выше 30%, отсутствовали только в описаниях 1954 года (группа 5). Эти группы видов отличаются самой большой теневыносливостью и наибольшей требовательностью к влажности почвы среди всех групп видов с дифференцированной по годам встречаемостью (табл. 3).
МОДЕЛЬНЫЕ ПРОБНЫЕ ПЛОЩАДИ
Рассмотрим изменение растительности на примере модельных пробных площадей, заложенных в близких координатах в 1954, 1984 и 2018 годах (рис. 8).
Рис. 8.
Строение древостоя пробных площадей № 1; 2 в разные годы описаний.
Fig. 8. Structure of tree stand in sample plots 1 and 2 in different years of description.
Д – Quercus robur, Я – Fraxinus excelsior, Л – Tilia cordata, К – Acer platanoides, В – Ulmus glabra.
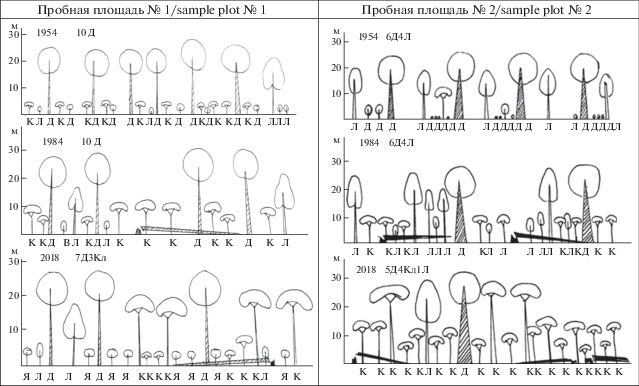
Пробная площадь № 1 (координаты 50.61182° с.ш.; 35.99615° в.д.) расположена в верхней части склона с крутизной 5–6° северо-западной экспозиции отвершка Монастырского яра. Микрорельеф выражен слабо, имеются небольшие западины и незначительные повышения возле комлей деревьев. Почва серая лесная слабо оподзоленная. Подстилающая порода – лессовидные суглинки. Гидрологический режим определяется атмосферными осадками и поверхностным стоком. В 1954 году древостой сомкнутостью 0.5 состоял из 70-летних дубов (формула древостоя 10Д) высотой до 28 м (рис. 9 ). На уровне второго яруса были зафиксированы отдельные липы высотой 20 м. В подросте произрастал дуб в возрастном диапазоне от 4 до 20 лет высотой до 2 м и плотностью 1000 экз./га. Аналогичные таксационные показатели были зафиксированы и для подроста липы. Более значительная плотность (3400 экз./га ) была у подроста клена, и высота некоторых особей достигала 3–3.5 м. Разреженный подлесок высотой от 0.5 м до 2 м суммарной плотностью 350 экз./га состоял из Euonymus verrucosus, Crataegus rhipidophylla и Prunus spinosa. Напочвенный покров равномерный с общим проективным покрытием 60–70% был образован 17 видами сосудистых растений, из них наибольшее обилие (20–30%) демонстрировали Carex pilosa, Lysimachia nummularia, Veronica сhamaedrys. В 1984 г. состав древесного полога не изменился (10Д), сомкнутость тоже осталась прежней (0.5), хотя часть дубов выпала, но кроны сохранившихся дубов разрослись. В подросте значительно усилилась роль клена. Его плотность достигла 10000 экз./га, преимущественно за счет молодых особей до 0.5 м высотой. Крупные особи 8–12 м сформировали разреженный полог III яруса, в этот же полог вошли отдельные особи липы. Появился в подросте ильм и ясень с плотностью 500 экз./га, но полностью исчез дуб. Опустел и ярус подлеска, а напочвенный покров стал представлять собой мертвопокровник из недоразложившегося листового опада дуба и клена, и лишь единичные особи Viola suavis и Lamium maculatum нарушали его монотонность (ОПП = 2%). В 2018 году дуб, не смотря на постепенное выпадание из древостоя в силу естественного старения, остался основной породой I яруса. Клен, формируя II ярус, упрочил свои позиции в древостое. Формула древостоя стала 7Д3Кл, сомкнутость возросла до 0.85. В подросте возросло участие ясеня (4500 экз./га), участие липы снизилось до 250, клена до 500 экз./га. В подлеске появилась черемуха и бузина. Напочвенный покров остался мертвопокровным (ОПП = = 5%), но видовое разнообразие увеличилось до 10 видов. Проективное покрытие более 1% показывали лишь Viola suavis, Asarum europaeum, Glechoma hederacea.
За 60 лет наблюдений в древостое постепенно ослабели позиции дуба, что связано с его выпадением и отсутствием возобновления. Позиции клена, наоборот, усилились. За счет формирования сомкнутого полога II яруса из клена почти в два раза возросла сомкнутость древостоя. Из подлеска исчезли светолюбивые ксеромезофитные виды, а появились теневыносливые гигромезофиты. Коэффициенты флористического сходства (Съеренсена) между описаниями разных лет очень низкие: KS/1954–1984 = 0; KS/1954–2018 = 7.4%; KS/1984–2018 = 16.7%.
Пробная площадь № 2 (координаты: 50.61279° с.ш.; 35.99155° в.д.) расположена на склоне крутизной 10° западной экспозиции основного русла Монастырского яра. Микрорельеф выражен слабо с вычленением небольших пристволовых повышений и пней от рубок во время Великой Отечественной войны. Почва серая лесная слабо оподзоленная. Подстилающая порода – лессовидные тяжелые суглинки. Гидрологический режим, как и на площади № 1, определяется атмосферными осадками и стоком воды по склону. В 1954 сомкнутость древостоя составила 0.4–0.6, он имел выраженное двухъярусное строение: в I ярусе могучие 300-летние дубы высотой 26–29 м, во II ярусе – более молодые липы высотой около 20 м. Формула древостоя 7Д3Л (рис. 9 ). На многих дубах было зафиксировано развитие трутовиков. Подрост был представлен дубом высотой до 6 м, плотностью 625 экз./га и липой до 3 м высотой и плотностью 875 экз./га. Подлесок составляли редкие 5-метровые деревья боярышника и местами густые заросли груши высотой 1 м. Напочвенный покров был разреженный, общее проективное покрытие едва достигало 30%. Его сформировали 22 вида сосудистых растений, наибольшее обилие (более 10%) было зафиксировано у Lysimachia nummularia и Poa angustifolia, несколько ниже (5–7%) – Torilis japonica и Astragalus glycyphyllos. В 1984 году лидирующие позиции в древостое сохранили 300–350-летние дубы, большая часть из них находилась в субсенильном и сенильном состоянии. Некоторые дубы выпали в результате старения. Их место в первом ярусе заняли подросшие липы. Формула древостоя 6Д4Л, средняя сомкнутость 0.4. В подросте в равных соотношениях были представлены клен и липа, с равной плотностью 1000 экз./га и высотой до подроста 3–6 м, но большая часть подроста приходилась на особи 0.6–0.8 м. Также в подросте были зафиксированы единичные особи вяза и груши. Подлесок отсутствовал. Общее проективное покрытие в напочвенном покрове, состоявшем лишь из 6 видов, снизилось до 24%. Отмечены следы пороев кабанов. Наибольшее обилие (20%) демонстрировала Aegopodium podagraria. В 2018 году в древостое снизилось участие дуба и липы, но возросла роль клена – 5Д3Кл2Л. Во втором ярусе уже господствует клен, в подросте – клен и ильм. Сомкнутость древесного полога достигла 0.8. В подлеске отмечены Sambucus nigra высотой до 2.5 м и мелкие (20–30 см) бересклеты: Euonymus europaeus и Euonymus verrucosus. Площадь пороев кабанов достигла 30%. Напочвенный покров образован 11 видами, его общее проективное покрытие составило 40%. Наибольший вклад в покрытие внесли Lamium maculatum (до 25%) и Glechoma hederacea (5%), остальные виды были единичны.
За 60 лет наблюдений в древостое, как и на пробной площади № 1, наблюдается интенсивное выпадение дуба и липы, но возрастает участие клена. В подлеске появилась бузина. За счет разрастания Lamium maculatum, отсутствовавшего в описаниях 1954 г. и лишь единично встреченного в 1984 г., до 40% увеличилось проективное покрытие в напочвенном покрове. Lamium maculatum – нитрофильный вид и его распространение возможно связано с увеличением в дубраве численности кабанов, чьи порои и экскременты послужили субстратом для его разрастания. Коэффициенты флористического сходства (Съеренсена) между описаниями разных лет низкие, хоть и выше чем на пробной площади № 1: KS/1954–1984 = 7.2%; KS/1954–2018 = 17.9%; KS/1984–2018 = 22.2%. В напочвенном покрове сохранился один малообильный общий вид – Stellaria holostea.
ОБСУЖДЕНИЕ
Массовое выпадение дуба в широколиственных лесах европейской лесостепи неоднократно отмечали исследователи на протяжении последней четверти ХХ века (Molchanov, 1975; Lositsky, 1981; Krasnitsky, Soshnin, 1986). Они объясняют это явление в основном погодными условиями, а именно периодическими засухами, провоцирующими вспышки насекомых-вредителей и патогенной микофлоры. В дубраве “Лес на Ворскле” выпадение дубов из древостоя и отсутствие его восполнения из подроста связано с достижением старшим поколением дуба возраста его средней продолжительности жизни в лесу – более 300 лет и невозможностью развития подроста Quercus robur под пологом тенистого древостоя из-за эколого-биологических свойств этого вида. Выпадение крупных деревьев дуба, а также крупных старых ясеней и лип, приводит к кратковременному осветлению под пологом дубравы, появлению ветровально-почвенных комплексов со сложной мозаикой экологических условий, активизации развития подроста и деревьев нижних ярусов, в первую очередь, клена, липы и ильма, что в конечном итоге увеличивает общую сомкнутость древостоя и тенистость леса. Это изменение экологических условий под пологом леса согласуется с полученными ранее данными для хвойно-широколиственных и южнотаежных лесов (Skvortsova et al., 1983).
Часть изменений в растительности лесостепной дубравы может быть связана с изменением климата. Ю.Н. Нешатаев и В.Н. Ухачева (Neshataev, Ukhacheva, 1998, 2001) указывают на мезофитизацию напочвенного покрова дубравы, объясняя ее потеплением зимнего сезона с одновременным увеличением количества осадков в летний период. На увеличение водообеспеченности в условиях нагорной дубравы за период с 1954 по 2018 годы указывает исчезновение опушечно-луговых и опушечно-степных ксеромезофитов среди трав и кустарников, появление гигромезофитов (Padus avium, Impatiens noli-tangere), присутствие в настоящее время в составе подроста и нижнего яруса древостоя ильма Ulmus glabra.
В годы Великой Отечественной войны произошло сильное изреживание древостоя. В середине XX века дубрава в 8 квартале “Леса на Ворскле” была весьма светлой, что позволяло под ее пологом встречаться даже некоторым лугово-степным видам (Galium verum, Origanum vulgare, Securigera varia и др.). Смыкание крон деревьев и развитие второго и третьего ярусов древостоя привело к тому, что в 1984 году нижние ярусы растительности (травостой и подлесок) оказались сильно подавлены и развиты очень слабо.
В послевоенное время почти до первого срока анализируемых описаний (1954 год) в этой части дубравы производился выпас скота. История выпаса скота на данном участке непродолжительная, т.к. до 1917 года это были частные охотничьи владения, с 1922 до 1951 года это был заповедник, правда, с зонированием режима использования и прерыванием заповедного режима во время Великой Отечественной войны (Shapovalov, 2005). При выпасе уничтожался подрост деревьев. В результате прекращения выпаса у подроста деревьев появилась возможность выхода в древостой, в ранее умеренно нарушенной дубраве развился третий ярус древостоя, создавший самое сильное затенение. Примерно с середины 1990-х годов практически никакого прямого влияния человека на лесные сообщества в пределах заповедника не стало. Прекращено выкашивание полян, вследствие чего они уже практически заросли лесом.
Во второй половине XX века значительно расширился ареал кабана в Восточной Европе. В 1950-х и 1960-х годах повсеместно увеличивалась численность кабана в Восточной Европе (Sludsky, 1956; Fadeev, 1998; Pavlov, 1996). В окрестностях заповедника первые за послереволюционное время кабаны появились в 1961 году. В сложные из-за социально-экономических перемен в стране 1990-е годы по материалам исследователей (Danilkin, 2002) началось снижение поголовья кабана на неохраняемых природных территориях по причине слабо контролируемой охоты и резкий рост численности в заповедниках, где звери оказались в безопасности. Такая же динамика наблюдалась в окрестностях участка “Лес на Ворскле” заповедника “Белогорье”. До середины 1990‑х годов по нашим наблюдениям их численность в заповедной дубраве “Лес на Ворскле” оставалась на невысоком уровне. С конца 1990-х годов плотность популяции кабанов резко возросла, достигая на максимумах 400 голов на территорию участка 1038 га (Tsarev, 2000; Chervonny, Shchekalo, 2010). Даже эпидемии и многоснежные зимы не снижали численность кабанов в заповеднике больше, чем на 1 год. В результате значительная территория дубравы превратилась в сплошные порои кабанов. Леса с очень низким проективным покрытием травостоя в описаниях 2018 года – это не мертвопокровные со сплошным слоем лесной подстилки, а беспокровные леса с обнаженной в результате деятельности кабанов минеральной почвой. Фактор высокой плотности популяции копытных отразился и в увеличении доли нитрофильных трав в травостое (особенно Lamium maculatum) и появлении нитрофильного кустарника (Sambucus nigra).
В последние годы в лесные сообщества дубравы “Лес на Ворскле” усилилось проникновение адвентивных, в т.ч. инвазионных видов. Поскольку обследуемая территория примыкает к старому дендрарию заповедника и поблизости располагаются посадки Robinia pseudoacacia L. и Juglans regia возрастом 50–90 лет, под пологом регулярно встречаются 1–3-летние сеянцы грецкого ореха, бархата амурского, изредка робинии, дуба красного. Нередок в возобновлении Fraxinus pennsylvanica Marshall. Регулярно внутри дубравы под пологом леса в качестве почвопокровного растения встречается Parthenocissus quinquefolium. В результате полного зарастания полян в составе лесных сообществ на их месте оказались Acer negundo и Prunus cerasifera, редко поселяющиеся под пологом леса (их проростки чаще всего гибнут в первый год жизни). Если Fraxinus pennsylvanica, Acer negundo, Prunus cerasifera, Parthenocissus quinquefolium, Quercus rubra L. отмечались под пологом дубравы, правда, в основном по просекам и около опушек на границе XX и XXI веков, то остальные указанные виды ранее под пологом дубравы не отмечались (Arbuzova, 2002). Начиная примерно с 2017 года, под пологом леса стал появляться Erigeron annuum, который очень обилен в составе лугов и рудеральных группировок окрестностей дубравы. Возможно, активизации проникновения адвентивных видов под полог дубравы в XXI веке способствовало массовое нарушение подстилки и напочвенного покрова кабанами.
ЗАКЛЮЧЕНИЕ
Таким образом, за 60 лет в заповедной лесостепной нагорной дубраве существенно изменилась структура древостоя, состав подроста и подлеска, структура, состав и видовое разнообразие травостоя. Описанные изменения носят необратимый характер, т.е. являются отражением идущей сукцессии. На естественную возрастную динамику леса, связанную со старением и отмиранием особей старшего поколения эдификатора – дуба, накладываются результаты влияния целого ряда факторов – изменения абиотических, биотических и антропогенных условий существования фитоценозов. Причинами сукцессии являются, по-видимому, следующие факторы:
– естественная смена лесов с резким преобладанием дуба на полидоминантные широколиственные леса с малым участием дуба, предопределенная эколого-биологическими особенностями возобновления древесных видов и достижением основным поколением дуба возраста средней продолжительности жизни,
– увеличение влажности климата,
– прекращение выпаса в послевоенные годы,
– появление и стабилизация на очень высоком уровне плотности популяции кабанов.
Список литературы
[Antipina, Maslov] Антипина Е.Е., Маслов С.П. 1994. Некоторые проблемы изучения истории взаимодействия хозяйственной деятельности человека с природными биоценозами. – Эволюционная и историческая антропоэкология. М. С. 111–120.
[Arbuzova] Арбузова М.В. 2002. Древесные интродуценты в лесных фитоценозах заповедника “Лес на Ворскле”. – Лесоведение. 4: 19–23.
[Bobrovsky] Бобровский М.В. 2010. Лесные почвы Европейской России: биотические и антропогенные факторы формирования. М. 359 с.
[Bulokhov, Semenishchenkov] Булохов А.Д., Семенищенков Ю.А. 2015. Типификация и коррекция синтаксонов лесной растительности южного Нечерноземья России и сопредельных регионов. – Бюллетень Брянского отделения РБО. 1 (5): 26–32.
[Chervonny, Schekalo] Червонный В.В., Щекало М.В. 2010. Население кабанов (Sus scrofa Linnaeus, 1758) в заповеднике “Белогорье” и Окском заповеднике. – Научные ведомости. Серия Естественные науки. 3 (74). Вып. 10: 61–69.
[Danilkin] Данилкин А.А. 2002. Свиные. М. 312 с.
[Fadeev] Фадеев Е.В. 1998. Размещение населения кабана в Волго-Камском крае во второй половине XX столетия. – Вестник Московского университета. Серия 16. Биология. 2: 22–26.
[Goryshina] Горышина Т.К. 1986. К истории заповедника “Лес на Ворскле”. – Комплексные исследования биогеоценозов лесостепных дубрав. Л. С. 3–18.
[Ipatov, Mirin] Ипатов В.С., Мирин Д.М. 2008. Описание фитоценоза: методические рекомендации. СПб. 71 с.
[Krasnitsky, Soshnin] Краснитский А.М., Сошнин Г.П. 1986. Характеристика некоторых сукцессионных процессов в дубравах Центрально-Черноземного заповедника. – Динамика биоты в экосистемах центральной лесостепи. М. С. 11–20.
[Küster] Кюстер Х. 2018. История леса. Взгляд из Германии. М. 304 с.
[Lositsky] Лосицкий К.Б. 1981. Дуб. М. 101 с.
[Molchanov] Молчанов А.А. 1975. Дубравы лесостепи в биогеоценологическом отношении. М. 234 с.
[Neshataev] Нешатаев Ю.Н. 1967. Опыт крупномасштабного геоботанического картирования лесостепной дубравы “Лес на Ворскле”. – Ученые записки ЛГУ. 331 (4). 50: 87–118.
[Neshataev, Ukhacheva] Нешатаев Ю.Н., Ухачева В.Н. 1998. Современное состояние и многолетняя динамика высокоствольной дубравы заповедника “Лес на Ворскле” (Белгородская область). – Проблема ботаники на рубеже XX–XXI веков. Материалы Х съезда РБО. СПб. С. 284–285.
[Neshataev, Ukhacheva] Нешатаев Ю.Н., Ухачева В.Н. 2001. Мониторинг растительности Среднерусской лесостепи. – Вестн. СПбГУ. 3 (2): 55–66.
[Pavlov] Павлов М.П. 1996. Кабан: почему он прижился в вятской тайге? – Охота и охотничье хозяйство. 7: 8–11.
[Ponomarenko et al.] Пономаренко Е.В., Сташенков Д.А., Кочкина А.Ф. 2015. Подходы к реконструкции заселения территории по почвенным признакам. – Поволжская археология. 1: 126–160.
[Ryabtsev et al.] Рябцев И.С., Тиходеева М.Ю., Рябцева И.М. 2009. Подпологовое возобновление лесообразующих пород в широколиственных лесах разного возраста с господством дуба черешчатого (Quercus robur L.). – Вестн. СПбГУ. 5 (2): 11–21.
[Semenishchenkov et al.] Семенищенков Ю.А., Агафонов В.А., Владимиров Д.Р., Казьмина Е.С. 2018. Мезофитные широколиственные леса союза Aceri campestris–Quercion в Воронежской области. – Вестник ВГУ, Серия: Химия. Биология. Фармация. 3: 98–105.
[Shapovalov] Шаповалов А.С. 2005. Истоки и организация заповедника “Лес на Ворскле” (“Белогорье”). – История заповедного дела: Материалы международной научной конференции. Борисовка. С. 9–11.
[Skvortsova et al.] Скворцова Е.Б., Уланова Н.Г., Басевич В.Ф. 1983. Экологическая роль ветровалов. М. 192 с.
[Sludsky] Слудский А.А. 1956. Кабан (морфология, экология, хозяйственное и эпизоотологическое значение, промысел). Алма-Ата. 220 с.
The Plant List. 30 Dec 2019. http://www.theplantlist.org
[Tsarev] Царев С.А. 2000. Кабан. Социальное и территориальное поведение. – Охотничьи животные России (биология, охрана, ресурсоведение, рациональное использование). Вып. 3. М. 114 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал