

Ботанический журнал, 2020, T. 105, № 8, стр. 794-806
ЦИКЛИЧНОСТЬ И МОРФОГЕНЕЗ ПОБЕГОВ PHLOMOIDES TUBEROSA (LAMIACEAE) В РАЗНЫХ УСЛОВИЯХ ПРОИЗРАСТАНИЯ НА ЮГЕ СИБИРИ
Е. К. Комаревцева *
Центральный сибирский ботанический сад СО РАН
630090 г. Новосибирск, ул. Золотодолинская 101, Россия
* E-mail: elizavetakomarevceva@yandex.ru
Поступила в редакцию 05.03.2020
После доработки 26.04.2020
Принята к публикации 28.04.2020
Аннотация
В статье рассматривается побегообразование у Phlomoides tuberosa (L.) Moench для выяснения целесообразности использования цикличности побега как одного из функциональных признаков. P. tuberosa – поликарпическое короткокорневищное растение с монокарпическими полурозеточными побегами разной цикличности, размножающееся эремами (семенами). Известно, что в европейской части России на заливных лугах у вида происходит развитие только дициклических полурозеточных побегов. Установлено, что в лугово-степных сообществах юга Сибири основная структурная единица особи P. tuberosa – моноциклический генеративный побег. Развитие его связано с ускорением темпов внутрипочечного развития полурозеточного побега, заключающимся в переходе весной инициальной зрелой почки в финальную, минуя стадию развития розеточного побега. Дициклический побег встречается у 20–30% цветущих особей, трициклические побеги единичны. Преобладание у P. tuberosa моноциклических побегов в условиях континентального (г. Новосибирск) и резко континентального (Горный Алтай) климата указывает на то, что для многолетних видов, размножающихся семенами, уменьшение цикличности монокарпического побега происходит при ухудшении условий. Это позволяет предполагать, что цикличность побегов P. tuberosa можно использовать как один из информативных косвенных признаков в функциональном анализе сообщества.
Для изучения и сравнения растительных сообществ в современной экологии растений используется функциональный подход, при котором изучаются такие морфологические признаки (функциональные), изменения которых наиболее наглядно выражают реакцию видов на факторы окружающей среды, и, следовательно, отражают их экологическую стратегию (Chapin et al., 1996; Weiher et al., 1999; Cornelissen et al., 2003; McIntyre, 2006). Наиболее популярна система признаков LHS, предложенная М. Westoby (1998), включающая удельную площадь листа (L), высоту растения (H) и массу семян (S). Вместе с тем идет активный поиск новых функциональных признаков, коррелирующих с LHS-признаками (Chave et al., 2009; Mommer, Weemstra, 2012; Klimešova et al., 2016; 2017; Vojtkó et al., 2017). Так в работе J. Klimešova с соавторами (Klimešova et al., 2016) рассматривается продолжительность жизни особи и цикличность побегов для разных видов в качестве перспективных косвенных параметров, влияющих на функциональные признаки сообщества. Авторами проанализирована встречаемость моноциклических и полициклических побегов у видов, входящих в состав 32 типов сообществ Центральной Европы. Было выяснено, что моноциклические побеги характерны для быстрорастущих видов, находящихся в благоприятных местообитаниях (прибрежная растительность, широколиственные леса и литораль), растения с ди- и полициклическими побегами приурочены к болотам, пустошам, хвойным лесам и альпийским лугам. В ходе эколого-биологических исследований растительности географических зон территории России выявлены закономерности распределения видов разных жизненных форм и экологических групп в растительных зонах, которые свидетельствуют о более сложной зависимости биоморфологических особенностей растения от условий произрастания (Serebryakov, 1959; 1964; Belostokov, 1962; Golubev, 1965; Gorshkova, 1966, 1975; Polyntseva, 1971; Fedoskin, 1974). В частности на массовом материале установлено, что при движении с юга на север происходит закономерное уменьшение числа видов с моноциклическими побегами: от 36% в полупустынях Казахстана до 5% в арктической тундре в низовьях р. Пясины на Приполярном Урале. И, следовательно, необходимость быстрого роста растения через развитие моноциклических побегов возникает в условиях обилия тепла и света, но дефицита влаги в почве (Serebryakov, 1959). На распространение видов с такими побегами влияет и длительность вегетационного периода. По сравнению с равнинными степями европейской части России, в степях Забайкалья преобладают виды с полициклическими побегами (56.4–81.2%), что обусловлено сокращением вегетационного периода в резко континентальном климате (Gorshkova, 1966).
Цикличность побегов – это видовая характеристика. Есть виды с постоянной цикличностью монокарпических побегов в пределах одной особи, и виды с изменяющейся цикличностью побегов (Serebryakov, 1952; 1959). Следовательно, для корректного использования цикличности побега в экологических исследованиях необходимо детальное изучение побегообразования у генеративной особи, особенно если вид характеризуется формированием побегов разной цикличности. Одним из таких видов является Phlomoides tuberosa (L.) Moench (Lamiaceae).
P. tuberosa – евроазиатский вид. На западе своего ареала он встречается в Средней Европе, Средиземноморье, на Кавказе, в Малой Азии. В России проходит северная граница (60–64° с.ш.) его ареала: от европейской части, через Западную и Восточную Сибирь, до Уссурийского края. Это лугово-степной вид, произрастающий на остепненных лугах, луговых степях, в смешанных, разреженных лесах, на их опушках, изредка залежах (Doronkin, 1997). В Алтае-Саянской горной области P. tuberosа – постоянный вид суходольных лугов, луговых и разнотравно-дерновиннозлаковых степей (Makunina, 2016). В горах он обильно и умеренно встречается в верхне- и среднегорном поясах и в малом обилии – в субальпийском поясе, нижнегорной и предгорной зонах. Вид является ксеромезофитом: массово произрастает на сухих лугах, умеренно распространен по сухим, средним и влажным степям, единично – во влажных и сырых лугах (Tsatsenkin, 1967).
В Западном Казахстане (окр. с. Жаныбек) в полупустынных и степных сообществах у особей P. tuberosa формируются моно-, ди- и трициклические генеративные побеги (Belostokov, 1962; Serebryakov 1959). В Калужской области на заливных лугах по берегам р. Угра Н.М. Григорьева отмечает развитие ди- и полициклических полурозеточных побегов у генеративной особи (Grigorieva, 1994). Ранее нами было установлено, что на территории Горного Алтая, Хакасии и Тувы для P. tuberosa характерно развитие преимущественно моноциклических полурозеточных побегов (Komarevtseva et al., 2018). Дициклические побеги встречаются значительно реже, появление полициклических полурозеточных побегов единично. В генеративном периоде отмечены перерывы в цветении, связанные не с цикличностью генеративного побега, а с появлением вегетативных розеточных побегов, не переходящих к цветению. Для объяснения изменения цикличности побегов этого вида в разных регионах необходимо детальное исследование формирования побегов.
Цель работы заключалась в анализе цикличности полурозеточных побегов P. tuberosa в градиенте эколого-ценотических условий. Для этого проводилось изучение побегообразования и структуры генеративных побегов и сопоставление особенностей развития P. tuberosa в Новосибирской области, Горном Алтае в сравнении с литературными данными по европейской части России.
МАТЕРИАЛЫ И МЕТОДЫ
Сборы генеративных особей проводились в конце июня в окрестностях с. Усть-Кан (Горный Алтай) в двух ценопопуляциях (ЦП 1, 2) и в лесопарковой зоне Центрального сибирского ботанического сада (ЦСБС) (г. Новосибирск) (ЦП 3). Для изучения внутрипочечного развития сбор материала проводился только в ЦП 3 в несколько сроков: в конце апреля, начале июня, конце июля и в начале октября. В каждом местообитании были изучены по 30–45 цветущих особей. Для оценки мощности развития измерялись длина генеративного побега и его паракладиев, определялась масса побега и масса частей побега, несущих генеративные органы (на главной оси и на паракладиях) и число цветков в одном пазушном дихазии на главном соцветии и репродуктивное усилие особи. Все полученные данные статистически обработаны в пакете программ Exсel. Достоверность различий между параметрами определялось по по t-критерию Стьюдента при 5%-м уровне значимости (Zaitsev, 1991).
ЦП 1 входит в состав злаково-разнотравной луговой степи на опушке лиственничника по северо-западному склону (уклон 5°). Общее покрытие (ОПП) – 70%. Проективное покрытие (ПП) P. tuberosa – 8%. ПП злаков составляет 10%: Poa angustifolia (5%), Koeleria cristata (3%), Poa attenuata (3%). Разнотравье представлено Potentilla chrysantha (5%), Galatella biflora (5%), Ligularia glauca (5%), Fragaria viridis (5%), Adonis vernalis (3%) и др. ЦП 2 расположена в нижней части юго-восточного склона (крутизна 30°) в составе каменистой закустаренной луговой степи. ОПП составляет 80%. ПП P. tuberosa – 3–5%. Из кустарников присутствуют Spirea trilobata (8%), Caragana pygmaea (1%). ПП злаков составляет 15%: Phleum phleoides (5%), Poa botrioides (5%), Koeleria cristata (3%), Agropyron pectinatum (3%). Из разнотравья произрастают: Astragalus austriacus (5%), Onobrychis arenaria (3%), Artemisia frigida (3%), Artemisia commutata (3%), Bupleurum bicaule, B. multinerve, Dianthus versicolor, Thymus mongolicus и др. ЦП 3 входит в состав разнотравно-злакового луга, расположенного на северо-западном склоне (уклон 12°). ОПП травостоя – 80%. ПП P. tuberosa – 2%. ПП злаков – 60%, в том числе Dactylis glomerata (30%), Festuca pratensis (20%), Phleum pretense (5%), Poa angustifolia (5%). Из разнотравья присутствуют Fragaria viridis (20%), Lathyrus pratensis (10%), Heiracium umbellatum (5%), Trifolium pratense (5%), Vicia amoena (5%), Viola hirta (2%) и др. виды. Заметное покрытие имеют индикаторы антропогенного воздействия: Cirsium setosum (10%), Plantago major (3%) и Urtica dioica (3%).
Функциональное зонирование побеговой системы определялось согласно W. Troll (1964), И.В. Борисовой, Т.А. Поповой (Borisova, Popova, 1990), тип соцветия – по Т.В. Кузнецовой и А.К. Тимонину (Kuznetsova, Timonin, 2017). Морфогенез побега описан по представлениям Т.И. Серебряковой (Serebryakovа, 1971). Репродуктивное усилие вычислялось как доля веса репродуктивных органов от общего веса особи в процентах (Markov, Pleshchinskaya, 1987).
Развитие почки анализировалось по Т.И. Серебряковой (Serebryakovа, 1983) с дополнениями Т.А. Комаровой (Komarova, 1989). В ходе развития побега различаются три типа почек: 1. инициальная – от заложения первых листовых зачатков до их развертывания; 2. промежуточная – верхушечная почка розеточного побега до заложения в ней генеративных зачатков; 3. финальная почка, содержащая все метамеры будущего генеративного побега, включая его флоральную часть (Serebryakovа, 1983). В связи с принадлежностью P. tuberosa к группе видов с симподиальным типом нарастания корневища применяется подразделение основных типов почек (Komarova, 1989). Среди инициальных почек выделены начальная (I1), развивающаяся в материнской почке (почка в почке), и зрелая инициальная почка (I2), освободившаяся от покровов материнской почки. Промежуточные почки P. tuberosa подразделяются на вегетативные (М1) (до образования кроющих листьев соцветий) и генеративные (М2) (от заложения первых генеративных органов до формирования финальной почки). В финальных почках различаются: незрелая (F1) – все метамеры побега заложены, но развитие цветочных зачатков не закончено; зрелая (F2) – полностью сформированная почка.
РЕЗУЛЬТАТЫ
P. tuberosa – поликарпическое короткокорневищное растение с полурозеточными побегами. В подземной части растения развивается короткое симподиально нарастающее корневище, состоящее из последовательно сменяющихся базальных частей побегов (резидов), с многочисленными длинными придаточными корнями, на концах которых образуются шаровидные клубни. Размножение растений этого вида происходит только семенным способом.
Структура моно- и дициклического генеративных побегов Р. tuberosa представлена на рис. 1. Базальная часть полурозеточных побегов несет чешуевидные и розеточные ассимилирующие листья. На самых первых 2–3 метамерах развиваются чешуевидные листья (катафиллы), далее – переходные листья (разросшиеся основания зачаточных листьев с редуцированной листовой пластинкой), затем розеточные листья c нормально развитой пластинкой.
Рис. 1.
Структурно-функциональные зоны генеративных побегов Phlomoides tuberosa: а – моноциклического, в – дициклического.
НЗТ – нижняя зона торможения, ЗВ – зона возобновления, СЗТ – средняя зона торможения, ЗО – зона обогащения, ЗГС – зона главного соцветия; 1 – отмерший розеточный лист; 2 – остаток прошлогоднего генеративного побега (резид); 3а – спящая почка; 3b – почка возобновления; 4 – розеточный лист; 5 – паракладий; 6 – главное соцветие; 7 – многочленный дихазий; 8 – верхушечные недоразвитые дихазии главного соцветия; 9 – уровень почвы; 10 – первый годичный прирост дициклического побега.
Fig. 1. Structural and functional zones of generative shoots of Phlomoides tuberosa: a – monocyclic, b – dicyclic.
НЗТ (LIZ) – lower inhibition zone, ЗВ (IZ) – innovation zone, СЗТ (MIZ) – middle inhibition zone, ЗО (AZ) – amplification zone, ЗГС (MI) – main inflorescence; 1 – dead rosette leaf, 2 – remnant of the previous year’s generative shoot; 3а – dormant bud; 3b – innovation bud; 4 – rosette leaf; 5 – paracladium; 6 – main inflorescence; 7 – dichasium; 8 – apical underdeveloped dichasia of the main inflorescence; 9 – soil level; 10 – first annual growth of a dicyclic shoot.
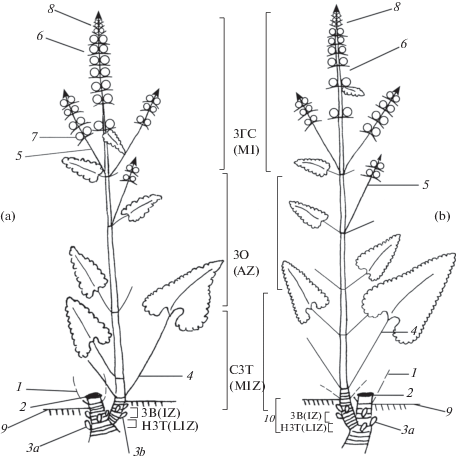
Розеточные листья длинночерешковые, с треугольной листовой пластинкой и глубоко сердцевидным основанием. На удлиненной части полурозеточного побега располагаются листья, форма которых меняется снизу-вверх от треугольной до яйцевидно-ланцетной с сердцевидным основанием и городчатым краем. В верхней части побега развивается полителическая синфлоресценция. Флоральная единица – открытый колосовидный тирс, боковые оси его представлены 3–4-членными дихазиями (парциальные флоресценции). Тирс главной оси (главная флоресценция) состоит из большего числа метамеров (до 9) по сравнению с паракладиями – боковыми осями (до 6). Паракладии (кофлоресценции) развиваются сразу под главной флоресценцией на 1–3 метамерах. На всем протяжении полурозеточного побега в пазухах всех листьев располагаются почки. В побегообразовании участвуют почки, располагающиеся в подземной части побегов: 1. почки возобновления в базальной части побега текущего года; 2. спящие почки в основании прошлогодних побегов, находящиеся в покое в течение нескольких лет и только потом прорастающие или отмирающие.
В моно- и дициклических генеративных побегах Р. tuberosa выделяются следующие функциональные зоны (рис. 1). В базальной части побега, находящейся в приземном слое, располагаются нижняя зона торможения, зона возобновления и начинается средняя зона торможения. В надземной удлиненной части выделяются средняя зона торможения, зона обогащения с паракладиями. В верхней части побега располагается главная флоресценция – зона главного соцветия. Нижняя зона торможения характеризуется слаборазвитыми почками (1–2 пары зачатков) в пазухах чешуевидных листьев. В зоне возобновления образуется 2–5 крупных почек возобновления (до 11 пар зачатков) (рис. 1a; табл. 1). Средняя зона торможения состоит из 2–4 коротких метамеров с переходными и розеточными листьями, а также 1–4 длинных метамеров со стеблевыми черешковыми листьями. Зона обогащения представлена 1–4 (до 6) паракладиями, состоящими из 2–4 длинных метамеров с пазушными цветками. При отсутствии паракладиев эта часть побега включается в среднюю зону торможения. Зона главного соцветия – открытый тирс из 2–9 метамеров, на которых в пазухах сидячих листьев развиваются цветки, составляющие многочленные дихазии. Побег заканчивается 3–6 сближенными метамерами с заложенными, но не достигающими полного развития, элементами соцветия, которые впоследствии засыхают. В результате моноциклический полурозеточный побег состоит в базальной части из 5–14 коротких метамеров с чешуевидными и розеточными листьями, в надземной части из 1–4 длинных метамеров со стеблевыми черешковыми листьями и 2–9 метамеров с развитыми парциальными соцветиями в пазухах сидячих листьев. В исследуемых местообитаниях у цветущих особей моноциклические генеративные побеги составляют 75–80%. Их развитие чаще происходит из спящих почек (70–75% от общего числа моноциклических побегов), только 25–30% моноциклических побегов развиваются из почек возобновления. Это определяет высокую вариабельность числа метамеров нижней зоны торможения у моноциклических побегов – от 2 до 7 метамеров в зависимости от типа материнской почки.
Таблица 1.
Зональная структура моноциклического полурозеточного побега Phlomoides tuberosa Table 1. Zonal structure of the monocyclic semirosette shoot of Phlomoides tuberosa
| Зоны Zones |
Пары листьев Leaf pairs |
||||
|---|---|---|---|---|---|
| Чешуевидные Cataphylls |
Розеточные Rosette |
Стеблевые
с черешком Petiolate cauline |
Стеблевые сидячие
с пазушными дихазиями Sessile cauline with axillary dichasia |
Верхушечные зачаточные
с недоразвитыми дихазиями ЗГС Apical primordial with underdeveloped dichasia of MI |
|
| НЗТ (LIZ) | 2–7 | ||||
| ЗВ (IZ) | 1–3 | ||||
| СЗТ (MIZ) | 1–2 | 1–2 | 1–4 | ||
| ЗО (AZ) | – | – | 1–3 | ||
| ЗГС (MI) | – | – | – | 2–9 | 3–6 |
Примечание. НЗТ – нижняя зона торможения, ЗВ – зона возобновления, СЗТ – средняя зона торможения, ЗО – зона обогащения, ЗГС – зона главного соцветия.
Note. НЗТ (LIZ) – lower inhibition zone, ЗВ (IZ) – innovation zone, СЗТ (MIZ) – middle inhibition zone, ЗО (AZ) – amplification zone, ЗГС (MI) – main inflorescence.
Для структуры дициклического побега характерна такая же последовательность зон, как и у моноциклического побега, несмотря на то, что он состоит из 2-х годичных приростов (рис. 1b; табл. 2). В его базальной части уменьшается протяженность нижней зоны торможения (1–3 метамера), в средней зоне торможения увеличивается число коротких метамеров за счет удлинения розеточной фазы развития побега: 5–10 коротких и 1–4 длинных метамера. Зона обогащения: 1–2 метамера с 1–4 паракладиями из 2–4-х метамеров. При отсутствии паракладиев эта часть побега замещается средней зоной торможения. В зоне главного соцветия число метамеров с развитыми цветками изменяется в меньшей степени (3–6 метамеров), число верхушечных сближенных метамеров с недосфомировавшимися цветками составляет 3–4 метамера. В итоге число коротких и длинных метамеров у дициклического побега сопоставимо с числом этих метамеров у моноциклического. В базальной части дициклического побега насчитывается 7–16 коротких метамеров, в надземной части – 1–4 метамера с черешковыми листьями, число метамеров главного соцветия варьирует с 3 до 6, что заметно меньше по сравнению с моноциклическим побегом. Встречаемость дициклического побега составляет 20–30%, и в основном он развивается из почек возобновления (70–90% от общего числа побегов этого типа), возможно развитие и из спящих почек (10–30% побегов).
Таблица 2.
Зональная структура дициклического побега Phlomoides tuberosa Table 2. Zonal structure of the dicyclic semirosette shoot of Phlomoides tuberosa
| Зоны Zones |
Пары листьев Leaf pairs |
||||||
|---|---|---|---|---|---|---|---|
| 1-й год развития 1st year of development |
2-й год развития 2nd year of development |
||||||
| Чешуе-видные Cataphylls |
Розеточ-ные Rosette |
Чешуе-видные Cataphylls |
Розеточ-ные Rosette |
Стеблевые
с черешком Petiolate cauline |
Стеблевые сидя-чие с пазушными дихазиями Sessile cauline with axillary dichasia |
Верхушечные зача-точные с недоразви-тыми дихазиями ЗГС Apical primordial with underdeveloped dichasia of MI |
|
| НЗТ (LIZ) | 1–3 | ||||||
| ЗВ (IZ) | 1–3 | ||||||
| СЗТ (MIZ) | 1–2 | 1–2 | 2–4 | 1–2 | 1–4 | ||
| ЗО (AZ) | – | – | – | – | 1–2 | ||
| ЗГС (MI) | – | – | – | – | – | 3–6 | 3–4 |
Примечание. НЗТ – нижняя зона торможения, ЗВ – зона возобновления, СЗТ – средняя зона торможения, ЗО – зона обогащения, ЗГС – зона главного соцветия.
Note. НЗТ (LIZ) – lower inhibition zone, ЗВ (IZ) – innovation zone, СЗТ (MIZ) – middle inhibition zone, ЗО (AZ) – amplification zone, ЗГС (MI) – main inflorescence.
Моно- и дициклические побеги у растений, произрастающих в одном местообитании, достоверно не отличаются между собой по основным биоморфологическим параметрам (табл. 3). Мощность развития всех типов полурозеточных побегов определяется только условиями среды. Достоверно отличаются особи из степных сообществ (ЦП 1, 2) от растений на разнотравно-злаковом лугу (ЦП 3) по длине генеративного побега, его массе и массе репродуктивной части побега. Побеги Р. tuberosa в степных сообществах более чем в 3 раза уступают по своей длине и массе побегам из более мезофитных условий разнотравно-злакового луга. Несмотря на заметное различие значений других параметров (длина паракладиев, число цветков в дихазии на главном соцветии, репродуктивное усилие) достоверных отличий между побегами особей из разных ценопопуляций не обнаружено.
Таблица 3.
Основные параметры моно- и дициклических побегов Phlomoides tuberosa из разных местообитаний Table 3. Main traits of mono- and dicyclic shoots of Phlomoides tuberosa from different habitats
| Признаки Traits |
Цикличность побегов Shoot cyclicity |
|||||
|---|---|---|---|---|---|---|
| ЦП (CP) 1 | ЦП (CP) 2 | ЦП (CP) 3 | ||||
| моно- mono- |
ди- di- |
моно- mono- |
ди- di- |
моно- mono- |
ди- di- |
|
| Длина побега, см Shoot length, cm |
32.7* 16–67.5** |
39.6 27.5– 57 |
39.1 18–66.5 |
37.7 29.5–52 |
120.1 73.5–167 |
139.9 100–210 |
| Надземная масса, г Aboveground weight, g |
5.5 2.3–11 |
4.4 2.6–6.2 |
6.2 2.1–14.4 |
4.4 2–12 |
17.5 5.3–44.3 |
25.3 10.7–55.8 |
| Масса репродуктивной части побега, г Weight of the reproductive part of shoot, g |
1.9 0.4–5 |
1.4 0.4–2.8 |
1.8 0.1–6.3 |
2 0.6–6.6 |
6.4 1.3–22.6 |
8.5 1.9–31.2 |
| Длина паракладиев, см Paracladium length, cm |
16.5 1.2–33.5 |
12.1 7–20.5 |
13.8 3.5–25.5 |
15.9 4.5–35.5 |
32.9 9.8–64.5 |
30.9 14–65 |
| РУ (RE) | 34.9 12.6–64 |
31 16.8–44.9 |
25.7 4.4–49 |
41.3 22.2–55 |
14.1 2–26.8 |
17.2 8.6–25 |
| Число цветков в дихазии на главном соцветии, шт. Number of flowers in dichasia on the main inflorescence, pcs. |
10.9 5–18 |
15.3 11–20 |
12.8 4–21 |
10.4 7–15 |
14.6 8–27 |
16.9 8–24 |
Условия среды оказывают влияние не только на морфологические параметры разных типов побегов, но и на соотношение числа розеточных, стеблевых черешковых листьев и развитие паракладиев. В ксерофитных сообществах (ЦП 1, 2) на побегах развиваются в среднем 2(1–4) пары розеточных листьев и 2(1–3) пары стеблевых, на лугу – 1(до 2) пара розеточных и 3(2–4) пары стеблевых листьев. В степных сообществах (ЦП 1, 2) у более половины особей формируется только главная флоресценция и длина имеющихся паракладиев не более 35 см. В разнотравно-злаковом лугу (ЦП 3) практически все цветущие особи имеют развитые синфлоресценции, в которых число паракладиев доходит до 6, а их длина – свыше 60 см. У моноциклических побегов зона главного соцветия более чувствительна к условиям среды: в степных сообществах она состоит из 2–7 метамеров, в более мезофитных условиях (ЦП 3) увеличивается до 4–9. У дициклических побегов число метамеров в главном соцветии постоянно во всех изученных ценопопуляциях – 3–6 (в среднем 4) метамеров.
Таким образом, структура моно- и дициклических побегов характеризуются одинаковой последовательностью функционально-структурных зон. Зона возобновления у обоих типов побегов располагается на первом годичном приросте. Побеги сопоставимы по числу коротких метамеров в базальной части и длинных метамеров в надземной части побега за исключением генеративной сферы побега: у моноциклического побега она характеризуется большей изменчивостью по сравнению с дициклическим. На степень развития всех типов полурозеточного побега большое воздействие оказывают эколого-ценотические условия. Во всех исследованных местообитаниях у цветущих особей преобладают моноциклические генеративные побеги, развивающиеся в основном из спящих почек. Дициклические побеги чаще развиваются из почек возобновления.
Генеративный период P. tuberosa характеризуется перерывами в цветении. Для выяснения причины этого явления изучено изменение емкости почки возобновления в процессе ее роста. Развитие почки происходит весной и летом (Serebryakovа, 1956). Весной в почке закладывается генеративная сфера будущего побега, что отмечают Г.П. Белостоков (Belostokov, 1962) для Западного Казахстана и Н.М. Григорьева (Grigorieva, 1994) для Калужской области. Наши исследования подтверждают это для юга Сибири.
Внутрипочечный период у побега P. tuberosa двухгодичный, в ходе его формируются инициальные почки I1 и I2 (рис. 2). Внепочечный период длится один год с развитием моноциклического генеративного побега или два года при развертывании дициклического полурозеточного побега. В последнем случае в первый год на верхушке розеточного побега развивается промежуточная вегетативная почка (М1), из которой весной второго года последовательно формируются промежуточная генеративная почка (М2), затем финальная почка (F2), дающая начало удлиненной части дициклического полурозеточного побега. Побегообразование из спящей почки проходит по такой же схеме, но с увеличением емкости почек за счет нижних метамеров (с чешуевидными листьями).
Рис. 2.
Морфогенез побегов Phlomoides tuberosa разной цикличности.
I1, I2, M1, M2, F2 – типы почек; 1 – недифференцированные зачатки листьев; 2 – чешуевидные листья; 3 – зачатки переходных листьев; 4 – зачатки с листовыми пластинками; 5 – зачатки стеблевых листьев; 6 – зачатки дихазий; 7 – конус нарастания; 8 – зеленый розеточный лист; 9 – отмерший розеточный лист; 10 – метамеры.
Fig. 2. Morphogenesis of Phlomoides tuberosa shoots of different cyclicity.
I1, I2, M1, M2, F2 – types of buds; 1 – undifferentiated leaf primordia; 2– cataphylls; 3 – primordia of transitional leaves; 4 – primordia with leaf blades; 5 – primordia of cauline leaves; 6 – primordia of dichasia; 7 – vegetative cone; 8 – green rosette leaf; 9 – dead rosette leaf; 10 – metamers.
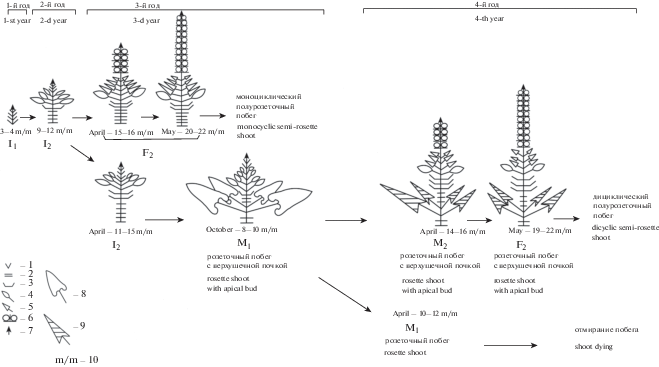
Инициальная начальная почка (I1) развивается в почке возобновления (I2) цветущего побега и в конце июля состоит из 3–4 пар недифференцированных зачатков. Во 2-й вегетационный сезон из материнской почки развивается побег, у которого в зоне возобновления из I1-почек формируются 1–3 (5) крупных инициальных зрелых почек (I2), емкость которых в октябре составляет 9–12 метамеров (4–6 пар зачатков чешуевидных и переходных листьев, 3 пары зачатков с листовыми пластинками и 2–3 пары недифференцированных зачатков).
На третий год, как правило, только одна из почек возобновления (I2) получает дальнейшее развитие. Остальные почки в зоне возобновления замедляют свое развитие, нарастая в течение года только на 2–3 метамера, и переходят в разряд спящих почек, сохраняющих свою жизнеспособность до 3-х лет. Развитие I2-почки может пойти двумя путями с формированием розеточного или полурозеточного побегов (рис. 2). При заложении в ней генеративной сферы образуется финальная почка (F2). Формирование ее растянуто во времени. В конце апреля закладывается 5–6 новых метамеров, из которых 4 верхних несут зачаточные соцветия, в 1-й декаде мая закладывается еще 5–6 метамеров только с генеративными зачатками. Среди зачатков с листовыми пластинками различаются зачатки розеточных (широкая пластинка на длинном черешке) и стеблевых (узкая пластинка на коротком черешке) листьев. В результате емкость зрелой финальной F2-почки составляет 20–22 метамера: 7 пар чешуевидных и переходных, 1 пара зачатков розеточных, 3–4 пары зачатков стеблевых листьев и 9–10 метамеров с цветочными зачатками. Из этой почки в середине мая начинается отрастание полурозеточного генеративного побега, зацветающего в конце июня. Но не из всех генеративных зачатков развиваются цветки, на последних 3–4 метамерах они остаются в зачаточном состоянии и засыхают. Число засохших метамеров на верхушке генеративных побегов в степных сообществах увеличивается до 6.
Не всегда почка возобновления преобразуется в финальную. В этом случае емкость I2-почки на конец апреля увеличивается только на 2–4 метамера и составляет 11–16 метамеров: 6–9 пар чешуевидных и переходных, 3–4 пары зачатков с листовой пластинкой и 2–3 пары недифференцированных зачатков. Из нее развивается розеточный побег с 1–2 (до 3) парами зеленых листьев, ниже которых в пазухе верхних чешуевидных листьев формируются почки возобновления (I2) следующего порядка. В конце сезона зеленые листья на побеге отмирают, но сохраняется верхушечная почка розеточного побега (M1). Ее емкость на начало октября составляет 8–10 метамеров: 2–4 пары чешуевидных, 3 пары листовых зачатков и 3 пары недифференцированных зачатков.
На четвертый год отмечены два варианта дальнейшего развития розеточного побега, что обусловлено заложением генеративных зачатков в его апексе или их отсутствием (рис. 2). При появлении в верхушечной почке новых 5–6 метамеров (4-х верхних с генеративными зачатками) формируется промежуточная генеративная почка (М2). Емкость ее в конце апреля составляет 14–16 метамеров: 6–7 пар чешуевидных, 1 пара розеточных листьев, 3–4 пары стеблевых и 4 пары верхушечных листьев с пазушными соцветиями. В 1-й декаде мая в почке закладывается еще 5–6 метамеров с генеративными зачатками. Формируется зрелая финальная F2-почка, емкость которой составляет 19–22 метамеров: догенеративная часть аналогична с таковой у М2-почки, а генеративная представлена 9–10 верхними метамерами с зачатками пазушных соцветий. До 4-х последних пар соцветий остаются в зачаточном состоянии и засыхают, в степных сообществах число засохших метамеров доходит до 5. Из такой почки развертывается дициклический полурозеточный побег, зацветающий в конце июня.
Если заложение генеративной сферы в верхушечной почке розеточного побега не происходит, верхушечная почка (М1) весной увеличивает свою емкость незначительно (до 10–12 метамеров). В единичных случаях розеточный побег продолжает вегетацию, развертывая розеточные листья. На следующий (5-й) год при закладке весной в его верхушечной почке флоральной части формируется трициклический полурозеточный побег. Но, как правило, отрастания розеточных листьев на второй сезон не происходит. В его основании сохраняются 1–2 зрелых инициальных почек, из которых возможно развитие новых побегов.
Следовательно, перерывы в цветении генеративной особи связаны с тем, сформируется ли весной генеративная сфера в зрелых инициальных и промежуточных почках или нет. Если этого не происходит, то из инициальной почки развивается розеточный побег, а однолетний розеточный побег с промежуточной вегетативной почкой на второй год чаще всего отмирает.
ОБСУЖДЕНИЕ
Развитие полурозеточного побега P. tuberosa проходит по-разному в разных частях ареала. На заливных лугах Калужской области для генеративной особи характерно развитие из почки возобновления дициклического полурозеточного побега с последовательным прохождением всех фаз развития – от инициальной почки через развитие розеточного побега с верхушечной промежуточной вегетативной, затем и генеративной почкой, до формирования удлиненной части генеративного побега (Grigorieva, 1994). Большая часть почек в зоне возобновления побегов развивается нормальными темпами и не переходит в разряд спящих, они прорастают с развитием розеточного побега в 1-й год и на следующий год – с образованием полурозеточного. Это подтверждается данными Н.М. Григорьевой об одновременном развитии у цветущего растения 8–9 розеточных и 4 генеративных побегов. Перерыв в цветении автор связывает с появлением озимых розеточных побегов из почек возобновления, не зацветающих на следующий год.
В континентальном климате юга Сибири наблюдается несколько путей развития почек возобновления у P. tuberosa. Большая часть почек возобновления (I2) замедляет свое развитие. Весной емкость их увеличивается только на 1 метамер (в течение года появляются еще 1–2), и они переходят в разряд спящих. Только одна (редко 2) почка возобновления увеличивается и становится зрелой инициальной. Дальнейшее развитие этой почки зависит от активности ее меристемы весной. Высокая активность выражается в заложении дважды (в апреле и в мае) по 5–6 метамеров, что в сумме дает 10–12 метамеров, из них 9–10 метамеров несут цветочные зачатки. Из такой финальной почки развивается моноциклический полурозеточный побег. Но чаще заложение зачатков в инициальной почке проходит не столь интенсивно, и в конце апреля появляются только 2‒4 новых метамера без генеративных зачатков – из такой почки развивается розеточный побег с 1–2 парами розеточных листьев. В свою очередь, от активности верхушечной почки (М1) растущего розеточного побега следующей весной (2-й вегетационный сезон) зависит развитие дициклического полурозеточного побега. Как и при формировании финальной почки из инициальной, переход М1 в М2-почку, а затем в финальную почку (F2) происходит при заложении новых 10–12 метамеров (9–10 верхних метамеров с цветочными зачатками). В результате развивается удлиненная часть дициклического полурозеточного побега. Можно предположить, что развитие дициклических побегов из почек возобновления на заливном лугу в Калужской области происходит, видимо, по такой же схеме. Следовательно, описанный ход развития из почки возобновления дициклического побега типичен для P. tuberosa. Это подтверждается и нашими данными, что 70–90% дициклических побегов развивается именно из почек возобновления.
Однако не всегда в весенний период в апикальной части розеточного побега закладываются метамеры с цветочными зачатками. В отсутствии генеративных зачатков в верхушечной почке розеточного побега пластические вещества используются для окончательного созревания 1–2 почек возобновления, заложенных еще в прошлом году. Одна из них (редко две) прорастает в побег следующего порядка, а материнский розеточный побег заканчивает свою вегетацию.
Максимальный период ежегодного цветения P. tuberosa составляет 6 лет, но чаще 2–4 года. В исследованных нами местообитаниях цветущие особи развивают в основном моноциклические генеративные побеги (в 75–80% случаев), формирующиеся чаще из спящих почек (70–75% от общего числа моноциклических побегов), и только 25–30% моноциклических побегов развиваются из почек возобновления. Перерыв в цветении особи может длиться от 1 до 4-х лет: в надземной части развиваются 1–2 розеточных однолетних побега. Отсутствие генеративных побегов связано как с внутриорганизменными процессами, так и с условиями произрастания: в степных сообществах период ежегодного цветения короче (2–4 года) по сравнению с лугом (3–6 лет).
Итак, большая роль в побегообразовании P. tuberosa на юге Сибири принадлежит спящим почкам, из которых чаще всего развиваются моноциклические полурозеточные побеги. Это не вызывает изменения жизненной формы вида, в отличие от других представителей сем. Lamiaceae. У некоторых видов Scutellaria L. (Cheryomushkina, Guseva, 2015) и Nepeta L. (Astashenkov, Cheryomushkina, 2019) при пробуждении спящих почек развиваются побеги, отличающиеся от других побегов своей геофильной частью. Они служат основой для построения новых побеговых систем, что приводит к трансформации жизненной формы вида. У P. tuberosa побеги, развившиеся из спящей почки или почки возобновления, могут отличаться только цикличностью. Наши исследования показали, что цикличность побега существенно не влияет на структуру полурозеточных побегов: последовательность функциональных зон не меняется. Условия произрастания оказываются фактором, определяющим мощность побегов независимо от его цикличности (длина и масса полурозеточного побега, масса его флоральной части), а также вызывающим изменение соотношения числа розеточных и стеблевых листьев. На разнотравно-злаковом лугу особи развивают полурозеточные побеги наибольшей мощности по сравнению с растениями, произрастающими в степных ценозах и имеющими в 3 раза меньшие значения основных параметров побегов. В степных сообществах происходит увеличение числа розеточных листьев при уменьшении числа листьев в удлиненной части побега, а также уменьшение синфлоресценции, которая у большинства особей представлена только терминальной флоральной единицей (главным соцветием) без паракладиев. Развитие главного соцветия при этом достаточно стабильно. У дициклического побега его размеры мало зависят от условий произрастания (3–6 метамеров). Главное соцветие моноциклического побега характеризуется большей лабильностью числа метамеров: она меняется от 2–7 метамеров в ксерофильных условиях до 4–9 метамеров в более мезофильных условиях. Но по среднему значению репродуктивного усилия растения с побегами разной цикличности из разных ценозов, не имеют достоверных отличий, что подтверждает стабильность флоральной части побега независимо от его цикличности.
В рассмотренных лугово-степных сообществах юга Сибири у генеративного растения P. tuberosa в надземной части развивается в основном 1–2 моноциклических полурозеточных побега, появление одновременно удлиненного и розеточного побегов наблюдается редко. По сравнению с Калужской областью здесь происходит сокращение периода вегетации особи, что выражается в наличии только весенне-летней генерации листьев и выпадении осенней. Ди- и полициклические побеги у растения встречаются значительно реже.
Таким образом, для P. tuberosa характерно преобладание моноциклических полурозеточных побегов в условиях континентального (г. Новосибирск) и резко континентального (Горный Алтай) климата, а дициклических побегов – в более мягких условиях европейской части России (Калужская область). Выявленная у P. tuberosa изменчивость цикличности побегов в зависимости от условий произрастания отмечается у целого ряда видов, относящихся к разным семействам (Serebryakov, 1959). Это позволяет согласиться с предложением J. Klimešova (Klimešova et al., 2016) о перспективности использования цикличности побега как одного из информативных косвенных признаков в функциональном анализе сообщества.
Список литературы
Astashenkov A.Yu., Cheryomushkina V.A. 2019. The role of dormant buds in transformation of herbaceous biomorphs (by the example of Nepeta L. sect. Spicatae (Benth.) Pojark.). – Biol. Bull. 46 (6): 562–569. https://doi.org/10.1134/S1062359019040046
[Belostokov] Белостоков Г.П. 1962. Ритм сезонного развития растений полупустыни. – Бюлл. МОИП. Отд. биол. 67 (6): 68–80.
[Borisova, Popova] Борисова И.В., Попова Т.А. 1990. Разнообразие функционально-зональной структуры побегов многолетних трав. – Бот. журн. 75 (10): 1420–1426.
Chapin F.S., Bret-Harte M.S., Hobbie S.E., Zhong H. 1996. Plant functional types as predictors of transient responses of arctic vegetation to global change. – J. Veg. Sci. 7 (3): 347–358. https://doi.org/10.2307/3236278
Chave J., Coomes D., Jansen St., Lewis S.L., Swenson N.G., Zanne A.E. 2009. Towards a worldwide wood economics spectrum. – Ecol. Lett. 12: 351–366. https://doi.org/10.1111/j.1461-0248.2009.01285.x
Cheryomushkina V.A., Guseva A.A. 2015. Life forms of Scutellaria supina L. (Lamiaceae) – Contem. Probl. Ecol. 8 (5): 624–635. https://doi.org/10.1134/S1995425515050054
Cornelissen J.H.C., Lavorel S., Garnier E., Diaz S., Buchmann N., Gurvich D.E., Reich P.B., H. ter Steege H., Morgan H.D., van der Heijden M.G.A., Pausas J.G., Poorter H. 2003. A handbook of protocols for standardised and easy measurements of plant functional traits worldwide. – Aust. J. Bot. 51: 335–380. https://doi.org/10.1071/BT02124
[Doronkin] Доронькин В.М. 1997. Phlomis L. – Зопник. – Флора Сибири. Новосибирск. 11: 188, 267.
[Fedoskin] Федоскин Н.В. 1974. Жизненные формы травянистых растений степей юго-восточного Забайкалья. – Бюлл. МОИП. Отд. биол. 29 (6): 98–108.
[Golubev] Голубев В.Н. 1965. Эколого-биологические особенности травянистых растений и растительных сообществ лесостепи. М. 287 с.
[Gorshkova] Горшкова А.А. 1966. Биология степных пастбищных растений Забайкалья. М. 272 с.
[Gorshkova] Горшкова А.А. 1975. Ритмика развития и экология степных растений Забайкалья. – Бот. журн. 60 (6): 889–893.
[Grigorieva] Григорьева Н.М. 1994. Структура особи и онтогенез Phlomis tuberosа L. – В сб.: Успехи экологической морфологии растений и ее влияние на смежные науки. М. С. 85–86.
Klimešová J., Nobisb M., Herben T. 2016. Links between shoot and plant longevity and plant economics spectrum: environmental and demographic implications. – Perspect. Plant Ecol. Evol. Syst. 22: 55–62. https://doi.org/10.1016/j.ppees.2016.09.002
Klimešová J., Danihelka J., Chrtek J., Bello F., Herben T. 2017. CLO-PLA: a database of clonal and bud-bank traits of the Central European flora. – Ecology. 98 (4): 1179. http://onlinelibrary.wiley.com/doi/10.1002/ecy.1745/suppinfo
[Komarevtseva et al.] Комаревцева Е.К., Асташен-ков А.Ю., Гордеева Н.И., Гусева А.А., Курочкина Н.Ю. 2018. Phlomoides tuberosa (L.) Moench на юге Сибири: биология и состояние ценопопуляций. – Растительный мир Азиатской части России. 4: 55–64.
[Komarova] Комарова Т.А. 1989. Внутрипочечное и внепочечное развитие побега как единый процесс. Автореф. дисс. …канд. биол. наук. М. 16 с.
[Kuznetsova, Timonin] Кузнецова Т.В., Тимонин А.К. 2017. Соцветие: морфология, эволюция, таксономическое значение (применение комплементарных подходов). М. 183 с.
[Makunina] Макунина Н.И. 2016. Растительность лесостепи Западно-Сибирской равнины и Алтае-Саянской горной области. Новосибирск. 183 с.
[Markov, Pleshchinskaya] Марков М.В., Плещинская Е.Н. 1987. Репродуктивное усилие у растений. – Журн. общ. биол. 68 (1): 77–83.
McIntyre S., Lavorel S., Tremont R.M. 2006. Plant Life-History Attributes: Their Relationship to Disturbance Response in Herbaceous Vegetation. – J. Ecol. 83 (1): 31. https://www.jstor.org/stable/2261148
Mommer L., Weemstra M. 2012. The role of roots in the resource economics spectrum. – New Phytol. 195: 725–727. https://doi.org/10.1111/j.1469-8137.2012.04247.x
[Polyntseva] Полынцева Н.А. 1971. Ритм годичного развития растений березовых перелесков Таштыпской лесостепи (Минусинская котловина). – Бюлл. МОИП. Отд. биол. 76 (5): 96–106.
[Serebryakov] Серебряков И.Г. 1952. Морфология вегетативных органов высших растений. М. 392 с.
[Serebryakov] Серебряков И.Г. 1959. Типы развития побегов у травянистых многолетников и факторы их формирования. – Ученые записки МГПИ им. В.П. Потемкина. Вопросы биологии растений. 100 (5): 3–38.
[Serebryakov] Серебряков И.Г. 1964. Сравнительный анализ некоторых признаков ритма сезонного развития растений различных ботанико-географических зон СССР. – Бюлл. МОИП. Отд. биол. 69 (5): 62–75.
[Serebryakova] Серебрякова Т.И. 1956. Побегообразование и ритм сезонного развития растений заливных лугов Средней Оки. – Ученые записки. 97(3): 45–120.
[Serebryakova] Серебрякова Т.И. 1971. Морфогенез побегов и эволюция жизненных форм злаков. М. 359 с.
[Serebryakova] Серебрякова Т.И. 1983. Почка как этап развития побега. – В сб.: Тез VII съезда Всесоюзного ботанического общества. Донецк. Л. С. 232–233.
Troll W. 1964. Die Infloreszenzen. Bd 1. Jena. 615 S.
[Tsatsenkin] Цаценкин И.А. 1967. Экологические шкалы для растений пастбищ и сенокосов горных и равнинных районов Средней Азии, Алтая и Урала. Душанбе. 226 с.
Vojtkó A., Freitag M., Bricca A., Martello F., Compañ J.M., Küttim M., Kun R., Bello F., Klimešová J., Götzenberger L. 2017. Clonal vs leaf-height-seed (LHS) traits: which are filtered more strongly across habitats? – Folia Geobot. 52: 269–281. https://doi.org/10.1007/s12224-017-9292-1
Weiher E., van der Werf A., Thompson K., Roderick M., Garnier E., Eriksson O. 1999. Challenging Theophrastus: a common core list of plant traits forfunctional ecology. – J. Veg. Sci. 10 (5): 609–620. https://www.jstor.org/stable/3237076
Westoby M. 1998. A leaf–height–seed (LHS) plant ecology strategy scheme. – Plant Soil. 199 (2): 213–227. https://doi.org/10.1023/A:1004327224729
[Zaitsev] Зайцев Г.Н. 1991. Математический анализ биологических данных. М. 182 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал