

Ботанический журнал, 2021, T. 106, № 1, стр. 52-60
НОВЫЕ ВИДЫ ДИАТОМОВЫХ (BACILLARIOPHYTA) ДЛЯ ФЛОРЫ ЗАЛИВА КАРА-БОГАЗ-ГОЛ (КАСПИЙСКОЕ МОРЕ)
Международный институт моделирования и прогнозирования развития морских и гипергалинных экосистем
141603 Московская обл., Клин, пос. Майданово, 18, 19, Россия
* E-mail: mimge_rus@mail.ru
** E-mail: mimge2020@gmail.com
Поступила в редакцию 27.02.2020
После доработки 15.09.2020
Принята к публикации 29.09.2020
Аннотация
Впервые проведены электронно-микроскопические исследования с использованием сканирующего электронного микроскопа диатомовых водорослей (Bacillariophyta) из гипергалинного залива Кара-Богаз-Гол, позволившие выявить 8 новых для флоры залива видов, способных обитать при солености воды от 44.0 до 240.0‰. Выявлено, что клетки Staurosira binodis и Fragilaria capucina отличаются меньшим числом структурных элементов на створках, чем указывалось ранее в диагнозах. Размеры створок S. binodis из залива также расходятся с общепринятыми диагнозами. Показан предел солености, при котором в условиях залива происходит редукция количества структурных элементов на створках диатомовых водорослей. Виды Aulacoseira ambigua, Plano-thidium lanceolatum, S. binodis, F. rumpens, F. vaucheriae, F. famelica и Pantocsekiella ocellata впервые приводятся для Каспийского моря.
Залив Кара-Богаз-Гол является самым крупным природным соляным резервуаром морского происхождения, расположенным в восточной части Каспийского моря на территории Туркменистана, с длиной по меридиану 165 км и параллели 154 км и границами между 50°40′E—52°30′E и 40°35′N—41°50′N (Akovetskiy, Bogdanov, 1988). Залив Кара-Богаз-Гол связан с Каспийским морем проливом Карабогазгол, через который морские воды Каспия в огромных количествах поступают в залив (Leroy et al., 2006). Площадь поверхности зеркала залива составляет около 18000 км2, уровень воды в нем на 28 м ниже уровня Каспийского моря. Соленость вод залива колеблется от 40.0 до 270.0‰ (Bulatov, Shakirova, 2005). Более постоянная величина солености в заливе отмечается в его центральной части, менее постоянная – в литоральной зоне водоема.
Первые сведения по альгофлоре залива Кара-Богаз-Гол содержатся в работе А.Д. Пельша (Pel’sh, 1936). Позднее Н.И. Караева (Karayeva, 1972) приводит сведения об обитании 21 вида и разновидности диатомовых водорослей в заливе Кара-Богаз-Гол при солености 20.4‰. Более подробное изучение таксономического разнообразия и экологии диатомовых водорослей в заливе Кара-Богаз-Гол были проведены в начале 2000-х годов, в результате чего список был расширен до 65 видов и разновидностей (Bulatov, 2002, 2004). Причем, из 21 таксона диатомовых водорослей, упоминаемых в работе Н.И. Караевой, для залива Кара-Богаз-Гол, автором настоящей работы было отмечено только 5 таксонов. Возможно, что изменение таксономического разнообразия диатомовых водорослей в заливе Кара-Богаз-Гол связано с его полным перекрытием в 1980 г. дамбой, оказавшей существенное влияние на химический состав рассолов (Bulatov, 2020), в результате чего залив полностью высох. В 1992 г. дамба, преграждавшая путь морских вод из Каспийского моря в залив, была разрушена, отток вод в залив был восстановлен.
Недавние исследования диатомовых водорослей залива Кара-Богаз-Гол позволили описать новый для науки вид – Brachysira carabogazgolensis Bulatov, обитающий при солености 50.0‰ (Bulatov, 2020). Следует отметить, что этот вид является единственным в мире представителем рода Brachysira Kützing, обитающим исключительно в условиях гипергалинных вод.
Наши предыдущие исследования показали, что диатомовые водоросли залива Кара-Богаз-Гол представлены множеством мелких форм, структура которых трудно различима при изучении с помощью световой микроскопии. В этой связи, для уточнения видовой принадлежности таких мелких форм, мы использовали метод электронной микроскопии.
В ходе проведенных исследований было выявлено 8 новых для флоры Кара-Богаз-Гола видов диатомовых водорослей, в отношении которых ранее отсутствовали сведения об обитании в природных рассолах, семь из них являются новыми для Каспийского моря.
МАТЕРИАЛЫ И МЕТОДЫ
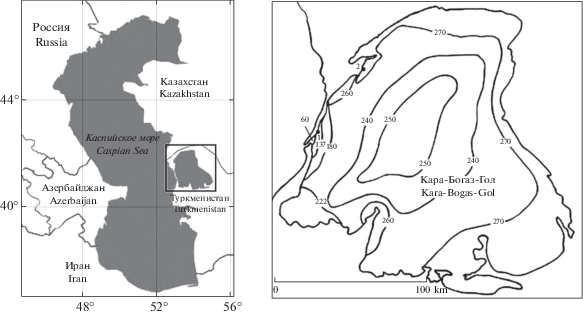
Материалом для настоящей статьи послужили пробы, собранные автором в течение 2000–2001 гг. с двух станций в акватории залива Кара-Богаз-Гол, первая станция располагалась в акватории зоны смешения вод залива и Каспийского моря (далее по тексту – зона смешения вод), в районе с координатами 41°06′43′′N, 52°54′32′′E, а вторая – у косы Карасукут, в районе с координатами 41°32′51′′N, 52°52′13′′E (рис. 1).
Рис. 1.
Карта-схема расположения залива Кара-Богаз-Гол с указанием величин солености на акватории залива, по литературным данным, в 2000–2001 гг. (Bulatov, Shakirova, 2005), а также расположением станций отбора проб диатомовых водорослей. Станция 1 – зона смешения вод, район с координатами 41°06′43′′N – 52°54′32′′E; Станция 2 – коса Карасукут, район с координатами 41°32′51′′N – 52°52′13′′E.
Fig. 1. Schematic map of the location of Kara-Bogaz-Gol Bay, indicating salinity values in the area of the bay according to published data in 2000–2001 (Bulatov, Shakirova, 2005), and the diatom sampling stations. Station 1 – mixing zone, 41°06′43′′N – 52°54′32′′E; Station 2 – Karasukut, 41°32′51′′N – 52°52′13′′E.
Освобождение створок от органического содержимого проводили методом сжигания концентрированной серной кислотой (Opredelitel’…, 1951; Vodorosli …, 1989). Исследование проводилось на базе Института биологии внутренних вод им. И.Д. Папанина РАН (пос. Борок, Ярославская область) с помощью сканирующего электронного микроскопа JEOL JSM-25S. Препараты напылялись золотом с помощью установки марки Eico IB 3 ion coater.
Фотографии створок диатомовых водорослей из залива Кара-Богаз-Гол являются частью коллекции С.А. Булатова, находящейся в Международном институте моделирования и прогнозирования развития морских и гипергалинных экосистем.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Впервые проведено изучение диатомовых водорослей залива Кара-Богаз-Гол с использованием сканирующей электронной микроскопии, позволившей выявить особенности структуры створок мелких форм диатомовых водорослей в условиях обитания в водах с высоким содержанием солей. Описание 8 новых для залива Кара-Богаз-Гол видов диатомовых, а также их фотографии, приводятся ниже.
Aulacoseira ambigua (Grunow) Simonsen 1979 (табл. I, 1–3)
Таблица I. 1–3 – Aulacoseira ambigua (1 – соединенные створки; 2 – видны соединительные шипы и форма ареол; 3 – структура загиба створки); 4–6 – Planothidium lanceolatum (4 – шовная створка, 5, 6 – бесшовная створка). Масштабная линейка: 1–6 – 10 мкм.
Table I. 1–3 – Aulacoseira ambigua (1 – connected valves; 2 – view of linking spines and areolae shape; 3 – structure of valve mantle); 4–6 – Planothidium lanceolatum (4 – raphe valve, 5, 6 – rapheless valve). Scale bars: 1–6 – 10 µm.

Панцирь высокоцилиндрический. Створки диаметром 6.4–9.8 мкм. Загиб высотой 14.4–16.6 мкм. Структура загиба створки в виде продольных спирально закрученных рядов ареол, 16–17 поперечных рядов ареол в 10 мкм и 17–18 продольных рядов ареол в 10 мкм. Соединительные шипы – раздвоенные на концах.
При изучении в световом микроскопе A. ambigua внешне сходен с A. granulata (Ehrenberg) Simonsen. Но детальное изучение структуры A. ambigua из залива Кара-Богаз-Гол в сканирующем электронном микроскопе выявило ряд морфологических отличий от A. granulata, в том числе в строении соединительных шипов. На особенность строения соединительных шипов у A. ambigua, которые раздвоены на концах, как отличительный признак от A. granulata, также указывают в своей работе K. Krammer и H. Lange-Bertalot (1991а). Кроме того, на наш взгляд, отличительной особенностью A. ambigua от A. granulata является большее число ареол в поперечных и продольных рядах, что также не опровергается другими исследователями (Usol’tseva, Likhoshvay, 2007; Genkal, Chekryzheva, 2015; Kulikovskiy et al., 2016). Еще одной характерной особенностью A. ambigua, отличающей его от A. granulata, является наличие спирально правосторонне закрученных рядов ареол, берущих начало между шипами (Houk, 2003, Tab. XXVIII , Fig. 1–15 ), что также наблюдалось на исследованном нами материале.
Следует отметить, что ранее исследователями диатомовых водорослей Каспийского моря А.И. Прошкиной-Лавренко и И.В. Макаровой ставилась под сомнение возможность обитания A. ambigua в Каспийском море, а все ранее встречаемые единичные клетки A. ambigua, по их мнению, были отмершими, случайно попавшими в воды Северного Каспия с водами рек Волги и Урала (Proshkina-Lavrenko, Makarova, 1968). Однако, как показали наши исследования, вид A. ambigua свободно акклиматизировался в условиях высокосоленых вод залива Кара-Богаз-Гол, где отмечался в большом количестве в районе зоны смешения вод, характеризующейся непостоянством минерализации.
В заливе вид отмечался в прибрежном планктоне в виде колоний, состоящих в основном из соединенных между собой двух, реже трех клеток, при солености 50.0–210.0‰ и температуре воды 18.0–22.5°С. Максимальное развитие A. ambigua отмечалось в мае–июле и октябре в районе зоны смешения вод.
Для Каспийского моря вид указывается впервые.
Ранее считалось, что A. ambigua относится к планктонным пресноводным видам, предпочитающим мезотрофные и эвтрофные водоемы (Opredelitel’ …, 1951; Krammer, Lange-Bertalot 1991а; Genkal, Vekhov, 2007; Genkal, Trifonova, 2009; Kulikovskiy et al., 2016).
Planothidium lanceolatum (Brébisson) Lange-Bertalot 1999 (табл. I, 4–6)
Створки ланцетные, короткие или длинные, длиной 16.8–31.8 мкм, шириной 7.3–9.5 мкм, концы широкозакругленные. На створке со швом имеется линейное осевое поле, центральное поле прямоугольно расширено в стороны, не доходит до краев створки. На бесшовной створке расположено линейно-ланцетное осевое поле, в центральной части имеется подковообразная структура. Штрихи радиальные, грубые, с многорядными ареолами на обеих створках, 9–11 штрихов в 10 мкм.
В заливе вид отмечался в прибрежном планктоне зоны смешения вод и косы Карасукут при солености 50.0–240.0‰ и температуре воды 19.0–25.0°С. Максимальное развитие P. lanceolatum наблюдалось в сентябре и октябре.
Для Каспийского моря приводится впервые. Имеются сведения об обитании P. lanceolatum в Иваньковском, Угличском, Горьковском, Чебоксарском водохранилищах, а также низовьях Волги (Genkal, 1992).
Ранее указывался исключительно как пресноводный вид, широко распространенный в водоемах различного типа (Opredelitel’ …, 1951; Kogan et al., 1985; Krammer, Lange-Bertalot, 1991b; Kulikovskiy et al., 2016).
Staurosira binodis (Ehrenberg) Lange-Bertalot 2011 (табл. II, 1-3)
Таблица II. 1–3 – Staurosira binodis (вид со створки); 4 – Fragilaria capucina (вид со створки); 5, 6 – F. rumpens (5 – вид со створки, 6 – конец створки, виден двугубый вырост); 7 – Pantocsekiella ocellata (вид со створки); 8, 9 – F. vaucheriae (8 – вид структуры створки, 9 – вид со створки, видна центральная часть створки); 10 – F. famelica (вид со створки). Масштабные линейки: 1–6, 9, 10 – 10 мкм, 7, 8 – 1 мкм.
Table. II. 1–3 – Staurosira binodis (valve view); 4 – Fragilaria capucina (valve view); 5, 6 – F. rumpens (5 – valve view, 6 – valve from the end, view of labiate process); 7 – Pantocsekiella ocellata (valve view); 8, 9 – F. vaucheriae (8 – valve structure, 9 – valve view, central part of the valve is visible); 10 – F. famelica (valve view). Scale bars: 1–6, 9, 10 – 10 µm, 7–8 – 1 µm.

Клетки одиночные. Створки широколанцетные, посередине суженные, длиной 22.3–25.7 мкм, шириной 5.4–8.6 мкм. Концы оттянутые, клювовидные. Осевое поле линейное или ланцетное. Штрихов 8–11 в 10 мкм.
У встреченного в заливе S. binodis количество штрихов на створках было значительно меньше (8–11 в 10 мкм), в отличие от указаний в диагнозах (13–16 в 10 мкм) (Kulikovskiy et al., 2016). Из литературных источников известно, что длина и ширина створок S. binodis составляет 7.0–21.0 мкм и 4.0–6.0 мкм, соответственно (Kulikovskiy et al., 2016), наши исследования показали, что S. binodis обладает створками большего размера (22.3–25.7 мкм и 5.4–8.6 мкм).
В заливе вид встречался в прибрежном планктоне, при солености 50.0–240.0 ‰ и температуре воды 19.0–25.0°С. В больших количествах отмечался в сентябре и октябре в районах зоны смешения вод и косы Карасукут.
Для Каспийского моря S. binodis приводится впервые.
Солоноватоводный широко распространенный вид (космополит), в стоячих эвтрофных и мезотрофных водоемах, а также среди обрастаний (Opredelitel’ …, 1951; Krammer, Lange-Bertalot, 1991а; Kulikovskiy et al., 2016).
Fragilaria capucina Desmazières 1830 (табл. II, 4)
Створки линейные, с ровными краями и оттянутыми головчатыми концами, длиной 53.9–57.8 мкм, шириной 3.6–5.8 мкм. Осевое поле линейное. Центральное прямоугольное. Штрихи линеолированные, 8–10 в 10 мкм.
В существующих диагнозах для данного вида приводится 12–17 штрихов в 10 мкм (Krammer, Lange-Bertalot, 1991a; Hofmann et al., 2011; Kulikovskiy et al., 2016).
Вид отмечался в прибрежном планктоне, при солености 240.0‰ и температуре воды 25.0°С. Максимальное развитие наблюдалось в сентябре в районе косы Карасукут.
В литературе имеются указания на обитание F. capucina в составе планктона в иранском секторе Каспийского моря (Bagheri, Fallahi, 2014).
Солоноватоводный широко распространенный вид (Opredelitel’ …, 1951; Krammer, Lange-Bertalot, 1991а).
F. rumpens (Kützing) G.W.F. Carlson 1913 (табл. II, 5, 6)
Створки ланцетные, посередине слегка расширенные, с клювовидными широко закругленными концами, длиной 65.4–68.0 мкм, шириной 5.0–5.5 мкм. Поперечные штрихи линеолированные, 16–18 в 10 мкм. Центральное поле почти округлое, осевое – линейное. На одном из концов створки имеется двугубый вырост.
В заливе отмечен в прибрежном планктоне при солености 73.2 ‰ и температуре воды 22.5°С. Развитие наблюдалось в июне в районе зоны смешения вод.
Для Каспийского моря F. rumpens приводится впервые. Имеются сведения об обитании вида в Иваньковском, Угличском, Рыбинском, Горьковском, Куйбышевском водохранилищах (Genkal, 1992).
Ранее считался исключительно пресноводным широко распространенным видом, встречающимся в водоемах различного типа (Opredelitel’ …, 1951; Hofmann et al., 2011; Kulikovskiy et al., 2016).
F. vaucheriae (Kützing) J.B. Petersen 1938 (табл. II, 8, 9)
Створки удлиненно ланцетные, длиной 36.4 мкм, шириной 3.6 мкм, концы оттянутые, почти головчатые. Штрихи пунктирные, радиальные, 10 в 10 мкм. Осевое поле ланцетное, центральное расширенное в одном направлении до края створки.
В заливе F. vaucheriae отмечен в прибрежном планктоне, при солености 50.0‰ и температуре воды 19.0°С. В небольшом количестве вид развивался в октябре в районе зоны смешения вод.
Для Каспийского моря вид приводится впервые. Имеются сведения об обитании F. vaucheriae по всей Волге (Genkal, 1992).
Солоноватоводный широко распространенный вид, встречается в литорали стоячих и текучих водоемов, обрастая макрофиты (Opredelitel’ …, 1951; Krammer, Lange-Bertalot, 1991а; Kulikovskiy et al., 2016).
F. famelica (Kützing) Lange-Bertalot 1980 (табл. II, 10)
Створки линейно-ланцетные, сужающиеся к широко закругленным концам, длиной 18.0 мкм, шириной 3.9 мкм. Штрихи прямые, в виде ареол, 17 в 10 мкм. Осевое поле узколинейное, центральное поле односторонне расширено к краю створки c короткими штрихами.
В заливе встречен в прибрежном планктоне, при солености 44.0 ‰ и температуре воды 5.0°С. Развитие F. famelica наблюдалось в октябре в районе зоны смешения вод.
Для Каспийского моря вид приводится впервые.
Пресноводный и солоноватоводный вид (Hofmann et al., 2011).
Pantocsekiella ocellata (Pantocsek) K.T. Kiss et Acs 2016 (табл. II, 7)
Створки круглые, 6.0–8.6 мкм в диаметре. Краевая зона с короткими штрихами, различной длины, 20–23 штриха в 10 мкм. В центре створки расположено 2–3 выроста с опорами, в краевой зоне створки также имеются выросты с опорами в количестве 7–8, располагающиеся через каждые четыре-шесть штрихов.
В заливе вид встречался в прибрежном планктоне на глубине 0.5 м при солености 50.0–72.3‰ и температуре воды 18.0–22.5°С. Максимальное развитие наблюдалось в июле и октябре в районе зоны смешения вод.
Для Каспийского моря вид приводится впервые.
По данным более ранних литературных источников, P. ocellata является олигогалобом, предпочитающим пресные воды (Krammer, Lange-Bertalot, 1991а; Acs et al., 2016).
ЗАКЛЮЧЕНИЕ
Полученные нами результаты указывают на перспективность продолжения исследований диатомовых водорослей в гипергалинном заливе Кара-Богаз-Гол с целью установления их более полного таксономического разнообразия, формируемого в условиях высокосоленых вод уникального природного водоема, а также выявления различных экологических форм, способных адаптироваться в условиях высокого содержания солей в воде.
Анализ литературных данных по схожим экосистемам показал, что видовой состав диатомовых водорослей залива Кара-Богаз-Гол существенно отличался от видового состава, географически близко расположенного и сопоставимого по размерам Аральского моря на этапе ультрагалинизации. Так, для донных отложений Арала, соленость которого колебалась от 81.0 до 136.0‰, в период с 2002 по 2009 г. приводится список из 97 таксонов диатомовых водорослей (Sapozhnikov, 2010; Sapozhnikov et al., 2009), из которых только 17 таксонов отмечены в заливе Кара-Богаз-Гол. Центрическая диатомея Actinocyclus octonarius Ehrenberg, обитающая в мелководном Кара-Богаз-Голе при солености 40.0–250.0‰ (Bulatov, 2002), отмечалась в Арале на глубине 24.6 м при солености 92.0‰, и в изобилии встречалась до глубины 15 м (Sapozhnikov et al., 2009). Кроме A. octonarius, в Аральском море развивались Campylodiscus neofastuosus Ruck et Nakov при солености 93.0‰ и Nitzschia sigma (Küt-zing) W. Smith при солености 136.0‰ (Sapozhnikov, 2010), являющиеся обычными для береговых альгоценозов залива Кара-Богаз-Гол.
Установлено, что наибольшей изменчивостью в условиях высокой минерализации вод залива обладают популяции Staurosira Ehrenberg и Fragilaria Lyngbye, которая проявилась в виде увеличения размеров створок и уменьшения числа структурных элементов на створках, отличного от их количества, указываемого в ранее известных систематических диагнозах. Признаки изменчивости в виде редукции количества структурных элементов створок диатомовых водорослей, в условиях залива Кара-Богаз-Гол отмечались уже при солености 240.0‰. Аналогичная картина наблюдалась нами ранее у некоторых видов рода Mastogloia Thwaites ex W. Smith (Bulatov, 2013). Из восьми новых для залива Кара-Богаз-Гол видов диатомовых семь впервые приводятся для Каспийского моря.
Список литературы
[Akovetskiy, Bogdanov] Аковецкий В.И., Богданов И.Я. 1988. Кара-Богаз-Гол: вчера, сегодня, завтра. Ашхабад. 195 с.
Acs É., Ari E., Duleba M., Dressler M., Genkal S.I., Jakó É., Rimet F., Ector L., Kiss K.T. 2016. Pantocsekiella, a new centric diatom genus based on morphological and genetic studies. – Fottea. 16 (1): 56–78. https://doi.org/10.5507/fot.2015.028
Bagheri S., Fallahi M. 2014. Checklist of phytoplankton taxa in the Iranian Waters of the Caspian Sea. – Caspian J. Env. Sci. 12 (1): 81–97.
[Bulatov] Булатов С.А. 2002. Структура и состояние диатомовых сообществ высокоминерализованных вод залива Кара-Богаз-Гол. – В сб.: Тез. докл. VIII школы диатомологов России и стран СНГ “Морфология, экология и биогеография диатомовых водорослей”. Борок. С. 11–12.
[Bulatov] Булатов С.А. 2004. Особенности экологии артемии ультрагалинного залива Кара-Богаз-Гол и некоторые аспекты ее питания в естественных условиях. – В сб.: Мат. докл. межд. научно-исслед. конф. “Биоразнообразие Артемии в странах СНГ: современное состояние ресурсов и их использование”. Тюмень. С. 94–101.
[Bulatov] Булатов С.А. 2013. Об обитании видов родов Navicula и Mastogloia в водах залива Кара-Богаз-Гол. – В сб.: Матер. XIII Межд. науч. конф. альгологов “Диатомовые водоросли: современное состояние и перспективы исследований”. Борок. С. 114–115.
Bulatov S.A. 2020. A New Species of the Bacillariophyta from the Kara-Bogaz-Gol Bay (Caspian Sea, Turkmenistan). – International Journal on Algae. 22 (2): 123–128. https://doi.org/10.1615/InterJAlgae.v22.i2.20
[Bulatov, Shakirova] Булатов С.А., Шакирова Ф.М. 2005. Современное состояние популяции Artemia sp. (Crustacea, Branchiopoda) залива Кара-Богаз-Гол и перспективы ее рационального использования. – Selevinia. P. 123–128.
[Genkal] Генкал С.И. 1992. Атлас диатомовых водорослей планктона реки Волги. СПб. 128 с.
[Genkal, Vekhov] Генкал С.И., Вехов Н.В. 2007. Диатомовые водоросли водоемов Русской Арктики. Архипелаг Новая Земля и остров Вайгач. М. 64 с.
[Genkal, Trifonova] Генкал С.И., Трифонова И.С. 2009. Диатомовые водоросли планктона Ладожского озера и водоемов его бассейна. Рыбинск. 72 с.
[Genkal, Chekryzheva] Генкал С.И., Чекрыжева Т.А. 2015. Центрические диатомовые водоросли озер южной части Республики Карелия (Вендюрская группа и Заонежье). – Биология внутренних вод. 3: 5–13. https://doi.org/10.7868/S0320965215030043
Hofmann G., Werum M., Lange-Bertalot H. 2011. Diatomeen im Sübwasser-Benthos von Mitteleuropa. Konigstein. 908 p.
Houk V. 2003. Atlas of freshwater centric diatoms with a brief key and description. Part I. Melosiraceae, Orthoseiraceae, Paraliaceae and Aulacoseiraceae. – Czech Phycology Suppl. 1: 3–29.
[Karayeva] Караева Н.И. 1972. Диатомовые водоросли бентоса Каспийского моря. Баку. 258 с.
[Kogan et al.] Коган Ш.И., Любезнов Ю.Е., Садыков Х.С. 1985. К альгофлоре водоемов Ташаузской области Туркменской ССР. – Изв. акад. наук Туркм. ССР. Сер. биол. наук. 6: 3–9.
Krammer K., Lange-Bertalot H. 1991a. Bacillariophytae. Teil 3. Centrales, Fragilariaceae, Eunotiaceae. Süsswasserflora von Mitteleuropa. Bd. 2/3. Stuttgart. 576 p.
Krammer K., Lange-Bertalot H. 1991b. Bacillariophytae. Teil 4. Achnanthaceae und Kritische Ergaenzungen zu Navicula (Lineolatae) und Gomphonema. Süsswasserflora von Mitteleuropa. Bd. 2/4. Stuttgart. 437 p.
[Kulikovskiy et al.] Куликовский М.С., Глущенко А.М., Генкал С.И., Кузнецова И.В. 2016. Определитель диатомовых водорослей России. Ярославль. 804 с.
Leroy S.A.G., Marret F., Giralt S., Bulatov S.A. 2006. Natural and anthropogenic rapid changes in the Kara-Bogaz Gol over the two centuries reconstructed from palynological analyses and a comparison to instrumental records. – Quaternary International. 150 (1): 52–70. https://doi.org/10.1016/j.quaint.2006.01.007
[Opredelitel’ …] Определитель пресноводных водорослей СССР: Диатомовые водоросли. 1951. Вып. 4. М. 618 с.
[Pel’sh] Пельш А.Д. 1936. К гидробиологии Карабугаза. – Тр. Соляной лаборатории АН СССР. Вып. 5. С. 49–80.
[Proshkina-Lavrenko, Makarova] Прошкина-Лавренко А.И., Макарова И.В. 1968. Водоросли планктона Каспийского моря. Л. 291 с.
[Sapozhnikov] Сапожников Ф.В. 2010. Ценозы донных микроводорослей Большого Арала на этапе ультрагалинизации: Дис. … канд. биол. наук. М. 179 с.
Sapozhnikov F.V., Ivanishcheva P.S., Simakova U.V. 2009. Modern assemblage changes of benthic algae as a result of hypersalinization of the Aral Sea. – Journal of Marine Systems. 76: 343–358. https://doi.org/10.1016/j.jmarsys.2008.03.021
[Usol’tseva, Likhoshvay] Усольцева М.В., Лихошвай Е.В. 2007. Тонкое строение панцирей видов рода Aulacoseira Thwaites (Bacillariophyta) из реки Обь (Россия). – Альгология. 17 (2): 139–147.
[Vodorosli …] Водоросли. Справочник. 1989. Киев. 608 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал