

Ботанический журнал, 2021, T. 106, № 4, стр. 382-396
АНДРОМОНОЭЦИЯ И ЕЕ ПРОЯВЛЕНИЕ В СОЦВЕТИЯХ CHAEROPHYLLUM AROMATICUM (APIACEAE) В МОСКОВСКОЙ ОБЛАСТИ
В. Н. Годин 1, *, Т. В. Архипова 1, А. Н. Тагиева 1
1 Московский педагогический государственный университет
129164 Москва, ул. Кибальчича 6, корп. 3, Россия
* E-mail: vn.godin@mpgu.su
Поступила в редакцию 05.11.2020
После доработки 10.12.2020
Принята к публикации 22.12.2020
Аннотация
Статья посвящена феномену андромоноэции у стержнекорневого травянистого поликарпического растения Chaerophyllum aromaticum. Проанализировано адаптивное значение как наличия тычиночных и обоеполых цветков у одной особи, так и изменения их соотношения в синфлоресценции. В тычиночных цветках наблюдается полная редукция гинецея, от которого остались только подстолбии, выполняющие функции нектарников. Флоральная единица у этого вида – открытый двойной зонтик, состоящий из закрытых простых зонтиков, в которых обоеполые цветки занимают краевое положение, а один обоеполый цветок расположен в центре. Между обоеполыми цветками находятся многочисленные тычиночные цветки. У C. aromaticum в Московской области выделяются три типа особей по степени разветвленности синфлоресценций: а) особи с двойными зонтиками на побегах I и II порядков; б) особи с двойными зонтиками на побегах I, II и III порядков; в) особи с двойными зонтиками на побегах I, II, III и IV порядков. Доля тычиночных цветков увеличивается при переходе от терминальных двойных зонтиков к двойным зонтикам на побегах IV порядка. При увеличении степени разветвленности синфлоресценций C. aromaticum участие тычиночных цветков в двойных зонтиках на побегах одного порядка закономерно уменьшается.
Андромоноэция – форма половой дифференциации, при которой популяции растений состоят из одного типа особей, образующих одновременно обоеполые и тычиночные цветки. Данная половая форма довольно часто встречается у цветковых растений, однако чаще всего характерна для представителей нескольких семейств: Apiaceae, Araliaceae, Fabaceae, Poaceae, Solanaceae и др. (Schlessman et al., 2001; Schlessmann, 2010; Kellogg, 2015; Barboza et al., 2016; Godin, 2017; Plunkett et al., 2018a, b). В каждом из этих семейств андромоноэция наряду с другими присущими им биологическими особенностями способствует сокращению вероятности автогамии и благоприятствует ксеногамии. Семейство Apiaceae характеризуется не только андромоноэцией (а также другими формами половой дифференциации), но и специфической структурой соцветий (разные варианты зонтиков и их агрегаций), наличием различных форм дихогамии (разновременное функционирование тычинок и плодолистиков в виде протандрии или протогинии), оригинальной последовательностью цветения флоральных единиц в синфлоресценции и др. (Knuth, 1898; Kuznetzova, 1982; Dem’yanova, 2000; Godin, Perkova, 2017; Plunkett et al., 2018a). Такой широкий набор механизмов, способствующих ксеногамии, необходим для снижения вероятности автогамии в связи с наличием системы самосовместимости у представителей этого семейства (Bell, 1971).
В результате многочисленных исследований выявлены некоторые закономерности локализации обоеполых и тычиночных цветков в синфлоресценциях у представителей семейства Apiaceae. Взаимное пространственное расположение этих двух половых типов цветков в простых зонтиках разнообразно и специфично для родов или даже отдельных видов. При этом обоеполые цветки чаще краевые, а тычиночные – центральные (Knuth, 1898; Reuther, Claßen-Bockhoff, 2010; Godin, Perkova, 2017; Kuznetzova, Timonin, 2017; Plunkett et al., 2018a). У протандричных видов зонтичных с увеличением порядка ветвления побегов и возраста особей доля тычиночных цветков в двойных зонтиках увеличивается, а обоеполых – снижается (Knuth, 1898; Reuther, Claßen-Bockhoff, 2010). Однако у растений этого же семейства с протогинией наблюдается обратная ситуация: терминальные зонтики характеризуются максимальной долей тычиночных цветков (Schlessman, Barrie, 2004), а в зонтиках на побегах следующих порядков их доля снижается. Тем не менее, у каждого вида проявление подобных закономерностей специфично и, скорее всего, зависит еще от экологических условий произрастания и жизненности особей (Dem’yanova, 2000; Reuther, Claßen-Bockhoff, 2010; Godin et al., 2019).
Chaerophyllum aromaticum L. (бутень ароматный) представляет собой двулетнее или многолетнее полурозеточное каудексовое стержнекорневое поликарпическое травянистое растение, гемикриптофит (Petrova, 2016). Его ареал охватывает европейскую часть России, Среднюю Европу, Средиземноморье, Балканы (Pimenov, Ostroumova, 2012). Одна из отличительных особенностей рода Chaerophyllum L. в трибе Scandicinae – образование терминальных цветков в зонтиках и, таким образом, наличие закрытых простых зонтиков как составных единиц сложных, или двойных, зонтиков (Kuznetzova, 1982; Spalik, Downie, 2001; Plunkett et al., 2018a). У ряда других видов этой трибы также возможно формирование закрытых зонтиков, однако это явление не постоянно. С другой стороны, у многих видов рода Chaerophyllum обоеполые и тычиночные цветки образуются в двойных зонтиках на побегах любого порядка (Knuth, 1898), что позволяет проанализировать характер как размерных, так и количественных характеристик этих типов цветков в синфлоресценции. Другие виды трибы также характеризуются андромоноэцией, однако зачастую формирование обоеполых и тычиночных цветков у них приурочено к разным двойным зонтикам в пределах синфлоресценции (Spalik, Downie, 2001), что делает невозможным получение сравнительной характеристики этих двух половых типов цветков в пределах одного двойного зонтика.
Несмотря на то, что андромоноэция у C. aromaticum известна давно, сведения об особенностях ее проявления у данного вида довольно фрагментарны и отрывочны. Согласно данным P. Knuth (1898), каждый простой зонтик у этого вида содержит центральный и 3–5 краевых обоеполых цветков, тычиночные цветки располагаются в промежуточной зоне между краевыми и центральным (терминальным) обоеполыми цветками. Немного более детальная информация приведена в работе A. Wróblewska (1992), которая, изучив особенности половой дифференциации данного вида на территории Польши, показала, что у C. aromaticum встречаются особи с двойными зонтиками на побегах I–III порядков ветвления, при этом доля обоеполых цветков уменьшается при увеличении порядка ветвления побега, на котором расположен двойной зонтик: от 17.9% в терминальных зонтиках до 11.2% в зонтиках на побегах III порядка ветвления. С.Е. Петрова (Petrova, 2016) показала наличие различий в размерах подстолбиев (нектарных дисков) у тычиночных и обоеполых цветков данного вида. Отсутствие детальных характеристик половой дифференциации у этого вида определило цель нашей работы – выявление особенностей гендерного полиморфизма у C. aromaticum на органном и организменном уровнях в Московской области.
МАТЕРИАЛ И МЕТОДЫ
При описании особенностей строения синфлоресценции у C. aromaticum мы использовали единую терминологию и систему обозначения для зонтиков, применяемую и для других представителей семейства Apiaceae (Kuznetzova, 1981; Kuznetzova, Timonin, 2017): простой зонтик; сложный, или двойной, зонтик, состоящий из простых зонтиков; терминальный двойной зонтик заканчивает главный побег.
Изучение половой дифференциации C. aromaticum проводили как в естественных условиях Московской области с 2017 по 2019 гг., так и в процессе камеральной обработки собранного материала. Основные исследования проведены на трех ценопопуляциях C. aromaticum. В ценопопуляции 1, расположенной в сероольшатнике бутеневом (окрестности п. Павловская Слобода, Московская обл.), проективное покрытие C. aromaticum достигало 50%. Среди других видов травяного покрова наиболее часто встречались Geranium sylvaticum L., Festuca gigantea (L.) Vill., Stachys sylvatica L., Campanula latifolia L. Ценопопуляция 2 расположена в Одинцовском р-не Московской обл., окр. г. Одинцово в составе осинника-сероольшатника недотрогового, в травяном покрове которого доминировали Impatiens noli-tangere L., Chaerophyllum aromaticum, Urtica dioica L., Dryopteris filix-mas (L.) Schott, Anthriscus sylvestris (L.) Hoffm., Geum urbanum L., Stachys sylvatica L. Ценопопуляция 3 расположена в Мытищинском р-не Московской обл., окр. п. Вешки в составе широколиственного леса (основные доминанты древесного яруса – Acer platanoides L. и Fraxinus excelsior L.) с злаково-разнотравным травостоем (доминировали – Stellaria holostea L., Asarum europaeum L., Asperula odorata L., Carex pilosa Scop., Aegopodium podagraria L., Equisetum sylvaticum L., Melica nutans L., Dryopteris lanceolato-cristata (Hoffm.) Alston). Для изучения морфологических особенностей обоеполых и тычиночных цветков их собирали с зонтиков на побегах разного порядка и фиксировали в смеси ледяной уксусной кислоты и абсолютного этилового спирта в пропорции 1:3. Для анализа использовали особи только средневозрастного генеративного онтогенетического состояния. В связи с тем, что обоеполые цветки C. aromaticum строго протандричны (разновременное созревание мужских и женских генеративных органов в цветке), часть цветков с зонтиков на побегах I–IV порядков ветвления фиксировали в тычиночной фазе развития (для изучения параметров околоцветника и андроцея). Оставшиеся цветки в зонтиках собирали уже в рыльцевой фазе развития (для исследования признаков гинецея). После этого в камеральных условиях с помощью стереоскопического микроскопа Биомед МС-1 с окуляр-микрометром при увеличении 20 или 40 определяли диаметр венчика, линейные параметры (длину и ширину) абаксиальных, трансверзальных и адаксиальных лепестков, тычинок, пыльников, абаксиального стилоподия, длину завязи, столбика и диаметр рыльца. С каждой особи собирали цветки с одного терминального двойного зонтика и трех двойных зонтиков на побегах следующих порядков ветвления. В пределах отдельного двойного зонтика измеряли по 30 обоеполых и 30 тычиночных цветков. При этом цветки брали из разных частей двойного зонтика: по 10 цветков каждой половой формы из периферических, срединных и внутренних простых зонтиков. В каждой из исследованных ценопопуляций двойные зонтики были собраны у 25 разных генеративных особей одного онтогенетического состояния. В целом изучено 3000 обоеполых и 3000 тычиночных цветков. В дальнейшем полученные данные усредняли с учетом расположения цветков в зонтиках на побегах разного порядка и степенью разветвленности синфлоресценций. Поскольку в ценопопуляциях 1 и 2 встречались особи с наибольшей степенью разветвленности синфлоресценций, в статье использованы данные по морфометрии цветков ценопопуляции 1. В двух других ценопопуляциях характер изменения размерных показателей признаков цветков на побегах разного порядка был аналогичным.
Для изучения морфологических и биологических особенностей пыльцы обоеполых и тычиночных цветков C. aromaticum использовали тот же фиксированный материал, собранный для исследования морфологических особенностей цветков. Из всех пыльников 5 обоеполых и 5 тычиночных цветков каждого простого зонтика брали пыльцу для приготовления препаратов. В каждом двойном зонтике анализировали по 5 простых зонтиков. Пыльцу окрашивали ацетокармином, что позволяет определить ее фертильность. Определялись длина полярной оси пыльцевых зерен и их экваториальный диаметр. На каждом препарате учитывали не менее 100 пыльцевых зерен. Пыльцу просматривали не менее, чем в 20 полях зрения, что зависело от плотности расположения пыльцевых зерен. Параллельно на этих же препаратах подсчитывали число фертильных и стерильных пыльцевых зерен. В дальнейшем полученные данные усредняли в соответствии с порядком ветвления побега, на котором формируются двойные зонтики.
В анализируемых ценопопуляциях в конце цветения C. aromaticum в фазе молочно-восковой спелости мерикарпиев закладывали трансекты шириной 1 м и подсчитывали число особей C. aromaticum, находящихся в средневозрастном генеративном онтогенетическом состоянии с разной степенью разветвленности синфлоресценций. В это время заканчивают цветение цветки в простых и двойных зонтиках на побегах самых высоких порядков, и происходит интенсивное развитие формирующихся семян и плодов, что позволяет оценить соотношение тычиночных и обоеполых цветков во всех простых и двойных зонтиках синфлоресценции. На трансекте изучали 100 особей данного вида. Поскольку двойные зонтики у C. aromaticum могут формироваться на побегах I–IV порядков, то есть существует три типа особей, различающиеся степенью разветвления их синфлоресценций, были отобраны по 10 особей каждого типа (с двойными зонтиками на побегах I–II, I–III и I–IV порядков). В камеральных условиях у этих 30 особей из каждой ценопопуляции определяли как диаметр двойного зонтика (на побегах всех порядков ветвления), длину лучей двойного зонтика, число простых зонтиков в составе отдельного двойного зонтика, диаметр простого зонтика, длину цветоножек, число обоеполых и тычиночных цветков. В каждом двойном зонтике в анализ включены все простые зонтики. В дальнейшем полученные данные для особей с одним вариантом разветвленности синфлоресценций усредняли для получения обобщенной характеристики таких особей. Аналогичным образом поступали для всех трех выявленных вариантов особей. Как и в случае с размерными показателями признаков цветков, в дальнейшем в статье обсуждаются характеристики частей синфлоресценций особей из ценопопуляции № 1, поскольку характер изменений структурных компонентов синфлоресценций в изученных ценопопуляциях идентичен.
Все полученные данные обработаны методами вариационной статистики (Sokal, Rolf, 2012). Для каждого изучаемого признака определяли пределы варьирования (min–max), среднее значение (М), его ошибку (m). Сравнение средних арифметических проводили с помощью t-критерия Стьюдента. Результаты вычислений представлены в таблицах и на рисунке.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Морфология обоеполых и тычиночных цветков. C. aromaticum – андромоноэцичное растение, у которого обоеполые и тычиночные цветки встречаются в двойных зонтиках на побегах разного порядка ветвления (табл. 1).
Таблица 1.
Морфологические параметры обоеполых и тычиночных цветков в зонтиках на побегах разного порядка у Chaerophyllum aromaticum (на примере особей ценопопуляции № 1 с зонтиками на побегах I–IV порядков) Table 1. Values of morphological traits of perfect and male flowers in umbels on the shoots of different orders in Chaerophyllum aromaticum (for individuals of population 1 with umbels on the shoots of different orders)
| Признак Morphological trait |
Цветок Flower |
Терми-
нальные зонтики Terminal umbels |
Зонтики на побегах II порядка Umbels on shoots of the 2nd order |
Зонтики на побегах III порядка Umbels on shoots of the 3rd order |
Зонтики на побегах IV порядка Umbels on shoots of the 4th order |
|---|---|---|---|---|---|
| Диаметр венчика, мм Corolla diameter, mm |
b | 6.11 ± 0.16 | 5.73 ± 0.10 | 4.61 ± 0.07 | 4.45 ± 0.04 |
| p | 4.23 ± 0.24 | 3.34 ± 0.21 | 3.18 ± 0.28 | 2.85 ± 0.11 | |
| Длина абаксиальных лепестков, мм Length of abaxial petals, mm |
b | 2.68 ± 0.09 | 2.52 ± 0.03 | 2.14 ± 0.04 | 1.82 ± 0.08 |
| p | 1.98 ± 0.14 | 1.60 ± 0.09 | 1.53 ± 0.13 | 1.17 ± 0.05 | |
| Ширина абаксиальных лепестков, мм Width of abaxial petals, mm |
b | 2.16 ± 0.08 | 1.87 ± 0.04 | 1.52 ± 0.04 | 1.46 ± 0.03 |
| p | 1.55 ± 0.08 | 1.09 ± 0.09 | 1.09 ± 0.13 | 1.00 ± 0.06 | |
| Длина трансверзальных лепестков, мм Length of transversal petals, mm |
b | 2.31 ± 0.06 | 2.30 ± 0.06 | 1.91 ± 0.03 | 1.66 ± 0.06 |
| p | 1.72 ± 0.08 | 1.45 ± 0.11 | 1.37 ± 0.14 | 1.03 ± 0.04 | |
| Ширина трансверзальных лепестков, мм Width of transversal petals, mm |
b | 1.71 ± 0.05 | 1.63 ± 0.05 | 1.34 ± 0.03 | 1.27 ± 0.01 |
| p | 1.21 ± 0.07 | 1.05 ± 0.07 | 0.92 ± 0.08 | 0.85 ± 0.03 | |
| Длина адаксиальных лепестков, мм Length of adaxial petals, mm |
b | 2.11 ± 0.09 | 1.99 ± 0.06 | 1.58 ± 0.06 | 1.34 ± 0.06 |
| p | 1.62 ± 0.10 | 1.31 ± 0.07 | 1.21 ± 0.12 | 1.01 ± 0.05 | |
| Ширина адаксиальных лепестков, мм Width of adaxial petals, mm |
b | 1.65 ± 0.02 | 1.35 ± 0.06 | 1.17 ± 0.02 | 1.15 ± 0.02 |
| p | 1.14 ± 0.09 | 0.94 ± 0.05 | 0.93 ± 0.05 | 0.85 ± 0.06 | |
| Длина тычинок, мм Stamen length, mm |
b | 2.35 ± 0.09 | 2.00 ± 0.03 | 1.96 ± 0.04 | 1.64 ± 0.13 |
| p | 2.32 ± 0.05 | 2.15 ± 0.01 | 1.94 ± 0.03 | 1.29 ± 0.07 | |
| Длина пыльников, мм Anther length, mm |
b | 0.59 ± 0.03 | 0.58 ± 0.01 | 0.52 ± 0.01 | 0.49 ± 0.02 |
| p | 0.60 ± 0.01 | 0.57 ± 0.01 | 0.48 ± 0.01 | 0.47 ± 0.02 | |
| Ширина пыльников, мм Anther width, mm |
b | 0.47 ± 0.01 | 0.46 ± 0.01 | 0.46 ± 0.01 | 0.43 ± 0.02 |
| p | 0.51 ± 0.01 | 0.47 ± 0.01 | 0.46 ± 0.01 | 0.42 ± 0.01 | |
| Длина полярной оси пыльцевых зерен, мкм Length of polar axe of pollen, μm |
b | 25.35 ± 0.45 | 24.12 ± 0.42 | 24.12 ± 0.45 | 25.25 ± 0.74 |
| p | 23.75 ± 0.35 | 25.00 ± 0.79 | 25.50 ± 0.91 | 24.50 ± 0.45 | |
| Экваториальный диаметр пыльцевых зерен, мкм Equatorial diameter of pollen, μm |
b | 14.25 ± 0.45 | 13.25 ± 0.28 | 13.50 ± 0.22 | 14.00 ± 0.42 |
| p | 14.00 ± 0.42 | 14.18 ± 0.65 | 14.14 ± 0.68 | 14.23 ± 0.42 | |
| Фертильность пыльцы, % Pollen fertility, % |
b | 95.6 ± 0.8 | 95.2 ± 1.1 | 94.8 ± 0.8 | 94.6 ± 0.5 |
| p | 95.4 ± 1.1 | 94.4 ± 1.0 | 94.2 ± 1.2 | 95.2 ± 1.3 | |
| Длина абаксиального стилоподия, мм Abaxial stylopodium length, mm |
b | 0.96 ± 0.05 | 0.92 ± 0.02 | 0.80 ± 0.03 | 0.68 ± 0.02 |
| p | 0.37 ± 0.02 | 0.32 ± 0.01 | 0.27 ± 0.03 | 0.23 ± 0.04 | |
| Высота абаксиального стилоподия, мм Abaxial stylopodium height, mm |
b | 1.56 ± 0.06 | 1.42 ± 0.06 | 1.29 ± 0.05 | 1.08 ± 0.02 |
| p | 0.65 ± 0.02 | 0.56 ± 0.02 | 0.50 ± 0.05 | 0.42 ± 0.05 | |
| Длина завязи, мм Ovary length, mm |
b | 2.34 ± 0.10 | 1.66 ± 0.03 | 1.25 ± 0.04 | 0.94 ± 0.03 |
| Длина столбика, мм Stylodium length, mm |
b | 1.67 ± 0.08 | 1.10 ± 0.03 | 0.69 ± 0.02 | 0.47 ± 0.02 |
| Диаметр рыльца, мм Stigma diameter, mm |
b | 0.21 ± 0.01 | 0.19 ± 0.01 | 0.17 ± 0.01 | 0.17 ± 0.01 |
Обоеполые цветки C. aromaticum традиционно относят к актиноморфным (внутренние цветки в простых зонтиках) и зигоморфным (краевые цветки в простых зонтиках) (Shishkin, 1950; Pimenov, Ostroumova, 2012). Однако, ревизия представлений о симметрии цветков представителей порядка Apiales (Nuraliev et al., 2019) показала, что многие виды семейства Apiaceae характеризуются моносимметричными цветками. При этом зигоморфия венчика краевых цветков в простых зонтиках связана с увеличением одного лепестка целиком и примыкающих к нему половинок двух соседних лепестков. Это самый распространенный вариант формирования зигоморфии венчиков в семействе Apiaceae (Froebe, Ulrich, 1978). Однако, если принимать во внимание особенности строения всех частей цветков, то как краевые, так и внутренние цветки в простых зонтиках C. aromaticum правильнее относить к зигоморфным, несмотря на видимые различия в структуре их венчиков. Элементы обоеполых цветков C. aromaticum расположены в 4 кругах. В чашечке, венчике и андроцее круги 5-членные, и в гинецее – 2-членные. Околоцветник двойной, его части прикрепляются, как и тычинки, к верхней части завязи. Чашечка незаметная, представлена небольшими зубцами. Венчик состоит из пяти лепестков, белых, голых, свободных, цельных, обратнояйцевидной формы, на верхушке до 1/3 двулопастных, в выемке с загнутой внутрь долькой, при основании клиновидных или сразу переходящих в короткий ноготок. Наиболее крупные размеры характерны для абаксиальных лепестков, наиболее мелкие – для адаксиальных, при этом лепестки в трансверзальном положении занимают промежуточное положение между этими двумя лепестками (табл. 1). Тычиночные нити длинные, в бутоне загнутые внутрь цветка. Пыльники желто-зеленого цвета, продолговатые, эллиптические, раскрываются продольно щелями. Пыльники прикрепляются тыльной частью к тычиночным нитям, содержат четыре пыльцевых гнезда, которые сближены попарно в 2 теки. Пыльцевые зерна 3-бороздно-оровые, продолговатые, эллипсоидальные, в экваториальной зоне сжатые. Фертильность пыльцы выше 94% (табл. 1). Гинецей синкарпный, состоит из двух плодолистиков, завязь двугнездная, содержащая в каждом гнезде по одному развитому семязачатку. Стилодиев два, они имеют расширенные железистые подстолбия-нектарники, или стилоподии, у своего основания. В недавно раскрывшемся цветке стилодии короткие, плотно прижаты к спинной стороне подстолбиев, после опадения пыльников – выпрямленные и вытянутые. Рыльца головчатые. Завязь полунижняя, все части цветка прикреплены ниже подстолбиев, составляющих верхушку завязи.
Интересно отметить, что абаксиальные и адаксиальные подстолбии в обоеполых цветках различаются по размерам. Так, средние размеры абаксиальных подстолбиев составляют 0.96 мм, а адаксиальных – 0.83 мм (t = 2.670 при P = 0.09). Учитывая особенности пространственного расположения тычинок относительно гинецея и различия размеров абаксиальных и адаксиальных подстолбиев, через обоеполые цветки в итоге можно провести только одну плоскость симметрии. Таким образом, цветки C. aromaticum обладают зигоморфным строением даже без учета размерных отличий абаксиальных лепестков у краевых цветков в простых зонтиках.
Сравнение средних значений признаков различных частей обоеполых цветков, расположенных в зонтиках на побегах разного порядка, показывает следующее. Во-первых, наблюдаются достоверные различия между обоеполыми цветками в зонтиках на побегах I–IV порядков ветвления по таким признакам как диаметр венчика, длина тычинок, размеры стилоподия, длина завязи и диаметр рыльца. Все эти параметры закономерно уменьшаются у обоеполых цветков при переходе от зонтиков на побегах I порядка ветвления к зонтикам на побегах IV порядка. Остальные признаки также уменьшаются в этом направлении, но их различия недостоверны. Во-вторых, наиболее существенные изменения размеров обоеполых цветков и их частей наблюдаются между цветками, расположенными в зонтиках на побегах II и III порядков ветвления: по всем исследованным признакам за исключением ширины пыльников выявлены достоверные различия. Следовательно, диаметр венчика и размеры женских генеративных органов за исключением диаметра рыльца зависят от положения обоеполых цветков в зонтиках на побегах разного порядка ветвления. В-третьих, не выявлено достоверных отличий по размерам частей обоеполых цветков, занимающих разное положение в простых зонтиках. Другими словами, центральные обоеполые цветки характеризуются такими же размерами, как и обоеполые цветки, располагающиеся по периферии простого зонтика. Подобная закономерность наблюдается в двойных зонтиках на побегах любого порядка ветвления. Следует отметить, что какой-либо строгой закономерности изменения размеров пыльцевых зерен и фертильности пыльцы между обоеполыми цветками в двойных зонтиках на побегах разного порядка ветвления не выявлено. Следовательно, даже в двойных зонтиках на побегах самых высоких порядков уменьшение размеров пыльцевых зерен у C. aromaticum не отмечается.
Структурных различий между обоеполыми и тычиночными цветками не наблюдается по особенностям строения околоцветника и андроцея. Однако в связи с редукцией гинецея в тычиночных цветках C. aromaticum отсутствуют завязи, а стилоподии значительно уменьшились в размерах. Сравнение показателей разных частей обоеполых и тычиночных цветков в зонтиках на побегах одного порядка ветвления показало следующее. По всем изученным параметрам околоцветника и стилоподиев наблюдаются достоверные различия между обоеполыми и тычиночными цветками в зонтиках на побегах одного порядка ветвления: размеры частей обоеполых цветков всегда больше, чем у тычиночных. Исключение составляют параметры андроцея и пыльцы. По таким признакам андроцея, как длина тычинок и размеры пыльников, довольно часто не отмечается достоверных различий между двумя половыми типами цветков.
В целом тычиночные цветки и их составные части закономерно уменьшаются при переходе от зонтиков, расположенных на главных побегах, к зонтикам на побегах IV порядка ветвления. Особенно значительные различия в размерах частей наблюдаются между тычиночными цветками в терминальных двойных зонтиках и двойных зонтиках на побегах II порядка ветвления. При переходе от двойных зонтиков на побегах II порядка к двойным зонтикам на побегах III и IV порядков отмечается уменьшение размеров частей тычиночных цветков, но эти различия не всегда достоверны. Исключение составляют размеры пыльцевых зерен и фертильности пыльцы, которые закономерно не изменяются при переходе от терминальных двойных зонтиков к двойным зонтикам на побегах IV порядка ветвления (табл. 1).
Уменьшение размеров тычиночных цветков и их частей в сравнении с обоеполыми цветками в пределах простого зонтика, обусловлено как минимум двумя причинами. С одной стороны, в целом миниатюризация цветков в простых зонтиках в центростремительном направлении – общая особенность, характерная для многих изученных представителей семейств Apiaceae и Araliaceae. Большинство исследователей объясняют такое изменение размеров цветков и их частей градиентом доступности веществ, необходимых для образования и развития цветков, особенно обоеполых (Knuth, 1898; Braak, Kho, 1958; Quagliotti, 1967; Reuther, Claßen-Bockhoff, 2010). Этим же, по мнению большинства авторов, обусловлено более частое расположение тычиночных цветков в центре простых зонтиков, а обоеполых цветков – по его периферии. С другой стороны, рядом авторов высказана так называемая гипотеза “защитной функции” (Bawa, Opler, 1975; Delph et al., 1996), согласно которой цветки, образующие плоды и семена (у C. aromaticum это обоеполые цветки), как правило, имеют более крупные размеры, необходимые для защиты генеративных органов цветка и развивающихся плодов, чем у тычиночных цветков.
Наличие четких структурных и размерных различий обоеполых и тычиночных цветков C. aromaticum позволяет достаточно легко проанализировать особенности их расположения в зонтиках на побегах разного порядка ветвления и выявить связанные с этим общие закономерности строения синфлоресценций данного вида (табл. 2).
Таблица 2.
Морфометрические показатели простых и двойных зонтиков на побегах разного порядка у Chaerophyllum aromaticum Table 2. Values of Chaerophyllum aromaticum morphological traits of umbels and umbellets on shoots of different order
| Признак/Morphological trait | Min–max | M ± m |
|---|---|---|
| Особи с зонтиками на побегах I и II порядков/Individuals with umbels on shoots of 1st and 2nd orders | ||
| Терминальные двойные зонтики/Terminal double umbels | ||
| Диаметр двойного зонтика, см Diameter of double umbel, cm |
5.7–10.5 | 8.6 ± 1.2 |
| Длина лучей двойного зонтика, см Ray length in double umbel, cm |
1.8–5.2 | 3.1 ± 0.5 |
| Число двойных зонтиков Number of double umbels |
1 | 1 |
| Число простых зонтиков в двойном зонтике Number of umbellets per double umbel |
13–22 | 17.0 ± 2.2 |
| Диаметр простого зонтика, см Umbellet diameter, cm |
1.6–2.8 | 2.3 ± 0.3 |
| Длина цветоножек, см Pedicel length, cm |
0.3–0.9 | 0.6 ± 0.1 |
| Число цветков в простых зонтиках Number of flowers per umbellet |
17–20 | 18.7 ± 0.7 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellet, % |
61.1–94.1 | 78.9 ± 3.1 |
| Двойные зонтики на побегах II порядка Double umbels on shoots of the 2nd order | ||
| Диаметр двойного зонтика, см Diameter of double umbel, cm |
5.3–8.6 | 7.0 ± 0.5 |
| Длина лучей двойного зонтика, см Ray length in double umbel, cm |
1.3–4.0 | 2.3 ± 0.3 |
| Число двойных зонтиков Number of double umbels |
1–6 | 3.0 ± 0.2 |
| Число простых зонтиков в двойном зонтике Number of umbellets per double umbel |
15–21 | 17.3 ± 0.8 |
| Диаметр простого зонтика, см Umbellet diameter, cm |
1.3–2.4 | 1.9 ± 0.2 |
| Длина цветоножек, см Pedicel length, cm |
0.4–0.7 | 0.5 ± 0.03 |
| Число цветков в простых зонтиках Number of flowers per umbellet |
21–29 | 24.3 ± 1.1 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellet, % |
76.7–96.1 | 87.5 ± 1.9 |
| Особи с зонтиками на побегах I–III порядков/Individuals with umbels on shoots of 1st, 2nd and 3rd orders | ||
| Терминальные двойные зонтики/Terminal double umbels | ||
| Диаметр двойного зонтика, см Diameter of double umbel, cm |
6.0–10.5 | 8.5 ± 0.9 |
| Длина лучей двойного зонтика, см Ray length in double umbel, cm |
2.4–5.6 | 4.0 ± 0.4 |
| Число двойных зонтиков Number of double umbels |
1 | 1 |
| Число простых зонтиков в двойном зонтике Number of umbellets per double umbel |
11–14 | 12.3 ± 0.5 |
| Диаметр простого зонтика, см Umbellet diameter, cm |
2.0–3.8 | 2.9 ± 0.3 |
| Длина цветоножек, см Pedicel length, cm |
0.5–1.2 | 0.8 ± 0.08 |
| Число цветков в простых зонтиках Number of flowers per umbellet |
19–22 | 20.8 ± 0.2 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellet, % |
57.1–86.7 | 73.9 ± 2.0 |
| Двойные зонтики на побегах II порядка/Double umbels on shoots of the 2nd order | ||
| Диаметр двойного зонтика, см Diameter of double umbel, cm |
6.5–11.0 | 9.0 ± 0.4 |
| Длина лучей двойного зонтика, см Ray length in double umbel, cm |
1.5–5.0 | 3.1 ± 0.2 |
| Число двойных зонтиков Number of double umbels |
2–7 | 4.3 ± 0.1 |
| Число простых зонтиков в двойном зонтике Number of umbellets per double umbel |
13–19 | 15.9 ± 0.4 |
| Диаметр простого зонтика, см Umbellet diameter, cm |
1.8–3.6 | 2.4 ± 0.2 |
| Длина цветоножек, см Pedicel length, cm |
0.4–1.0 | 0.7 ± 0.03 |
| Число цветков в простых зонтиках Number of flowers per umbellet |
20–28 | 24.3 ± 0.8 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellet, % |
63.6–98.6 | 82.4 ± 1.4 |
| Двойные зонтики на побегах III порядка/Double umbels on shoots of the 3rd order | ||
| Диаметр двойного зонтика, см Diameter of double umbel, cm |
4.7–8.0 | 6.7 ± 0.3 |
| Длина лучей двойного зонтика, см Ray length in double umbel, cm |
1.1–3.5 | 2.2 ± 0.2 |
| Число двойных зонтиков Number of double umbels |
1–9 | 4.2 ± 0.3 |
| Число простых зонтиков в двойном зонтике Number of umbellets per double umbel |
10–16 | 14.7 ± 0.5 |
| Диаметр простого зонтика, см Umbellet diameter, cm |
1.1–1.9 | 1.6 ± 0.1 |
| Длина цветоножек, см Pedicel length, cm |
0.3–0.8 | 0.5 ± 0.03 |
| Число цветков в простых зонтиках Number of flowers per umbellet |
19–24 | 22.2 ± 0.5 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellet, % |
78.9–98.5 | 93.2 ± 1.0 |
| Особи с зонтиками на побегах I–IV порядков/Individuals with umbels on shoots of 1st – 4th orders | ||
| Терминальные двойные зонтики/Terminal double umbels | ||
| Диаметр двойного зонтика, см Diameter of double umbel, cm |
11.5–14.0 | 12.8 ± 0.9 |
| Длина лучей двойного зонтика, см Ray length in double umbel, cm |
3.0–6.2 | 4.6 ± 0.8 |
| Число двойных зонтиков Number of double umbels |
1 | 1 |
| Число простых зонтиков в двойном зонтике Number of umbellets per double umbel |
16–17 | 16.5 ± 0.4 |
| Диаметр простого зонтика, см Umbellet diameter, cm |
3.4–3.5 | 3.5 ± 0.04 |
| Длина цветоножек, см Pedicel length, cm |
0.7–1.3 | 1.0 ± 0.1 |
| Число цветков в простых зонтиках Number of flowers per umbellet |
19–21 | 20.0 ± 0.7 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellet, % |
31.6–53.3 | 42.5 ± 0.6 |
| Двойные зонтики на побегах II порядка/Double umbels on shoots of the 2nd order | ||
| Диаметр двойного зонтика, см Diameter of double umbel, cm |
8.0–10.2 | 8.7 ± 0.3 |
| Длина лучей двойного зонтика, см Ray length in double umbel, cm |
1.5–4.8 | 3.2 ± 0.4 |
| Число двойных зонтиков Number of double umbels |
4–6 | 4.9 ± 0.3 |
| Число простых зонтиков в двойном зонтике Number of umbellets per double umbel |
16–21 | 17.7 ± 0.7 |
| Диаметр простого зонтика, см Umbellet diameter, cm |
2.0–2.2 | 2.1 ± 0.03 |
| Длина цветоножек, см Pedicel length, cm |
0.5–1.0 | 0.7 ± 0.04 |
| Число цветков в простых зонтиках Number of flowers per umbellet |
21–26 | 23.8 ± 0.6 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellet, % |
60.9–79.2 | 64.9 ± 1.3 |
| Двойные зонтики на побегах III порядка/Double umbels on shoots of the 3rd order | ||
| Диаметр двойного зонтика, см Diameter of double umbel, cm |
6.5–7.7 | 7.1 ± 0.1 |
| Длина лучей двойного зонтика, см Ray length in double umbel, cm |
1.3–3.0 | 2.1 ± 0.2 |
| Число двойных зонтиков Number of double umbels |
3–9 | 7.1 ± 0.9 |
| Число простых зонтиков в двойном зонтике Number of umbellets per double umbel |
17–22 | 19.2 ± 0.8 |
| Диаметр простого зонтика, см Umbellet diameter, cm |
1.5–2.2 | 1.8 ± 0.1 |
| Длина цветоножек, см Pedicel length, cm |
0.4–0.8 | 0.6 ± 0.03 |
| Число цветков в простых зонтиках Number of flowers per umbellet |
20–26 | 22.2 ± 0.8 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellet, % |
76.2–91.8 | 88.9 ± 0.9 |
| Двойные зонтики на побегах IV порядка/Double umbels on shoots of the 4th order | ||
| Диаметр двойного зонтика, см Diameter of double umbel, cm |
3.8–5.0 | 4.6 ± 0.2 |
| Длина лучей двойного зонтика, см Ray length in double umbel, cm |
0.8–2.1 | 1.5 ± 0.1 |
| Число двойных зонтиков Number of double umbels |
2–5 | 2.9 ± 0.4 |
| Число простых зонтиков в двойном зонтике Number of umbellets per double umbel |
11–15 | 13.4 ± 0.6 |
| Диаметр простого зонтика, см Umbellet diameter, cm |
1.1–1.3 | 1.2 ± 0.04 |
| Длина цветоножек, см Pedicel length, cm |
0.3–0.6 | 0.5 ± 0.03 |
| Число цветков в простых зонтиках Number of flowers per umbellet |
14–17 | 16.2 ± 0.5 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellet, % |
82.4–99.1 | 97.6 ± 1.7 |
У C. aromaticum синфлоресценция метельчатого типа, а флоральной единицей является открытый совершенный двойной зонтик. Согласно данным М.Г. Пименова и Т.А. Остроумовой (Pimenov, Ostroumova, 2012), двойные зонтики лишены обертки или она состоит из нескольких рано опадающих по краю реснитчатых листочков с нитевидно оттянутой верхушкой. Простые зонтики с оберточками из 5–10 широколанцетных, по краю широко пленчатых, реснитчатых листочков с длинным кончиком. Простые зонтики, входящие в состав двойных зонтиков, у представителей рода Chaerophyllum всегда закрытые, поскольку имеют терминальный цветок, который всегда обоеполый (Kuznetzova, 1982). Кроме центрального обоеполого цветка, обоеполые цветки в простых зонтиках всегда располагаются по периферии зонтика, а между обоеполыми цветками располагаются тычиночные цветки (Knuth, 1898; Spalik, Downie, 2001). Однако, согласно нашим исследованиям, на побегах высоких порядков в центре двойных зонтиков часто встречаются простые зонтики, у которых сохраняется центральный обоеполый цветок, а периферическими являются тычиночные цветки из-за полной редукции краевых обоеполых цветков.
Изучение особенностей строения синфлоресценций у C. aromaticum в Московской области показало, что у данного вида выделяются три типа особей по степени разветвленности: 1) особи с двойными зонтиками, расположенными на побегах I и II порядков ветвления; 2) особи с двойными зонтиками на побегах I, II и III порядков ветвления; 3) особи с двойными зонтиками на побегах I, II, III и IV порядков ветвления. При этом соотношение особей с разной степенью разветвленности синфлоресценций различается в изученных нами ценопопуляциях (рис. 1). Во всех исследованных ценопопуляциях заметно преобладают особи, у которых двойные зонтики располагались на побегах I, II и III порядков ветвления (65–70.0% от числа изученных особей). Участие особей с двойными зонтиками на побегах IV порядка ветвления зависит от фитоценотических условий. В сероольшатниках (ценопопуляции 1 и 2) встречаются все три морфологических типа особей, различающиеся степенью разветвленности побеговой системы. В широколиственном лесу (ценопопуляция 3) особи с двойными зонтиками на побегах IV порядка ветвления нами не выявлены (рис. 1). Следовательно, у C. aromaticum формируется как минимум два типа синфлоресценций: кисть из двойных зонтиков (у особей с двойными зонтиками, расположенными на побегах I и II порядков ветвления) или метелка из двойных зонтиков (у особей с двойными зонтиками на побегах I–III или I–IV порядков ветвления). Таким образом, нами выявлен более широкий спектр степени разветвления синфлоресценции C. aromaticum, чем на территории Польши (Wróblewska, 1992).
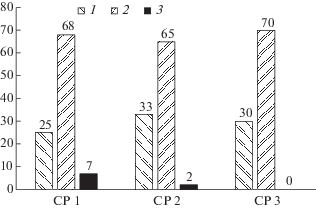
Рис. 1.
Соотношение особей с разной степенью разветвленности синфлоресценций у Chaerophyllum aromaticum в трех ценопопуляциях.
По оси абсцисс – изученные ценопопуляции и типы особей с разной степенью разветвленности синфлоресценций, по оси ординат – доля (в %) особей с разной степенью разветвленности синфлоресценций.
CP 1–CP 3 – изученные ценопопуляции, 1 – особи с двойными зонтиками на побегах I–II порядков ветвления, 2 – особи с двойными зонтиками на побегах I–III порядков ветвления, 3 – особи с двойными зонтиками на побегах I–IV порядков ветвления.
Fig. 1. The ratio of individuals with different degrees of synflorescence branching in three populations of Chaerophyllum aromaticum.
X-axis – studied populations and types of individuals with different degrees of synflorescence branching; Y-axis – share of individuals with different degrees of synflorescence branching (%).
CP 1–CP 3 – studied populations, 1 – individuals with double umbels on the shoots of 1st and 2nd orders, 2 – individuals with double umbels on the shoots of 1st, 2nd, and 3rd orders, 3 – individuals with double umbels on the shoots of 1st, 2nd, 3rd, and 4th orders.
На главном побеге у C. aromaticum, как и других представителей рода Chaerophyllum, всегда формируется единственный терминальный двойной зонтик – как правило, самый крупный из двойных зонтиков синфлоресценции. Его диаметр у особей с зонтиками на побегах I и II порядков ветвления варьирует от 5.7 до 10.5 см, при этом среднее число простых зонтиков в его составе – 17.0 (табл. 2). Простые зонтики располагаются в 3–4 кругах в пределах двойного зонтика, при этом их размеры и число цветков в них закономерно уменьшаются от краевых самых крупных с максимальным числом цветков к центральным самым мелким с минимальным числом цветков. Доля тычиночных цветков в простых зонтиках терминального двойного зонтика довольно высока и варьирует от 69.6% до 93.8%. При этом доля тычиночных цветков закономерно увеличивается при переходе от краевых простых зонтиков к центральным простым зонтикам в составе двойного зонтика.
Двойные зонтики на побегах II порядка ветвления в среднем меньше по диаметру, длине лучей двойного зонтика и диаметру простого зонтика в сравнении с терминальным двойным зонтиком. В состав кисти из двойных зонтиков у таких особей может входить от 1 до 6 двойных зонтиков на побегах II порядка. По сравнению с терминальными двойными зонтиками в зонтиках на побегах II порядка увеличивается число цветков и доля тычиночных цветков в простых зонтиках. При этом наблюдается та же закономерность, что и в терминальных двойных зонтиках: увеличение доли тычиночных цветков у центральных простых зонтиков по сравнению с краевыми простыми зонтиками. Уменьшение размеров и одновременное увеличение числа образующихся в них цветков у простых зонтиков, входящих в состав двойных зонтиков на побегах II порядка, по сравнению с простыми зонтиками терминального двойного зонтика, по нашему мнению, обусловлено увеличением числа тычиночных цветков, которые характеризуются меньшими размерами и в связи с этим более компактным пространственным расположением.
Следовательно, у особей с двойными зонтиками на побегах I и II порядков ветвления формируется синфлоресценция, представленная кистью из двойных зонтиков, отмечается уменьшение многих показателей морфологических признаков при переходе от терминальных двойных зонтиков к двойным зонтикам на побегах II порядка, а также довольно высокая доля тычиночных цветков как в терминальных зонтиках, так и в зонтиках на побегах II порядка.
Особи, у которых двойные зонтики расположены на побегах I, II и III порядков, составляют 65–70.0% от общей численности. У них формируется метелка из двойных зонтиков. При этом наиболее мощные и разветвленные боковые побеги II порядка характерны для средней части главного побега. Т.е. самые нижние и самые верхние боковые побеги II порядка оказываются менее разветвленными и могут нести лишь одиночные двойные зонтики, тогда как боковые побеги II порядка, расположенные в средней части главного побега, самые разветвленные, и на них образуется максимальное число двойных зонтиков на побегах следующего порядка ветвления.
Главный побег у таких особей заканчивается двойным зонтиком довольно крупным, но не самым большим в синфлоресценции: двойные зонтики на побегах II порядка сопоставимы по размерам, а иногда незначительно, но недостоверно (P = 0.613) могут превышать терминальный двойной зонтик (табл. 2). Если сравнивать терминальные двойные зонтики у особей с двумя и тремя порядками ветвления побеговой системы, видно, что с увеличением степени разветвленности доля тычиночных цветков в них уменьшается.
Число двойных зонтиков на побегах II порядка ветвления у такого типа особей варьирует от 2 до 7, но наиболее часто их 3–5. По сравнению с терминальными двойными зонтиками в двойных зонтиках на побегах II порядка увеличивается число простых зонтиков (до 13–19), число цветков в них (до 20–28) и доля тычиночных цветков (до 63.6–98.6%) (табл. 2). В пределах этих двойных зонтиков также наблюдается уменьшение числа цветков и возрастание доли тычиночных цветков при переходе от краевых простых зонтиков к центральным.
Число двойных зонтиков в синфлоресценции на побегах III порядка ветвления у таких особей может достигать 9 (чаще 4–5). При этом наблюдается уменьшение всех их параметров за исключением доли тычиночных цветков, которая у самых внутренних простых зонтиков может достигать 98.5%.
Если морфологические показатели двойных зонтиков не находятся в прямой зависимости от порядка ветвления побега, на котором они образуются, то доля тычиночных цветков напрямую зависит от этого параметра. С одной стороны, наиболее крупные, мощно развитые двойные зонтики и их структурные компоненты, как правило, характерны для побегов II порядка. Самые мелкие двойные зонтики и их элементы формируются на побегах III порядка. С другой стороны, доля тычиночных цветков закономерно увеличивается при возрастании порядка побега, на котором располагаются двойные зонтики. Тем не менее в пределах двойного зонтика также наблюдается увеличение доли тычиночных цветков в направлении от периферии двойного зонтика к его центру.
Самый редкий вариант образуют особи с двойными зонтиками, формирующимися на побегах I–IV порядков ветвления: их доля составляет всего 7% (в ценопопуляции 1) и 2% (в ценопопуляции 2). В ценопопуляции 3 особей с двойными зонтиками на побегах IV порядка ветвления не выявлено. Синфлоресценция таких особей представляет собой также метелку из двойных зонтиков, где, как правило, наиболее разветвленные боковые побеги II порядка располагаются в средней части главного побега. Следует отметить, что такой степени разветвленности синфлоресценций, согласно литературным данным, не отмечено в условиях Польши (Wróblewska, 1992).
Данный вариант синфлоресценций по целому ряду показателей значительно отличается от ранее рассмотренных вариантов. У особей с двойными зонтиками на побегах I–IV порядков ветвления терминальный двойной зонтик – самый крупный в синфлоресценции по таким параметрам как диаметр, длина лучей, диаметр простых зонтиков и длина цветоножек. В отличие от всех рассмотренных ранее вариантов строения синфлоресценции у особей с наличием четырех порядков ветвления побегов отмечается преобладание обоеполых цветков над тычиночными или их равное участие как в простых, так и в целом в терминальном двойном зонтике: доля тычиночных цветков варьирует от 31.6 до 53.3%.
Число двойных зонтиков на побегах II порядка ветвления варьирует от 4 до 6. Зонтики на побегах II порядка в отличие от терминальных двойных зонтиков характеризуются уменьшением большинства размерных характеристик за исключением числа простых зонтиков, числа цветков в них и долей тычиночных цветков. Эти показатели увеличиваются при переходе от терминальных к двойным зонтикам на побегах II порядка. Тычиночные цветки начинают преобладать в составе простых и двойных зонтиков. Кроме того, также наблюдается увеличение доли тычиночных цветков в направлении от периферических к центральным простым зонтикам в пределах двойного зонтика.
Двойные зонтики на побегах III порядка ветвления встречаются в числе от 3 до 9, но наиболее широко распространенный вариант – 8–9 двойных зонтиков. За исключением числа простых зонтиков в двойном зонтике и доли тычиночных цветков все остальные изученные параметры закономерно уменьшаются при переходе от двойных зонтиков на побегах II порядка к побегам III порядка (табл. 2). Увеличение числа простых зонтиков в составе двойных зонтиков, по нашему мнению, связано с возрастающей частотой встречаемости тычиночных цветков, входящих в простые зонтики.
Число двойных зонтиков на побегах IV порядка ветвления может варьировать от 2 до 5, но наиболее широко представленный вариант – 2–3 двойных зонтика. По всем исследованным морфологическим признакам двойные зонтики на побегах IV порядка меньше, чем двойные зонтики на побегах III порядка ветвления (табл. 2). Исключение составляет только доля тычиночных цветков в простых зонтиках, которая закономерно увеличивается и достигает 99.1% у простых зонтиков, расположенных в центре двойных зонтиков. Необходимо отметить, что у многих таких внутренних простых зонтиков уменьшение числа обоеполых цветков происходит за счет краевых обоеполых цветков, при этом центральный обоеполый цветок в простом зонтике всегда сохраняется, хотя он может быть и единственным среди тычиночных цветков.
Таким образом, у особей C. aromaticum с самыми разветвленными синфлоресценциями подавляющее число изученных параметров напрямую зависит от порядка побега, на котором формируются двойные зонтики. За исключением числа простых зонтиков в составе двойного зонтика и доли тычиночных цветков, все остальные признаки уменьшаются с повышением порядка ветвления побега, несущего двойные зонтики. Отмечается закономерное увеличение доли тычиночных цветков, которая минимальна в терминальных двойных зонтиках и максимальна в двойных зонтиках на побегах IV порядка.
В ряду особей, расположенных в порядке увеличения степени разветвленности побеговой системы, участие тычиночных цветков в двойных зонтиках на побегах одного порядка закономерно уменьшается (табл. 2). Например, при переходе от особей с двойными зонтиками на побегах I–II порядков ветвления к особям с двойными зонтиками на побегах I–IV порядков ветвления доля тычиночных цветков в терминальных двойных зонтиках снижается с 78.9% до 42.5% соответственно. Такая же закономерность проявляется при сравнении любых двойных зонтиков на побегах одного порядка ветвления у особей с разной структурой синфлоресценций.
Сравнение полученных нами данных с литературными источниками показывает следующее. Во-первых, в условиях Московской области выявлен более широкий спектр степени разветвленности синфлоресценций и образование трех типов особей по этому показателю. В исследованиях A. Wróblewska (1992) C. aromaticum обладал двойными зонтиками, формирующимися на побегах I–III порядков ветвления. В изученных нами условиях у этого вида двойные зонтики развиваются на побегах I–IV порядков ветвления. По всей видимости, большая степень разветвленности синфлоресценций этого вида в Московской области связана с тем, что две ценопопуляции (1 и 2) изучены в фитоценотическом оптимуме C. aromaticum (Vasilevich, 1998). Именно с этим мы связываем отсутствие особей с двойными зонтиками на побегах IV порядка ветвления в ценопопуляции № 3, изученной в условиях широколиственного леса. У C. aromaticum, как вероятно и у других видов этого рода, уже в терминальных двойных зонтиках отмечается высокая доля тычиночных цветков: 73.9% у особей с двойными зонтиками на побегах I–III порядков ветвления. У видов других родов, например, у Anthriscus caucalis M. Bieb., Angelica ampla A. Nelso и Cicuta douglasii (DC.) J.M. Coult. et Rose терминальные зонтики включают только обоеполые цветки, а у Aegopodium podagraria L., Conioselinum scopulorum (A. Gray) J.M. Coult. et Rose и Sium suave Walter в них формируются единичные тычиночные цветки (Schlessman, Barrie, 2004; Reuther, Claßen-Bockhoff, 2010; Godin et al., 2019). По всей видимости, такая высокая частота встречаемости тычиночных цветков уже в терминальных двойных зонтиках заключается в специфических особенностях андромоноэции у видов Chaerophyllum L.
Увеличение доли тычиночных цветков при возрастании порядка ветвления побега, на котором образуются зонтики, у представителей сем. Apiaceae известно довольно давно (Knuth, 1898; Doust, 1980; Webb, 1981; Dem’yanova, 2000). Однако cогласно исследованиям M.A. Schlessman и F.R. Barrie (2004), у протогиничных видов этого семейства отмечается противоположная ситуация, когда при возрастании порядка побега доля тычиночных цветков закономерно уменьшается. Вполне возможно, что такие разнонаправленные тенденции обусловлены особенностями развития цветков, а именно дихогамией. В исследованиях Wróblewska (1992) на примере C. aromaticum выявлено закономерное увеличение частоты тычиночных цветков при переходе от двойных зонтиков на главном побеге к двойным зонтикам на побегах III порядков. Однако согласно данным этого автора различия между двойными зонтиками на побегах I–II порядков по доли тычиночных цветков незначительны и недостоверны: 82.1 и 82.3%. По нашему мнению, такие несущественные отличия обусловлены тем, что A. Wróblewska объединила данные по доли тычиночных цветков у всех исследованных ею особей с разными вариантами разветвленности синфлоресценций. Мы долю тычиночных цветков определяли отдельно для каждого типа особей по степени ветвления их синфлоресценций. В результате у каждого типа особей выявлено закономерное увеличение частоты тычиночных цветков при возрастании порядка ветвления побега, на котором развиваются двойные зонтики (табл. 2). Увеличение частоты встречаемости тычиночных цветков с возрастанием порядка ветвления побега, на котором расположены двойные зонтики, многие исследователи рассматривают как адаптивный признак зонтичных (Dem’yanova, 2000; Reuther, Claßen-Bockhoff, 2010; Godin, Perkova, 2017; Endress, 2020). В сочетании с межцветковой протандрией и определенной последовательностью цветения двойных зонтиков в синфлоресценции преобладающие в двойных зонтиках на побегах высоких порядков ветвления тычиночные цветки служат донорами пыльцы для обеспечения ксеногамии в популяциях многих зонтичных. Когда раскрываются тычиночные цветки в двойных зонтиках на побегах высоких порядков, их пыльца не может опылять цветки двойных зонтиков на побегах предыдущего порядка, поскольку у них рыльцевая стадия уже закончена. Следовательно, за счет этой пыльцы может осуществляться только перекрестное опыление между особями в популяции, находящимися в разных стадиях цветения. По всей видимости, изменение доли тычиночных цветков в зонтиках на побегах одного порядка у особей с разной структурой синфлоресценций – результат влияния двух противоположных тенденций: с одной стороны, формирование максимально возможного числа семян и плодов и, с другой стороны, образование бóльшего числа тычиночных цветков для обеспечения ксеногамии в популяциях. У многих представителей сем. Apiaceae терминальные двойные зонтики самые крупные и многоцветковые в синфлоресценциях, в них образуется наибольшее число обоеполых цветков, которые в дальнейшем формируют значительно большее число семян и плодов в сравнении с другими двойными зонтиками особи (Knuth, 1898; Reuther, Claßen-Bockhoff, 2010). Образование бóльшего числа обоеполых цветков в двойных зонтиках на побегах предыдущего порядка ветвления у более разветвленных особей компенсируется возможностью формирования многочисленных тычиночных цветков в двойных зонтиках на побегах следующего порядка ветвления.
Список литературы
Barboza G.E., Hunziker A.T., Bernardello G., Cocucci A.A., Moscone A.E., Carrizo García C., Fuentes V., Dillon M.O., Bittrich V., Cosa M.T., Subils R., Romanutti A., Arroyo S., Anton A. 2016. Solanaceae. – In: Flowering Plants. Eudicots. 14: 295–357. https://doi.org/10.1007/978-3-319-28534-4_29
Bawa K.S., Opler P.A. 1975. Dioecism in tropical trees. – Evolution. 29 (1): 167–179. https://doi.org/10.1111/j.1558-5646.1975.tb00824.x
Bell C.R. 1971. Breeding systems and floral biology of the Umbelliferae. – In: Biology and Chemistry of the Umbelliferae. London. P. 93–108.
Braak J.P., Kho I.O. 1958. Some observations on the floral biology of the carrot (Daucus carota L.). – Euphytica. 7 (2): 131–139. https://doi.org/10.1007/BF00035726
Delph L.F., Galloway L.F., Stanton M.L. 1996. Sexual dimorphism in flower size. – Am. Naturalist. 148 (2): 299–320. https://doi.org/10.1086/285926
[Dem’yanova] Демьянова Е.И. 2000. К изучению антэкологии и полового полиморфизма у зонтичных лесостепного Зауралья. I. О протерандрии у зонтичных. – Вестн. Пермского ун-та. Биология. 2: 53–57.
Doust J.L. 1980. Floral sex ratios in andromonoecious Umbelliferae. – New Phytol. 85: 265–273. https://doi.org/10.1111/j.1469-8137.1980.tb04468.x
Endress P.K. 2020. Structural and temporal modes of heterodichogamy and similar patterns across angiosperms. – Bot. J. Linn. Soc. 193 (1): 5–18. https://doi.org/10.1093/botlinnean/boaa001
Froebe H.A., Ulrich G. 1978. Pseudanthien bei Umbelliferen. – Beitr. Biol. Pflanzen. 54: 175–206.
Godin V.N. 2017. Sexual forms and their ecological correlates of flowering plants in Siberia. – Russ. J. Ecol. 48 (5): 433–439. https://doi.org/10.1134/S1067413617050058
[Godin, Perkova] Годин В.Н., Перкова Т.В. 2017. Биология цветения и половой полиморфизм у видов семейства Apiaceae (Московская область). – Бот. журн. 102 (1): 35–47. https://doi.org/10.1134/S0006813617010033
[Godin et al.] Годин В.Н., Дозорова С.В., Архипова Т.В. 2019. Андромоноэция у Aegopodium podagraria (Apiaceae) в Московской области. – Вестн. Томского ун-та. Биология. 45: 47–68. https://doi.org/10.17223/19988591/45/3
Kellogg E.A. 2015. Reproductive Systems. – In: Flowering Plants. Monocots. 13: 93–101. https://doi.org/10.1007/978-3-319-15332-2_10
Knuth P. 1898. Handbuch der Blütenbiologie. Bd 2. T. 1. Leipzig. 705 S.
[Kuznetsova] Кузнецова Т.В. 1981. О соотношении структуры соцветий представителей семейств Araliaceae и Umbelliferae. – Биол. науки. 3: 65–73.
[Kuznetsova] Кузнецова Т.В. 1982. Морфология соцветий семейства зонтичных (Umbelliferae Juss. – Apiaceae Lindl.): Автореф. дис. … канд. биол. наук. М. 17 с.
[Kuznetzova, Timonin] Кузнецова Т.В., Тимонин А.К. 2017. Соцветие: морфология, эволюция, таксономическое значение (применение комплементарных подходов). М. 183 с.
Nuraliev M.S., Sokoloff D.D., Karpunina P.V., Oskolski A.A. 2019. Patterns of diversity of floral symmetry in angiosperms: a case study of the order Apiales. – Symmetry. 11 (4). Art. n. 473. https://doi.org/10.3390/sym11040473
[Petrova] Петрова С.Е. 2016. Зонтичные (Umbelliferae) Средней России: биоморфологический анализ. М. 280 с.
[Pimenov, Ostroumova] Пименов М.Г., Остроумова Т.А. 2012. Зонтичные (Umbelliferae) России. М. 477 с.
Plunkett G.M., Pimenov M.G., Reduron J.P., Kljukov E.V., van Wyk B.E., Ostroumova T.A., Henwood M.J., Tilney P.M., Spalik K., Watson M.F., Lee B.Y., Pu F.D., Webb C.J., Hart J.M., Mitchell A.D., Muckensturm B. 2018a. Apiaceae. – In: Flowering Plants. Eudicots. 15: 9–206. https://doi.org/10.1007/978-3-319-93605-5_2
Plunkett G.M., Wen J., Lowry P.P., Mitchell A.D., Henwood M.J., Fiaschi P. 2018b. Araliaceae. – In: Flowering Plants. Eudicots. 15: 413–446. https://doi.org/10.1007/978-3-319-93605-5_4
Quagliotti L. 1967. Effect of different temperatures on stalk development, flowering habit and sex expression in the carrot Daucus carota L. – Euphytica. 16 (1): 83–103. https://doi.org/10.1007/BF00034102
Reuther K., Claßen-Bockhoff R. 2010. Diversity behind uniformity – inflorescence architecture and flowering sequence in Apiaceae-Apioideae. – Plant Div. Evol. 128 (1–2): 181–220. https://doi.org/10.1127/1869-6155/2010/0128-0009
Schlessman M.A., Barrie F.R. 2004. Protogyny in Apiaceae, subfamily Apioideae: systematic and geographic distributions, associated traits, and evolutionary hypotheses. – S. African J. Bot. 70 (3): 475–487. https://doi.org/10.1016/S0254-6299(15)30232-5
Schlessman M.A., Plunkett G.M., Lowry P.P.II., Lloyd D.G. 2001. Sexual systems of New Caledonian Araliaceae: A preliminary phylogenetic appraisal. – Edinb. J. Bot. 58 (2): 221–228. https://doi.org/10.1017/S0960428601000592
Schlessmann M.A. 2010. Major events in the evolution of sexual systems in Apiales: ancestral andromonoecy abandoned. – Plant Div. Evol. 128 (1–2): 233–245. https://doi.org/10.1127/1869-6155/2010/0128-0011
[Shishkin] Шишкин Б.К. 1950. Род Бутень – Chaerophyllum L. – В кн.: Флора СССР. Т. 16. М., Л. С. 94–117.
Sokal R.R., Rohlf F.J. 2012. Biometry: the principles and practice of statistics in biological research. New York. 937 p.
Spalik K., Downie S.R. 2001. The utility of morphological characters for inferring phylogeny in Scandiceae subtribe Scandicinae (Apiaceae). – Ann. Missouri Bot. Gard. 88 (2): 270–301. https://doi.org/10.2307/2666227
[Vasilevich] Василевич В.И. 1998. Сероольшатники Европейской России. – Бот. журн. 83 (8): 28–42.
Webb C.J. 1981. Andromonoecism, protandry, and sexual selection in Umbelliferae. – New Zealand J. Bot. 19 (4): 335–338. https://doi.org/10.1080/0028825X.1981.10426389
Wróblewska A. 1992. Flowering and pollen value of selected species of umbelliferous family – Umbelliferae Juss. (Apiaceae Lindl.) in Lublin region. Part 1. Biology and abundance of flowering. – Acta Agrobot. 45 (1–2): 5–24. https://doi.org/10.5586/aa.1992.001
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал