

Ботанический журнал, 2021, T. 106, № 4, стр. 353-362
ИЗМЕНЧИВОСТЬ ЧИСЛА ХРОМОСОМ И ХРОМОСОМНЫЕ ПЕРЕСТРОЙКИ У PINUS SYLVESTRIS (PINACEAE) В ЗАСУШЛИВЫХ УСЛОВИЯХ НИЖНЕГО ПОВОЛЖЬЯ И ЮЖНОЙ СИБИРИ
Т. С. Седельникова 1, *, А. В. Пименов 1, **
1 Институт леса им. В.Н. Сукачева СО РАН,
Федеральный исследовательский центр “Красноярский научный центр СО РАН”
660036 Красноярск, Академгородок, 50/28, Россия
* E-mail: tss@ksc.krasn.ru
** E-mail: pimenov@ksc.krasn.ru
Поступила в редакцию 03.08.2020
После доработки 14.12.2020
Принята к публикации 22.12.2020
Аннотация
Исследованы основные критерии хромосомного полиморфизма – изменчивость числа хромосом, встречаемость и спектр хромосомных перестроек – в естественных популяциях и искусственных насаждениях сосны обыкновенной (Pinus sylvestris), произрастающих на юге ареала в степной зоне Нижнего Поволжья (Волгоградская область) и Южной Сибири (Республика Тыва, Республика Хакасия). В семенном потомстве всех исследованных деревьев выявлена миксоплоидия, уровень распространения которой возрастает по мере усиления засушливости и ухудшения эдафических условий произрастания. В семенном потомстве P. sylvestris в Волгоградской области и Хакасии обнаружены хромосомные перестройки, представленные кольцевыми хромосомами, полицентрическими хромосомами, фрагментами, множественными нарушениями. Существенное увеличение встречаемости хромосомных перестроек и расширение их спектра отмечается в проростках семян из изолированного происхождения P. sylvestris в каменистой степи Хакасии. Уровень встречаемости миксоплоидии и хромосомных перестроек у внутривидовых форм деревьев P. sylvestris неодинаков, что связано с различиями условий их произрастания. Выявленная цитологическая нестабильность семенного потомства деревьев в насаждениях P. sylvestris, произрастающих в степных экотопах, может свидетельствовать об активизации микроэволюционных процессов, что очевидно, сопровождается естественным отбором устойчивых генотипов и форм, адаптированных к засушливым лесорастительным условиям.
На южной границе своего распространения сосна обыкновенная (Pinus sylvestis L.) произрастает в виде разобщенных между собой боров, отстоящих друг от друга на большие расстояния. Изолированный, или островной, характер расположения популяций P. sylvestris на юге ареала определяется лимитирующим влиянием неблагоприятных почвенно-климатических факторов – контрастностью температур, недостатком влаги, сухостью и бедностью почвы (Pravdin, 1964). К изолированным популяциям P. sylvestris относятся боры в Воронежской и Оренбургской областях, боровые массивы в Северном Казахстане, ленточные боры в Приобье и на юге Красноярского края, островные боры в Хакасии и Туве. Степные и лесостепные автохтонные боры имеют средообразующее, а в ряде случаев – сырьевое значение, обладают значительным селекционно-генетическим потенциалом (Milyutin et al., 2013). За пределами южной границы ареала, в степных и пустынных районах Волгоградской, Астраханской областей и Калмыкии, где насаждения P. sylvestris созданы искусственно, они выполняют защитную и рекреационную функции (Bogun, Tsembelev, 2012; Manaenkov, Kulik, 2017; Semenyutina, Noyanova, 2019).
Популяции P. sylvestris на юге ареала отличаются биологическим разнообразием, выражающимся в наличии широкого спектра экотипов и форм деревьев: P. sylvestris subsp. kulundensis Sukaczew в остепненных борах азиатской части ареала, P. sylvestris var. cretaceae Kalinicz. на меловых обнажениях юга Европейской России и Украины (Pravdin, 1964). Степной экотип сосны (P. sylvestris var. stepposa Sukachev) описан для условий засушливого климата, каменистых почв и сильных ветров Казахского мелкосопочника (Petrov, 1961). Формы деревьев с нарушениями габитуса и развития – “ведьмиными метлами”, наростами, плакучей кроной, карликовые формы – встречаются в степных борах юга Сибири, Казахстана и Алтая. Количество таких деревьев возрастает по мере увеличения сухости лесорастительных условий (Shul’ga, 1979; Sosna…, 1988; Korovin et al., 2003; Tikhonova, Semerikov, 2010).
Работы, посвященные исследованию генетических факторов, контролирующих процесс формообразования у P. sylvestris на юге ареала и за его пределами, немногочисленны. В них показано, что в засушливых условиях произрастания популяции сосны обыкновенной характеризуются повышенной частотой встречаемости хромосомных мутаций, а также редких аллелей, присутствием в клетках проростков семян микроядер (Sosna…, 1988; Sedel’nikova, 2003; Butorina et al., 2007; Ti-khonova, Semerikov, 2010; Mashkina et al., 2012).
Целью настоящего исследования является анализ основных критериев хромосомного полиморфизма – изменчивости числа хромосом, спектра и уровня хромосомных перестроек в насаждениях P. sylvestris, произрастающих в засушливых условиях Нижнего Поволжья и Южной Сибири. Оценка данных критериев позволяет выявлять не только морфо-фенотипическое, но и генетическое разнообразие популяций сосны обыкновенной, диагностировать качество семенного потомства деревьев и их устойчивость.
ОБЪЕКТЫ И МЕТОДЫ
Материалами для исследования послужили естественные популяции и искусственные лесонасаждения P. sylvestris в пределах Волгоградской области, Республики Тыва и Республики Хакасия.
Волгоградская область. Сбор семенного материала проводился в 2019 г. в искусственных насаждениях P. sylvestris, созданных в 1960–70-х гг. на территории Калачевского и Суровикинского районов. К настоящему времени эти лесные массивы достигли предельного для условий степной зоны возраста – 50–60 лет. Их естественное старение и сокращение площади усугубляются недостаточностью ухода и воздействием ряда негативных факторов, связанных с нарастающей засушливостью климата и ухудшением лесопожарной обстановки. Климатические условия исследованных районов отличаются континентальностью, сухим жарким летом, дефицитом осадков, периодическими сильными юго-восточными ветрами. Почвенный покров со светло-каштановыми почвами в комплексе с солонцами имеет низкие показатели содержания гумуса, активной влаги и общей пористости (Latkina, Latkin, 2018; Semenyutina, Noyanova, 2019).
Было сформировано 4 выборки семян (каждая – с 30 деревьев), собранных в окрестностях г. Калач-на-Дону в “зеленом кольце” вокруг города (48°42'17" с.ш. 43°30'33" в.д.), х. Рюмино-Красноярский в 2 км от ландшафтного памятника природы “Голубинский песчаный массив” (48°48'03" с.ш. 43°35'43" в.д.), пос. Пятиморск (48°39'25" с.ш. 43°36'32" в.д.) и г. Суровикино (48°37'53" с.ш. 42°52'45" в.д.) вдоль федеральной автотрассы А-260. В последних двух точках семенной материал собирался с деревьев, растущих в глубине лесных массивов, в 100–200 м от проезжей части.
Исследованные насаждения представляют собой сухие боры (состав – 10С, класс бонитета – III, возраст деревьев – 40–50 лет, высота – 17–21 м, диаметр – 18–22 см). Деревья характеризуются многовершинностью, их семенная продуктивность очень низкая (выход жизнеспособных семян на одну шишку составляет 0.1–3.2 шт.).
Республика Тыва. Сбор семян проводился в 2019 г. на территории Тандинского района в естественных популяциях в Балгазынском бору в зоне Государственного природного заказника “Балгазынский” (51°02'43" с.ш. 95°07'20" в.д.). Климат района полузасушливый резко-континентальный, характеризующийся недостатком влаги, низкими зимними температурами воздуха и почв, глубоко промерзающих зимой. Балгазынский бор расположен на древних эоловых песках возвышенности Саргалык-тайга и занимает периферийные участки песчаного массива на границе с полынно-дерновинно-злаковой степью (Sosna…, 1988; Milyutin et al., 2013). Была сформирована одна общепопуляционная выборка семян (с 40 деревьев) в сосновых насаждениях, представляющих собой остепненный разнотравно-злаковый тип леса, характеризующихся разновозрастностью и низко опущенными кронами деревьев. Состав насаждений – 10С, классы бонитета – III–V. Возраст деревьев – 50–170 лет, высота – 18–22 м, диаметр – 26–44 см.
Республика Хакасия. Сбор семян проводился в 2013 г. в естественных популяциях и искусственных насаждениях P. sylvestris на территории Ширинского (“Ширинская степь”) и Бейского (“Койбальская степь”) районов. Климат исследованных районов резко континентальный, засушливый, с большими температурными контрастами, жарким летом и продолжительной малоснежной зимой. “Койбальская” и “Ширинская” степи представляют собой сухие территории с южными черноземами, местами темно-каштановыми почвами с солонцами и солончаками, в предгорьях и низкогорьях распространены каменистые почвы с неразвитым профилем (Rastitel’nyy…, 1976).
В “Ширинской степи” нами исследованы два местонахождения P. sylvestris. 1. “Карыш” – в естественной популяции на склоне горного обрамления долины р. Карыш (54°24'33" с.ш. 89°59'11'' в.д.), тип леса – сосняк низкотравно-мшистый пятнисто-мертвопокровный V класса бонитета, средние возраст, высота и диаметр деревьев соответственно – 109 лет, 15.0 м и 21.2 см. 2. “Шира” – в искусственном насаждении на прибрежных склонах вокруг озера Шира (54°30'53" с.ш. 90°08'35" в.д.), тип леса – сосняк вейниково-полынно-разнотравный IV класса бонитета, средние возраст, высота и диаметр деревьев соответственно – 52 года, 10.8 м и 21.9 см. В “Койбальской степи” изучено искусственное насаждение P. sylvestris, расположенное в зоне охраняемой территории регионального значения “Смирновский бор” (53°22'08" с.ш. 91°22'20" в.д.), тип леса – сосняк зеленомошно-разнотравно-кустарниковый мозаично-остепненный III класса бонитета, средние возраст, высота и диаметр деревьев соответственно – 68 лет, 17.6 м и 28.8 см. Популяция P. sylvestris “Карыш”, произрастающая в каменистной степи, представляет собой петрофитное редкостойное разновозрастное насаждение с преобладанием световых форм деревьев с низкой кроной. Насаждения P. sylvestris в “Ширинской” и “Койбальской” степях заложены в 50–60-е гг. XX в. (в “Ширинской степи” они сильно пострадали от пожаров). Были сформированы популяционные выборки семян внутривидовых форм (каждая – с 30–40 деревьев), выделенных по окраске микростробилов: краснопыльниковая (f. erythrantera Sanio) и желтопыльниковая (f. sulfurantera Kozubow) в происхождениях “Карыш” и “Шира” и окраске семенной кожуры: коричневосемянная (f. phaeo-sperma Kurdiani) и черносемянная (f. melanosperma Kurdiani) в происхождении “Смирновский бор”.
Семена проращивали в год их сбора в чашках Петри, проростки обрабатывали 1% р-ром колхицина в течение 4–6 часов, после промывания производили их фиксацию спиртово-уксусной смесью (3:1), хранили в холодильнике при постоянной температуре от +3 до +5°. Для исследования использовали меристематические ткани кончиков корешков проросших семян. Материал окрашивали 1% р-ром железоацетогематоксилина. Для просмотра использовали “давленые” препараты, приготовленные стандартным способом. Подсчет числа хромосом и анализ хромосомных перестроек производили на стадии метафазы митоза. Препараты просматривали под микроскопом Axiostar plus (Carl Zeiss) с применением системы формирования изображений AxioVision (окуляр ×10, объектив ×90).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Выявлено, что в диплоидном хромосомном наборе P. sylvestris, произрастающей в исследованных насаждениях, содержится 24 хромосомы (2n = 2x = 24). В семенном потомстве деревьев обнаружена миксоплоидия, при которой в клетках проростков, наряду с диплоидными, содержатся триплоидные (2n = 3x = 36) и тетраплоидные (2n = 4x = 48) клетки (рис. 1). Наиболее часто наблюдаются хромосомные расы, содержащие тетраплоидные клетки (2n = 24, 48), менее часто – триплоидные (2n = 24, 36), а также одновременно тетраплоидные и триплоидные (2n = 24, 36, 48). При этом уровень полиплоидизации диплоидных тканей проростков семян существенно варьирует (табл. 1).
Рис. 1.
Метафазные пластинки с различным числом хромосом в происхождениях P. sylvestris: a) 2n = 24 (“Балгазынский бор”), b) 2n = 24, 36 (“Балгазынский бор”), c) 2n = 24, 36 (“Пятиморск”), d) 2n = 36 (“Смирновский бор”, f. phaeosperma), e) 2n = 24, 48 (“Карыш”, f. erythranthera), f) 2n = 48 (“Калач-на-Дону”), g) 2n = 48 (“Пятиморск”), h) 2n = 48 (“Рюмино-Красноярский”), i) 2n = 48 (“Смирновский бор”, f. phaeosperma). Масштабная линейка – 10 µm.
Fig. 1. Metaphase plates with different chromosome numbers in provenances of P. sylvestris: a) 2n = 24 (“Balgazynskiy Bor”), b) 2n = 24, 36 (“Balgazynskiy Bor”), c) 2n = 24, 36 (“Pyatimorsk”), d) 2n = 36 (“Smirnovskiy Bor”, f. phaeosperma), e) 2n = = 24, 48 (“Karysh”, f. erythranthera), f) 2n = 48 (“Kalach-na-Donu”), g) 2n = 48 (“Pyatimorsk”), h) 2n = 48 (“Ryumino-Krasnoyarsk”), i) 2n = 48 (“Smirnovskiy Bor”, f. phaeosperma). Scale bar – 10 µm.
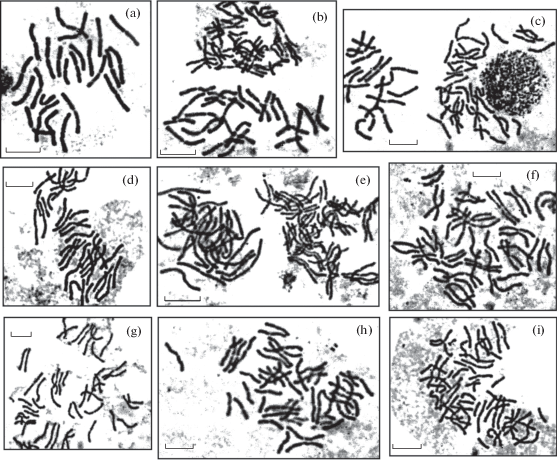
Таблица 1.
Изменчивость числа хромосом у Pinus sylvestris Table 1. The variability of the chromosome number in Pinus sylvestris
| Происхождение Provenance |
Число изученных корне-
вых меристем/клеток, шт. Number of studied root meristems/cells, pcs. |
Число корневых меристем
с измененным числом
хромосом, шт./% Number of root meristems with a changed chromo- some number, pcs./% |
Число клеток с измененным
числом хромосом, шт./% Number of cells with a changed chromosome number, pcs./% |
Хромосомный набор
корневых меристем, шт./% Chromosome set of root meristems, pcs./% |
|||
|---|---|---|---|---|---|---|---|
| 2n = 24 | 2n = 24; 36 | 2n = 24; 48 | 2n = 24; 36; 48 | ||||
| “Калач-на-Дону” “Kalach-na-Donu” |
12/263 | 4/33.3 | 9/3.4 | 8/66.7 | 1/8.3 | 3/25.0 | – |
| “Суровикино” “Surovikino” |
15/241 | 3/20.1 | 4/1.7 | 12/79.9 | 1/6.7 | 1/6.7 | 1/6.7 |
| “Пятиморск” “Pyatimorsk” |
15/474 | 11/73.7 | 29/6.1 | 4/26.7 | 3/20.0 | 5/33.3 | 3/20.0 |
| “Рюмино-Красноярский” “Ryumino-Krasnoyarskiy” |
3/165 | 3/100 | 14/8.5 | – | – | 1/33.3 | 2/66.7 |
| “Балгазынский бор” “Balgazynskiy Bor” |
15/443 | 11/73.7 | 24/5.4 | 4/26.7 | 2/13.3 | 2/13.3 | 7/46.7 |
| “Смирновский бор”, f. melanosperma “Smirnovskiy Bor”, f. melanosperma |
10/213 | 2/20.0 | 4/1.9 | 8/80.0 | – | 1/10.0 | 1/10.0 |
| “Смирновский бор”, f. phaeosperma “Smirnovskiy Bor”, f. phaeosperma |
8/289 | 4/50.0 | 6/2.1 | 4/50.0 | 1/12.5 | 2/25.0 | 1/12.5 |
| “Шира”, f. erythranthera “Shira”, f. erythranthera |
12/333 | 8/66.7 | 17/5.1 | 4/33.3 | 1/8.3 | 7/58.4 | – |
| “Шира”, f. sulfuranthera “Shira”, f. sulfuranthera |
12/341 | 7/58.3 | 23/6.7 | 5/41.7 | – | 4/33.3 | 3/25.0 |
| “Карыш”, f. erythranthera “Karysh”, f. erythranthera |
12/306 | 12/100 | 31/10.1 | – | – | 10/83.3 | 2/16.7 |
| “Карыш”, f. sulfuranthera “Karysh”, f. sulfuranthera |
12/325 | 11/91.7 | 37/11.4 | 1/8.3 | – | 9/75.0 | 2/16.7 |
Встречаемость миксоплоидных проростков в семенном потомстве деревьев различна. Минимальное их количество выявлено в популяциях “Смирновский бор” и “Суровикино”, максимальное – в насаждениях “Карыш” и “Рюмино-Красноярский” (табл. 1). Насаждения сосны в происхождениях “Карыш” и “Рюмино-Красноярский” являются территориально и репродуктивно изолированными от смежных сосновых массивов и произрастают в жестких лесорастительных условиях, первое – на сухой каменистой почве, второе – в аридных условиях окрестностей Голубинских песков, представляющих собой фрагмент пустыни в сухой степи. Тот факт, что практически все проростки в насаждениях “Карыш” и “Рюмино-Красноярский” являются миксоплоидами, может свидетельствовать о влиянии экстремальных условий произрастания деревьев на повышение гетерогенности клеточных линий, что, вероятно, способствует лучшей выживаемости семенного потомства. Для происхождений “Рюмино-Красноярский” и особенно “Карыш” характерным является разнообразие морфологических типов деревьев и наличие особей, несущих “ведьмины метлы”, что также может обусловливать высокую частоту встречаемости миксоплоидии у таких растений.
Доля миксоплоидов в проростках семян внутривидовых форм деревьев также варьирует. В локалитете “Смирновский бор” у f. phaeosperma она значительно выше, чем у f. melanosperma, а у f. erythrantera и f. sulfurantera в происхождениях “Карыш” и “Шира” имеет близкие значения. При этом сохраняется сходный баланс уровня полиплоидизации корневых меристем как между экземплярами f. phaeosperma и f. melanosperma, так и между f. erythrantera и f. sulfurantera (табл. 1). Вероятно, в засушливых степных условиях в насаждениях P. sylvestris отдельные формы деревьев являются более адаптированными и имеют преимущество, определяемое, в том числе, количеством полиплоидных клеток в соматических тканях проростков семян.
По данным предыдущих лет, доля миксоплоидов среди проростков семян из насаждений в Волгоградской области не превышала 4.8% (Sedel’nikova, 2003). В степных борах Южной Сибири встречаемость миксоплоидов в семенном потомстве деревьев составляла 3.0–25.0%, возрастая у особей с аномалиями развития и в годы с сухой и жаркой погодой (Sosna…, 1988; Mashkina et al., 2012). Сопоставление полученных нами результатов с опубликованными ранее данными показывает, что в сосновых насаждениях Волгоградской области, Тывы и Хакасии количество миксоплоидов в семенном потомстве деревьев существенно выше. Очевидно, что миксоплоидия как общий феномен характерна для южных степных насаждений сосны обыкновенной.
Одновременное присутствие клеток разного уровня плоидности в тканях обусловливается соматической редукцией хромосом, их неразъединением, элиминацией, удвоением и другими причинами. Миксоплоидия рассматривается некоторыми авторами в качестве фактора, обеспечивающего устойчивость растений, в том числе древесных, к экстремальным условиям среды. Предполагается, что сочетание клеток разной плоидности усиливает пластичность растения в целом, обеспечивает генетическое и фенотипическое разнообразие потомства. У таких растений возникают формы хромосомного полиморфизма, связанного с повышенной геномной нестабильностью. Полиплоидные и миксоплоидные формы растений чаще встречаются в неблагоприятных эдафических условиях, высокогорьях, пустынях, арктической зоне и прибрежной Антарктике (Butorina, 1989; Kunakh, 2011; Navrotska et al., 2014).
Показано, что миксоплоиды преобладают в группе устойчивых форм вяза приземистого (Ulmus pumila L.), отобранных в массово усыхающих в условиях острого дефицита влаги насаждениях в Астраханской области и Калмыкии. Значительная доля миксоплоидов выявлена среди высоко смолопродуктивных форм деревьев сосны обыкновенной, отличающихся устойчивостью к вредителям и болезням (Butorina, 1989). Явления миксоплоидии и анеуплоидии наблюдаются в популяциях большинства видов хвойных, в том числе P. sylvestris, в условиях экологического экстремума. При этом прослеживается связь степени экстремальности условий произрастания с уровнем изменчивости хромосомных чисел в семенном потомстве деревьев (Sedel’nikova, 2016).
В корневых меристемах проростков семян в большинстве исследованных происхождений P. sylvestris обнаружены структурные перестройки хромосом (табл. 2). Количество проростков и число метафазных клеток с хромосомными мутациями минимально в популяции “Смирновский бор”, максимально – в популяции “Карыш”, где высокая встречаемость и широкий спектр хромосомных мутаций в семенном потомстве деревьев, вероятно, может объясняться эффектом инбридинга вследствие высокого уровня самоопыления, свойственного изолированным популяциям. Самоопыление выявлено практически у всех деревьев в изолированных борах Южной Сибири (Sosna…, 1988). В метонахождениях “Балгазынский бор” и “Рюмино-Красноярский” хромосомных перестроек в проростках не зафиксировано, что в последнем случае может быть связано с малым количеством семян, вовлеченных в исследование вследствие их крайне низкого качества. Установлено также, что частота возникновения хромосомных перестроек в семенном потомстве внутривидовых форм деревьев неодинакова (табл. 2). Если встречаемость мутаций в проростках f. erythrantera по сравнению с f. sulfurantera (“Карыш”) характеризуется близкими значениями, то в местонахождении “Шира” – различными. В обоих случаях частота встречаемости хромосомных перестроек в корневых меристемах f. erythrantera и f. sulfurantera также существенно различается. Встречаемость мутаций в проростках f. phaeosperma и f. melanosperma в популяции “Смирновский бор” имеет сходные значения.
Таблица 2.
Встречаемость и спектр хромосомных перестроек у Pinus sylvestris Table 2. Occurrence and spectrum of chromosomal rearrangements in Pinus sylvestris
| Происхождение Provenances |
Число корневых меристем
с хромосомными перестройками шт./% Number of root meristems with chromosomal rearrangements pcs./% |
Число и спектр хромосомных перестроек
в метафазных клетках, шт./% Number and spectrum of chromosomal rearrangements in metaphase cells, pcs./% |
||||
|---|---|---|---|---|---|---|
| Всего Total |
В том числе/Including | |||||
| I | II | III | IV | |||
| “Калач-на-Дону” “Kalach-na-Donu” |
2/16.7 | 2/0.8 | 1/0.4 | – | 1/0.4 | – |
| “Суровикино” “Surovikino” |
2/13.3 | 2/0.8 | 1/0.4 | 1/0.4 | – | – |
| “Пятиморск” “Pyatimorsk” |
2/13.3 | 2/0.8 | – | – | 2/0.8 | – |
| “Рюмино-Красноярский” “Ryumino-Krasnoyarskiy” |
– | – | – | – | – | – |
| “Балгазынский бор” “Balgazynskiy Bor” |
– | – | – | – | – | – |
| “Смирновский бор”, f. melanosperma “Smirnovskiy Bor”, f. melanosperma |
1/10.0 | 1/0.5 | 1/0.5 | – | – | – |
| “Смирновский бор”, f. phaeosperma “Smirnovskiy Bor”, f. phaeosperma |
1/12.5 | 1/0.3 | 1/0.3 | – | – | – |
| “Шира”, f. erythranthera “Shira”, f. erythranthera |
2/16.7 | 2/0.6 | 2/0.6 | – | – | – |
| “Шира”, f. sulfuranthera “Shira”, f. sulfuranthera |
4/33.3 | 6/1.8 | 3/0.9 | 2/0.6 | – | 1/0.3 |
| “Карыш”, f. erythranthera “Karysh”, f. erythranthera |
9/75.0 | 27/8.8 | 18/5.9 | 1/0.3 | 4/1.3 | 4/1.3 |
| “Карыш”, f. sulfuranthera “Karysh”, f. sulfuranthera |
9/75.0 | 33/10.1 | 20/6.1 | 1/0.3 | 3/0.9 | 9/2.8 |
Примечание. Число изученных корневых меристем/клеток, шт. представлено в табл. 1. I – кольцевые структуры; II – полицентрические хромосомы; III – фрагменты; IV – множественные нарушения.
Note. The number of studied root meristems/cells, pcs. is presented in Table 1. I – ring structures; II – polycentric chromosomes; III – fragments; IV – multiple disturbances.
Необходимо отметить, что встречаемость хромосомных перестроек в проростках семян P. sylvestris, расположенных в непосредственной близости от автотрассы, не повышается по сравнению с другими изученными местонахождениями. Вероятно, выхлопные газы автотранспорта не оказывают влияния на уровень мутабильности клеток семенного потомства P. sylvestris, поскольку рассеиваются в атмосфере вследствие постоянных сильных ветров, “продувающих” сосновые насаждения в данных районах. Исследование уровня цитогенетических нарушений у древесных растений, в том числе P. sylvestris, произрастающих в условиях антропогенного загрязнения, включающего фоновые выбросы автотранспорта, показало, что его существенное увеличение происходит лишь при высоких концентрациях поллютантов (Butorina, Kalaev, 2000; Butorina et al., 2000; Goryachkina, Sizykh, 2012).
Во всех проростках семян, содержащих клетки с хромосомными перестройками, наиболее часто выявляются кольцевые хромосомы, ацентрические кольца, а также кольцевые хромосомы, “надетые” на палочковидные хромосомы (рис. 2). В проростках из местонахождений “Суровикино”, “Шира” и “Карыш” обнаружены полицентрические хромосомы, имеющие несколько центромер – дицентрические и трицентрические. Некоторые полицентрические хромосомы имеют сдвиг центромеры, диагностирующий перицентрическую инверсию. В проростках популяции “Карыш” найдена терминальная ассоциация “конец в конец” двух хромосом с перицентрической инверсией. Хроматиновые фрагменты обнаружены в проростках из насаждений “Калач-на-Дону”, “Пятиморск” и “Карыш”. В популяциях “Шира” и, особенно, “Карыш” в корневых меристемах проростков выявлены множественные хромосомные нарушения, включающие одновременно несколько типов хромосомных перестроек, которые наблюдаются в одной клетке.
Рис. 2.
Метафазные пластинки или части метафазных пластинок с хромосомными перестройками в происхождениях P. sylvestris: a) кольцевая хромосома (“Шира”, f. sulfuranthera), b-c) кольцевые хромосомы, “надетые” на палочковидные хромосомы (“Шира”, f. erythranthera – b), “Калач-на-Дону” – c)), d) ацентрическое кольцо (“Карыш”, f. erythranthera), e) кольцевая хромосома и хроматиновый фрагмент (“Карыш”, f. sulfuranthera), f) дицентрическая хромосома с перицентрической инверсией и ацентрическое кольцо (“Карыш”, f. erythranthera), g) множественные мутации одной хромосомы: хромосома с укороченным плечом, разошедшиеся хроматиды и хроматиновый фрагмент (“Пятиморск”), h) терминальная ассоциация “конец в конец” двух хромосом с перицентрической инверсией и ацентрическое кольцо (“Карыш”, f. erythranthera), i) дицентрическая хромосома и хроматиновый фрагмент (“Шира”, f. sulfuranthera). Масштабная линейка – 10 µm. Хромосомные перестройки указаны стрелками.
Fig. 2. Metaphase plates or parts of metaphase plates with chromosomal rearrangements in the provenances of P. sylvestris: a) ring chromosome (“Shira”, f. sulfuranthera), b–c) ring chromosomes “clipped” on the rod ones (“Shira”, f. erythranthera – b, “Kalach-na-Donu” – c), d) acentric ring (“Karysh”, f. erythranthera), e) ring chromosome and chromatin fragment (“Karysh”, f. sulfuranthera), f) dicentric chromosome with pericentric inversion and acentric ring (“Karysh”, f. erythranthera), g) multiple mutations of one chromosome: the chromosome with a shortened arm, diverging chromatids and chromatin fragment (“Pyatimorsk”), h) terminal association “end-to-end” of two chromosomes with pericentric inversion and acentric ring (“Karysh”, f. erythranthera), i) dicentric chromosome and chromatin fragment (“Shira”, f. sulfuranthera). Scale bar – 10 µm. Chromosomal rearrangements are indicated by arrows.
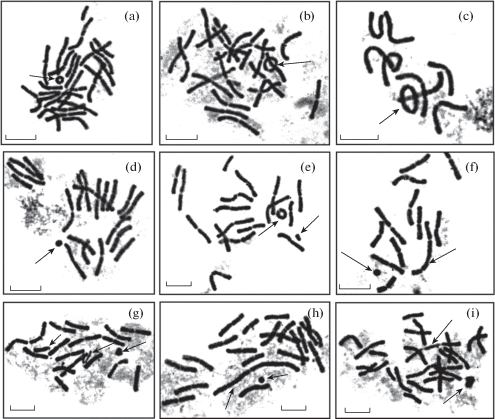
Кольцевые хромосомы, имеющие центромеру, образуются в результате делеций теломерных участков и последующего соединения плеч хромосомы в кольцо. При этом формируются дистальные ацентрические фрагменты. Полицентрические хромосомы появляются в результате транслокаций, представляющих собой обмен участками между гомологичными и негомологичными хромосомами. Перицентрическая инверсия является поворотом участка хромосомы на 180° с захватом центромеры, с точками разрыва в разных плечах. Ассоциация “конец в конец” образуется при концевом соединении хромосом вследствие стремления к конъюгации терминальных гетерохроматиновых участков (Koryakov, Zhimulyov, 2009; Kartel’ et al., 2011).
Ранее у сосны обыкновенной из засушливых местопроизрастаний Нижнего Поволжья и Южной Сибири выявлялись аналогичные хромосомные нарушения, а также другие патологии митоза. При этом доля клеток с хромосомными перестройками в проростках семян, собранных в насаждениях в Волгоградской области, составляла 6.9%, Хакасии и Тывы – 2.0–7.7%, увеличиваясь до 10.3% у карликовых и полукарликовых деревьев и в годы с длительной засухой (Sedel’nikova, 2003; Sosna…, 1988; Mashkina et al., 2012). Сравнивая полученные нами результаты с опубликованными данными, можно заключить, что в исследованных насаждениях P. sylvestris уровень мутабильности клеток и разнообразие типов хромосомных перестроек в семенном потомстве деревьев достаточно высоки. Установлено, что экстремальные факторы среды, провоцирующие геномную нестабильность, приводят к усилению мутационных процессов в популяциях хвойных растений (Sedel´nikova, 2016). Возможно, что высокий уровень и широкий спектр хромосомных перестроек в семенном потомстве деревьев в степных насаждениях P. sylvestris связаны с фактором влияния засушливых условий произрастания.
ВЫВОДЫ
1. В естественных популяциях и искусственных насаждениях сосны обыкновенной, произрастающих на юге ареала в степной зоне Нижнего Поволжья (Волгоградская область) и Южной Сибири (Республика Тыва, Республика Хакасия), выявлен хромосомный полиморфизм, выражающийся в наличии изменчивости числа хромосом – миксоплоидии, а также хромосомных перестроек в семенном потомстве деревьев.
2. Миксоплоидия (2n = 24, 36; 2n = 24, 48; 2n = = 24, 36, 48) обнаружена в семенном потомстве деревьев из всех исследованных степных популяций P. sylvestris. Установлено, что уровень распространения миксоплоидии возрастает по мере усиления засушливости и ухудшения эдафических условий произрастания деревьев. Максимальный уровень встречаемости миксоплоидии отмечается в проростках семян из изолированных местонахождений P. sylvestris – каменистой степи Хакасии и участка пустыни в сухой степи в Волгоградской области. Встречаемость миксоплоидных проростков семян варьирует у семенного потомства внутривидовых форм деревьев.
3. Хромосомные перестройки обнаружены в семенном потомстве деревьев из Волгоградской области и Хакасии. Встречаемость хромосомных перестроек различается в проростках семян внутривидовых форм деревьев. Существенное увеличение уровня встречаемости хромосомных мутаций и расширение их спектра наблюдаются в проростках семян в изолированном местообитании P. sylvestris в каменистой степи Хакасии.
4. Выявленная цитологическая нестабильность семенного потомства деревьев в насаждениях P. sylvestris, произрастающих в засушливых условиях, вероятно, свидетельствует об активизации микроэволюционных процессов, что очевидно, сопровождается естественным отбором устойчивых генотипов и форм, адаптированных к засушливым лесорастительным условиям.
Список литературы
[Bogun, Tsembelev] Богун А.П., Цембелев М.А. 2012. Пути лесоаграрного освоения аридных ландшафтов Калмыкии. – Вестник ИКИАТ. 24 (1): 25–31.
[Butorina] Буторина А.К. 1989. Факторы эволюции кариотипов древесных. – Успехи современной биологии. 108 (3): 342–357.
[Butorina, Kalaev] Буторина А.К., Калаев В.Н. 2000. Анализ чувствительности различных критериев цитогенетического мониторинга. – Экология. 3: 206–210.
[Butorina et al.] Буторина А.К., Калаев В.Н., Вострикова Т.В., Мягкова О.Е. 2000. Цитогенетические характеристики семенного потомства некоторых видов древесных растений в условиях антропогенного загрязнения г. Воронежа. – Цитология. 42 (2): 196–201.
[Butorina et al.] Буторина А.К., Черкашина О.Н., Ермолаева О.В., Чернодубов А.И., Авдеева И.А. 2007. Цитогенетический мониторинг аутохтонных лесов Усманского и Хреновского боров. – Изв. РАН. Сер. биол. 4: 508–512.
[Goryachkina, Sizykh] Горячкина О.В., Сизых О.А. 2012. Цитогенетические реакции хвойных растений в антропогенно нарушенных районах г. Красноярска и его окрестностей. – Хвойные бореальной зоны. 30 (1–2): 46–51.
[Kartel’ et al.] Картель Н.А., Макеева Е.Н., Мезенко А.М. 2011. Генетика: Энциклопедический словарь. Минск. 992 с.
[Korovin et al.] Коровин В.В., Новицкая Л.Л., Курносов Г.А. 2003. Структурные аномалии стебля древесных растений. М. 280 с.
[Koryakov, Zhimulyov] Коряков Д.Е., Жимулев И.Ф. 2009. Хромосомы. Структура и функции. Новосибирск. 258 с.
[Kunakh] Кунах В.А. 2011. Пластичность генома соматических клеток и адаптивность растений. – Молекулярная и прикладная генетика. 12: 7–14.
[Latkina, Latkin] Латкина Т.В., Латкин В.Н. 2018. Состояние лесозащитных полос в Волгоградской области. – Успехи современного естествознания. 9: 93–100.
[Manaenkov, Kulik] Манаенков А.С., Кулик А.К. 2017. Водно-минеральные особенности субстрата и засухоустойчивость деревьев сосны. – Лесохозяйственная информация. 2: 46–56. https://doi.org/10.24419/LHI.2304-3083.2017.2.05
[Mashkina et al.] Машкина О.С., Тихонова И.В., Муратова Е.Н., Мурая Л.С. 2012. Цитогенетические особенности семенного потомства карликовых сосен на юге Восточной Сибири. – Хвойные бореальной зоны. 30 (1–2): 127–135.
[Milyutin et al.] Милютин Л.И., Новикова Т.Н., Тараканов В.В., Тихонова И.В. 2013. Сосна степных и лесостепных боров Сибири. Новосибирск. 127 с.
Navrotska D.O., Twardovska M.O., Andreev I.O., Parnikoza I.Yu., Betekhtin A.A., Zahrychuk O.M., Drobyk N.M., Hasterok R., Kunakh V.A. 2014. New forms of chromosome polymorphism in Deschampsia antarctica Desv. from the Argentine Islands of the Maritime Antarctic region. – Ukrainian Antarctic J. 13: 185–191.
[Petrov] Петров С.А. 1961. Плодоношение и качество семян сосны обыкновенной в нагорных борах Северного Казахстана. – Вестник с.-х. наук. 9: 70–81.
[Pravdin] Правдин Л.Ф. 1964. Сосна обыкновенная. М. 190 с.
[Rastitel’nyi…] Растительный покров Хакасии. 1976. Новосибирск. 423 с.
[Sedel’nikova] Седельникова Т.С. 2003. Хромосомные и геномные мутации у сосны обыкновенной в Нижнем Поволжье. – Лесоведение. 6: 28–33.
Sedel’nikova T.S. 2016. Variability of genome size in conifers under extreme environmental conditions. – Biol. Bull. Rev. 6 (2): 177–188. https://doi.org/10.1134/S2079086416020079
[Semenyutina, Noyanova] Семенютина А.В., Ноянова Н.Г. 2019. Региональная специфика озеленения малых городов южной сухостепной зоны. – Известия высших учебных заведений. Лесной журнал. 6: 146–159. https://doi.org/10.17238/issn0536-1036.2019.6.146
[Shul’ga] Шульга В.В. 1979. О карликовой форме сосны и “ведьминой метле”. – Лесоведение. 3: 82–86.
[Sosna…] Сосна обыкновенная в Южной Сибири. 1988. Красноярск. 150 с.
[Tikhonova, Semerikov] Тихонова И.В., Семериков В.Л. 2010. Генетический полиморфизм карликовых сосен на юге Средней Сибири. – Экология. 5: 330–335.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал