

Ботанический журнал, 2021, T. 106, № 5, стр. 494-507
РАННИЕ ЭТАПЫ ОНТОГЕНЕЗА И РАЗМНОЖЕНИЯ ADONIS VERNALIS (RANUNCULACEAE) В УСЛОВИЯХ КРЫМА
С. В. Шевченко 1, *, Н. В. Марко 1, **
1 Никитский ботанический сад – Национальный научный центр РАН
298648 Крым, Ялта, пгт. Никита, Россия
* E-mail: shevchenko_nbs@mail.ru
** E-mail: nataly-marko@mail.ru
Поступила в редакцию 17.03.2020
После доработки 02.11.2020
Принята к публикации 10.11.2020
Аннотация
В работе представлены результаты изучения процессов опыления, оплодотворения, эндоспермо- и эмбриогенеза у редкого вида флоры Крыма Adonis vernalis L. (Ranunculaceae). Описаны также процессы семяобразования и диссеминации. Показано, что основной стратегией выживания A. vernalis в Крыму является семенное размножение и боковая вегетативная партикуляция. Для данного вида характерно наличие полового процесса и отсутствие апомиксиса. Основной тип опыления – аллогения. К причинам редкости A. vernalis можно отнести отрицательное антропогенное воздействие, критические показатели реальной семенной продуктивности (до 40%) и всхожести семян, а также наличие малого количества проростков в ценопопуляциях, что наряду с долголетием особей способствует длительному удержанию видом территории, но не обеспечивает его значительное расселение.
Одной из актуальных задач современной ботаники является разработка тактики и стратегии сохранения видов растений, которые находятся под угрозой исчезновения. Знание репродуктивной биологии позволяет установить причины образования аберрантных семязачатков и стерильности мужской генеративной сферы, дегенерации зародыша и эндосперма, нарушения процессов семяобразования и диссеминации и, в целом, причины снижения численности растений с целью последующей разработки приемов их сохранения. Особое значение имеет группа дикорастущих растений, обладающих лекарственными свойствами, а также высокой декоративностью. К таким видам с полным основанием можно отнести Adonis vernalis L. из семейства Ranunculaceae. Это ценное лекарственное и декоративное растение, имеющее 3-й природоохранный статус. В Крыму вид произрастает в степной и предгорной зонах, на яйлах (Zhivotenko, 2000; Red Book…, 2015). Природный ареал его охватывает Западную и Центральную Европу, Пиренеи, Балканы, Предкавказье, Сибирь и Крым (Adolf, 1930). Несмотря на довольно широкий природный ареал и продолжительность жизни особей, которая превышает 100 лет (Poshkurlat, 1975а), численность A. vernalis значительно сокращается (Lux, Kryukova, 1973; Shevchenko, 1986; Parubok, 2002). Размножение данного вида в ботанических садах и питомниках семенным путем также затруднено (Larin et al., 1952; Axerold, 1961; Mukovkina, 1974; Melnik, Parubok, 2000). Вследствие этого вид взят под охрану и включен в региональные Красные книги России (Red Book…, 2015; Red Book…, 2018). Эмбриологические особенности A. vernalis наряду с другими представителями семейства Ranunculaceae исследовала Е.Л. Кордюм (Kordyum, 1959, 1961, 1978), основное внимание при этом было уделено формированию элементов зародышевого мешка, однако вопросы эмбрио- и эндоспермогенеза освещены недостаточно. В работах индийских ученых представлены результаты исследования эмбриогенеза различных видов рода Adonis, но данный вид ими не изучался (Bhandari, 1962, 1966; Vijayaraghavan, 1970). Нами проведено изучение репродуктивной биологии A. vernalis с целью пополнения сведений по общей эмбриологии цветковых растений и выяснения причин снижения численности вида в пределах природного ареала (Marko, 2005; Marko, Shevchenko, 2005; Shevchenko et al., 2010). Целью данной работы было также выявление особенностей опыления, оплодотворения, эмбриогенеза, семяобразования и воспроизведения и размножения A. vernalis в условиях естественного произрастания в предгорном Крыму для разработки приемов оптимизации возобновления и размножения данного вида.
МАТЕРИАЛ И МЕТОДИКА
Adonis vernalis L. – многолетнее поликарпическое травянистое растение, высотой 10–40 см. В условиях Крыма цветет в марте – апреле, плодоносит в июне. Стебель – моноциклический, дважды-трижды разветвленный, прямостоячий или слабо отклоненный, неопушенный, густооблиственный, у ювенильных особей – простой. Листья очередные, сидячие, неопушенные, в очертании пятиугольные, ярко-зеленые, дважды или трижды-перисторазделенные на узколинейные или щетинковидные доли (Kryukova et al., 1980). В нижней части стебля имеются коричневые чешуи, в пазухах которых закладываются почки возобновления. Корневище восходящее, почти вертикальное, неразветвленное, короткое, черно-бурое (Golubev, 1962, 1967; Ziman, 1985).
A. vernalis – гемикриптофит, ксеромезофит, гелиофит, гликофит, степной энтроф (Golubev, 1996). Вид светолюбивый, теплолюбивый, не требовательный к количеству осадков, распространен в низинах и по склонам холмов, сопровождает степные и лесостепные растительные сообщества, особенно на известковых и каменистых склонах (Golubev, 1996; Kryukova et al., 1980). Основными местообитаниями являются степи с мезофильным разнотравьем и в меньшей степени типчаковые и ковыльные степи (Poshkurlat, 1970), а также остепненные луга по всему Крыму, в предгорьях и яйлах (Lux, Kryukova, 1973).
Основные наблюдения и фиксацию растительного материала проводили в ценопопуляции предгорного Крыма (окрестности с. Лозовое Симферопольского р-на). Фенологические наблюдения проводили по методике Г.Н. Зайцева (Zaitsev, 1978), для точного определения фенофаз у A. vernalis использовали методику Акош Мате (Mate, 1977). В исследованиях цветения и опыления использовали методики А.Н. Пономарева (Ponomarev, 1960), В.Н. Голубева и Ю.С. Волокитина (Golubev, Volokitin, 1986). Видовая принадлежность насекомых-опылителей была определена ведущим научным сотрудником лаборатории энтомологии и фитопатологии Никитского ботанического сада, д.б.н. А.А. Хаустовым. Изучение морфологической структуры генеративных органов цветка, а также наблюдения за ходом раскрытия пыльников и рыльца проводили на растениях в лабораторных условиях под микроскопом МБС-9. Морфологические описания составлены согласно терминологии А. А. Федорова и З.Т. Артюшенко (Fedorov, Artyushenko, 1975; Artyushenko, 1986). Анализ зрелой пыльцы проводили на временных препаратах, окрашенных ацетоорсеином, под микроскопом Jenaval 2 фирмы СarlZeiss (Germany) в десяти полях зрения. Для выяснения возможности завязывания семян у A. vernalis при самоопылении, а также проверки его способности к апомиксису проводили изоляцию 30 бутонов длиной 10–12 мм, часть бутонов кастрировали. Опыт с изоляцией и кастрацией цветков проводили в течение двух сезонов. Приготовление постоянных препаратов осуществляли по общепринятым методикам (Romeis, 1954; Pausheva, 1988). Препараты с толщиной срезов 10–12 мкм окрашивали метиловым зеленым и пиронином, а также гематоксилином по Гейденгайну с подкраской алциановым синим (Shevchenko et al., 1986; Shevchenko, Chebotar, 1992; Zhinkina, Voro-nova, 2000). Анализ постоянных препаратов проводили под микроскопом Jenaval 2 (СarlZeiss, Germany), рисунки выполняли с помощью рисовальных аппаратов РА-4 и РА-6, микрофотографии получены при использовании цифрового фотоаппарата Canon A-80.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Цветки A. vernalis крупные, 40–70 мм в диаметре, обоеполые, симметричные, актиноморфные, полные, верхушечные, одиночные, расположенные на верхушках побегов. Чашечка и венчик – опадающие. Чашечка подпестичная, имеет 5 свободных чашелистиков, расположенных горизонтально. Чашелистики обратнояйцевидные, вдвое короче венчика, с наружной стороны они зеленовато-серые, негусто покрыты короткими волосками, с внутренней стороны голые, желто-зеленые. Венчик раздельнолепестной, имеет 16–18 лепестков (Marko, 2009). Лепестки ланцетовидные или удлиненно обратнояйцевидные 20–30 мм длиной, цельные, с заостренной верхушкой и слабо выраженными зубцами по краю. Окраска лепестков золотисто-желтая, поверхность глянцевая.
Андроцей A. vernalis представлен многочисленными желтыми тычинками, короче венчика. Стенка микроспорангия формируется по типу двудольных: эндотеций и средние слои возникают из наружного париетального слоя, а в тапетум преобразуются клетки внутреннего париетального слоя. Мейоз в микроспороцитах, в основном, протекает нормально и синхронно внутри одного пыльника. Однако в некоторых пыльниках отмечены случаи нарушения хода мейоза, такие, как цитомиксис, выброс хромосом за пределы ахроматинового веретена, отставание хромосом при их расхождении в анафазе I. Зрелые пыльцевые зерна двуклеточные, сферической формы, трехпоровые, изредка – четырехпоровые.
Гинецей полимерно-апокарпный с многочисленными (до 100) сидячими плодолистиками, спирально расположенными на коническом удлиненном цветоложе. Завязи опушенные, обратнояйцевидные, желтые или светло-зеленые. В полураскрытом цветке все пыльники закрыты. Исследование показало, что пыльники одного цветка находятся на разных стадиях развития – в пыльниках периферических кругов проходит дифференцирующий митоз в микроспорах, приводящий к образованию пыльцевого зерна, а в пыльниках центральных кругов имеются лишь микроспоры. Женская генеративная сфера в это время готова к опылению и оплодотворению, что свидетельствует о наличии у изученного вида дихогамии в виде протогинии. Через сутки начинают вскрываться пыльники наружного круга тычинок. Многочисленные рыльца, скученные в центре цветка, окружены как бы венцом столь же многочисленных тычинок, примерно половина рылец выступает над пыльниками, предоставляя возможность их опыления пыльцой с других цветков.
Все цветки идентичны, в каждом из них андроцей и гинецей пространственно разделены, то есть в этот период в цветках A. vernalis наблюдается геркогамия (Marko, 2009). В течение 1–2 дней раскрываются все пыльники наружного круга (некоторые из них в пол-оборота закручиваются по спирали) и большая часть пыльников среднего круга тычинок, тычиночные нити принимают вертикальное положение (над пыльниками выступает уже 1/3 часть гинецея). Но, тем не менее, автогении препятствуют внутренние тычинки, пыльники которых в это время закрыты и смыкаются друг с другом цельным кругом, образуя вокруг завязей своеобразный щит.
В завязи каждого плодолистика закладывается по одному семязачатку. Семязачаток анатропный, с коротким фуникулусом, крассинуцеллятный, битегмальный. Внутренний интегумент обычно двух-, нередко трехслойный, наружный интегумент более мощный – трех-, четырехслойный. Нуцеллус двуслойный, в латеральной зоне – трехслойный. Со стороны фуникулуса формируется фуникулярный обтуратор. Микропиле узкое, длинное, прямое, представлено только эндостомом. В процессе развития семязачатка возможны некоторые отклонения, например, недоразвитие интегументов, отсутствие оперкулума. Согласно нашим наблюдениям, у A. vernalis могут формироваться зародышевые мешки двух типов – Poly-gonum и Allium (Marko, 2005), что было также отмечено и у Adonis chrisocyathys (Bhandari, 1962, 1966).
Основной способ переноса пыльцы у A. vernalis – энтомофилия. Опылителем, активно посещающим цветки, является Apis mellifera L., причем перенос пыльцы насекомыми осуществлялся как на цветки, принадлежащие одному растению (гейтоногения), так и на цветки разных растений того же вида (ксеногения). По мнению Е.И. Демьяновой (Demyanova, 2000), у дихогамных растений перенос пыльцы на цветки этого же растения может рассматриваться как резервный способ опыления при несостоявшемся перекрестном опылении.
Следует отметить, что для A. vernalis свойственна протогиния, и женская генеративная сфера готова к оплодотворению раньше, чем созревает пыльца. Однако, у данного вида в конце цветения цветка возможны автогения (самоопыление) и дальнейшая автогамия. Кроме того, для A. vernalis характерно наличие на одном растении довольно большого количества цветков, находящихся на разных стадиях развития, что в определенной степени может способствовать гейтоногамии. По мнению Н.П. Старшовой (Starshova, 2000), протогиния у растений носит приспособительный характер и является более эффективным механизмом, препятствующим самоопылению, чем протандрия. В результате эффективного опыления и оплодотворения у A. vernalis образуются плоды – многоорешки. Эксперименты на наличие формирования апомиктичных семян (кастрация тычинок) и изоляция цветков в естественных условиях произрастания дали отрицательный результат. В большинстве случаев завязи, а затем и формирующиеся плодики чернели и увядали. Лишь на одном растении в результате самоопыления образовалось 5 партенокарпических плодов (всего было опылено 106 плодолистиков).
Наблюдения показали, что в конце первой декады апреля (приводимые фенологические даты относятся к цветкам на побегах I порядка, для цветков на побегах второго порядка сроки сдвигаются в соответствии с ритмами их развития на 5–7 дней), вскоре после раскрывания цветка A. vernalis, в зародышевых мешках происходят процессы оплодотворения. В это время собственные пыльники в цветках еще не вскрыты, и оплодотворение осуществляется пыльцой других цветков. Иногда оплодотворение можно наблюдать перед раскрытием цветка в полураскрытых бутонах, длина которых составляет 17–18 мм, когда в них проникают насекомые и привносят пыльцу с других цветков. Для A. vernalis характерна порогамия (рис. 1). Пыльцевая трубка содержит ядро вегетативной клетки и два червеобразных спермия. Пройдя сквозь микропиле, она входит в зародышевый мешок через одну из синергид, в которую и изливает свое содержимое. При этом в синергиде происходят изменения ядра и цитоплазмы. Ядро синергиды сжимается, уменьшаясь в объеме, а впоследствии дегенерирует. В цитоплазме исчезает крупная вакуоль, цитоплазма становится гомогенной и хорошо окрашивается гематоксилином по Гейденгайну. Впоследствии один спермий направляется к яйцеклетке, другой – к центральной клетке. Слияние гамет у A. vernalis осуществляется по премитотическому типу.
Рис. 1.
Порогамия и сингамия у Adonis vernalis (pt – пыльцевая трубка, sn – сингамия, sg – синергида, еg – яйцеклетка).
Fig. 1. Porogamy and syngamy in Adonis vernalis (pt – pollen tube, sn – syngamy, sg – synergid, eg – egg-cell).

Слияние спермия с центральной клеткой (тройное слияние) происходит раньше, чем сингамия. Деление первичного ядра эндосперма также идет намного раньше деления ядра зиготы и не сопровождается цитокинезом, а образующиеся дочерние ядра расходятся к противоположным полюсам и свободно располагаются в тяже цитоплазмы, что позволяет предположить у A. vernalis нуклеарный тип эндоспермогенеза (рис. 2). На стадии 4-ядерного эндосперма в формирующихся семенах A. vernalis еще можно наблюдать антиподы.
По нашим наблюдениям, зигота приступает к делению тогда, когда эндосперм представлен множеством ядер. Е.Л. Кордюм (Kordyum, 1961) у некоторых представителей сем. Ranunculaceae, в том числе у A. vernalis, наблюдала деление зиготы при многоядерном эндосперме. Согласно данным N.N. Bhandari (1966), у однолетних видов рода Adonis деление зиготы происходит на стадии 64-ядерного эндосперма. После поперечного деления зиготы происходит поперечное деление апикальной клетки (ca) с образованием верхней (сс) и нижней (сd) клеток (рис. 3). Впоследствии поперечно делится базальная клетка (cb) с образованием нижней клетки (ci) и средней клетки (m), в результате чего формируется линейная тетрада проэмбрио (рис. 4). Стадию линейной тетрады проэмбрио (рис. 4) мы наблюдали в конце апреля, когда в цветке A. vernalis опадали чашечка и венчик.
Рис. 2.
Зародышевый мешок Adonis vernalis на стадии двуядерного эндосперма (два последовательных среза): d sg – дегенерирующая синергида, n e – ядро эндосперма, z – зигота.
Fig. 2. Embryo sac of Adonis vernalis at the stage of binuclear endosperm (two consecutive sections): d sg – degenerating synergid, n e – nucleus of the endosperm, z – zygote.

Рис. 3.
Трехклеточный проэмбрио Adonis vernalis: a – микропилярная часть зародышевого мешка; б – трехклеточный проэмбрио (обозначения в тексте).
Fig. 3. Three-celled proembryo of Adonis vernalis: a – micropylar part of the embryo sac; b – three-celled proembryo (see the text for notation).

Рис. 4.
Линейная тетрада проэмбрио (обозначения в тексте).
Fig. 4. Linear tetrad of embryo (see the text for notation).

Затем происходит первое вертикальное деление клеток проэмбрио. Обычно сначала вертикально делятся верхние (cc), а затем нижние (cd) производные апикальной клетки (рис. 4, 5). На этой стадии еще видны остатки дегенерирующих синергид (рис. 5).
Рис. 5.
Закладка вертикальных перегородок в проэмбрио Adonis vernalis: а, б – последовательные этапы (обозначения в тексте).
Fig. 5. Laying vertical partitions in proembryo of Adonis vernalis: а, b – sequential stages (see the text for notation).

После заложения первых вертикальных перегородок, в производных апикальной клетки (cc и cd), происходит второе вертикальное деление в плоскости, расположенной под прямым углом к плоскости первого вертикального деления (рис. 6). В результате этого деления в обоих ярусах производных апикальной клетки (cc и cd) образуется по 4 клетки. Таким образом, на апикальном конце проэмбрио образуются октанты: восемь клеток, которые располагаются в два яруса (этажа) – l и l' (рис. 6). Средняя клетка (m), делится поперечно, а затем продольно, продуцируя клетки n и n' (рис. 6). Поперечные деления производных базальной клетки (ci и m) приводят к образованию суспензора – s. Характер ориентации стенок и последовательность клеточных делений проэмбрио свидетельствуют о том, что эмбриогенез A. vernalis проходит по Solanad-типу (Johansen, 1950), поскольку для данного вида характерно образование линейной тетрады клеток проэмбрио и образование основных органов зародыша из производных клетки ca, а подвеска и гипофизиса – из производных клетки cb. Необходимо отметить, что на стадии октантов наблюдается переход эндосперма из ценоцитной стадии развития в клеточную. В пристенной цитоплазме между ядрами появляются клеточные перегородки (рис. 7).
Рис. 6.
Проэмбрио Adonis vernalis на стадии октантов (серия последовательных срезов).
Fig. 6. Proembyo of Adonis vernalis on the octand stage (series of consecutive sections).
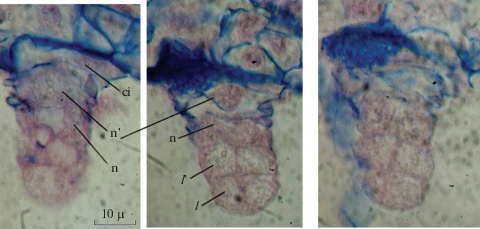
Рис. 7.
Закладка клеточных перегородок в эндосперме Adonis vernalis (pr – проэмбрио, е – эндосперм, c s – клеточная перегородка, n e – ядра эндосперма).
Fig. 7. Laying cell septa in the Adonis vernalis endosperm (pr – proembryo, e – endosperm, c s – cell septa, n e – nucleus of the endosperm).

Закладка клеточных перегородок в эндосперме происходит от периферии к центру (центростремительно) и сопровождается активными, преимущественно синхронными, делениями его ядер. В связи с этим периферические клетки эндосперма имеют правильную прямоугольную форму (рис. 8). Ко времени формирования глобулярного зародыша весь эндосперм становится клеточным (рис. 8).
Рис. 8.
Фрагменты семени Adonis vernalis на стадии глобулярного зародыша (a – микропилярная зона; б – глобулярный зародыш; в – латеральная зона эндосперма; г – центральная зона эндосперма; g e – глобулярный зародыш, е – эндосперм, sp – суспензор).
Fig. 8. Fragments of Adonis vernalis seed at the stage of the globular embryo (a – micropilar zone; б – globular embryo; в – lateral endosperm zone; г – central endosperm zone; g e – globular embryo, e – endosperm, sp – suspensor).
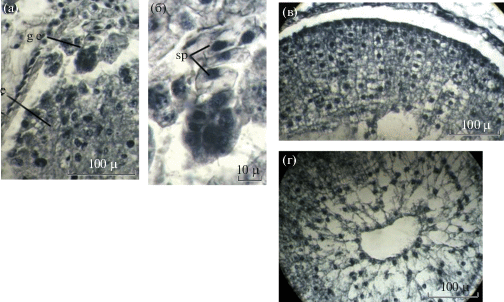
Во второй декаде мая, незадолго до диссеминации (в плоде длиной 18–20 мм), в семенах формируется глобулярный зародыш, который имеет эмбриодерму (протодерму), 3–4-клеточный суспензор и окружен слоем мощного эндосперма (рис. 8).
В целом, формирование семян A. vernalis происходит относительно быстро, в течение 24–30 дней. В сформированном семени A. vernalis, к моменту диссеминации, содержится обильный, мощный эндосперм и мелкий, недифференцированный или слабо дифференцированный на органы, зародыш, достигающий к этому времени стадии сердечка. Отношение длины зародыша к длине эндосперма в семенах A. vernalis составляет 1 : 10 (10–15%). В лабораторных условиях свежесобранные семена A. vernalis не прорастали. Анализ структуры семян, хранившихся в лаборатории, показал, что доразвитие зародыша у A. vernalis проходит во внутрисеменной период на протяжении 3 лет при температуре 18–20°С (рис. 9). Эту особенность для видов Adonis секции Consolligo также отмечала З.Е. Шлангена (Shlangena, 1979). На 3-й год доразвития зародыш A. vernalis становится отчетливо дифференцированным, при этом сформированы две семядоли, апексы побега и корня с корневым чехликом (рис. 9).
Рис. 9.
Зародыши Adonis vernalis из опавших в 2004 г., 2006 г. и в 2007 г. семян (анализ проведен в 2007 г., хранились в лабораторных условиях при t 18–20°С и относительной влажности воздуха 45%).
Fig. 9. Embryos of Adonis vernalis from seeds fallen in 2004, 2006, and 2007 (seeds were analyzed in 2007, stored under laboratory conditions at t 18–20°С and relative humidity 45%).

По литературным данным, в условиях культуры всхожесть семян A. vernalis низкая и варьирует от 1–3.3% до 16% (Larin et al., 1952; Axerold, 1961; Mukovkina, 1974; Shlangena, 1979; Mukovkina et al., 1985; Melnik, Parubok, 2004), хотя имеются данные, когда всхожесть семян достигала 40–50% (Poshkurlat, 1969а, 1975а; Florya, Kretsu, 1981), 64% (Zhivotenko, 1997), 72% (Yudin, 1997, 2002), 80–83% (Gawlowska, 1958; Nikolaeva et al., 1985). При изучении возрастной структуры ценопопуляций A. vernalis нами было отмечено довольно низкое количество проростков, в среднем составляющее от 0.3 до 4.28 экз. на 1 м2 при высоком числе генеративных особей (в пределах 10 растений), что свидетельствует о довольно низкой всхожести семян (Marko, Shevchenko, 2005).
В условиях предгорной зоны Крыма плодоношение у A. vernalis наступает в начале мая. Плод A. vernalis – многоорешек. Отдельные плодики – односемянные орешки, при созревании приобретающие сероватую или желтовато-зеленую окраску. Орешки обратнояйцевидные, мелкие (3 × 5 мм), ячеисто-морщинистые, волосистые, на верхушке с крючкообразно изогнутым носиком длиной около 1 мм. Плодики сидят на выпуклом коническом плодоложе, их количество на одном побеге A. vernalis может варьировать в пределах от 50 до 120. В среднем, один плод образует 25–30 морфологически нормальных семян. Уже в конце мая начинают осыпаться плодики с побегов I-го порядка. Вслед за ними в течение 7–10 суток они осыпаются с побегов II-го и III-го порядков (состояние репродуктивных органов определяли по Л. Д. Ефремовой (Efremova, 1971). Согласно литературным данным, A. vernalis является мирмекохором (Levina, 1967). Наши наблюдения подтверждают распространение семян муравьями у A. vernalis. Осыпание плодиков у A. vernalis происходит как раз в тот период (май–июнь), когда, согласно данным A.A. Захарова (Zakharov, 1991), муравьи выкармливают личинок и особенно активны в собирании пищи. Кроме того, A. vernalis характеризуется наличием прочных покровов плодиков и незначительных выростов тканей в базальной части околоплодника, возможно элайосом, поедаемых муравьями. На протяжении всего вегетационного периода на растениях A. vernalis (ценопопуляция в с. Лозовое) нами неоднократно наблюдались муравьи: Camponotus aethiops Lafr. и Formica cunicularia Lafr. По мнению Р. Е. Левиной (Levina, 1967), благодаря мирмекохории семена A. vernalis могут разноситься на расстояние до 10 м.
Наши исследования особенностей диссеминации A. vernalis также показали, что по способу распространения семян этот вид можно отнести и к механохорам. Зрелые плодики A. vernalis, осыпаясь с конического цветоложа, удерживаются крючочками плодов за тонко рассеченные листья и черешки, при сотрясении генеративных побегов плодики A.vernalis рассеиваются в радиусе 15–20 см от материнского растения. Крючковатые рыльца также, по-видимому, впоследствии являются приспособлением к диссеминациии, в частности к эпизоохории. Реальная семенная продуктивность ценопопуляций A. vernalis в условиях предгорного Крыма составляет 36.9–40.9%.
С целью выяснения способности вида к самовоспроизведению и самоподдержанию ценопопуляций была изучена возрастная структура ценопопуляции A. vernalis в окрестностях с. Лозовое. В результате установлено, что эта ценопопуляция нормальная, дефинитивная, неполночленная (отсутствуют особи сенильной группы) с абсолютным максимумом на средневозрастных генеративных особях. Максимум на средневозрастных генеративных особях при семенном самоподдержании, по мнению некоторых исследователей (Zaugolnova et al., 1992), связан с наибольшей продолжительностью этого состояния и с наименьшей элиминацией в этой группе особей. Возрастные спектры левостороннего типа в ценопопуляции A. vernalis динамичны по соотношению возрастных групп. Так, например, среднее количество проростков увеличивается с 0.33 на 1 м2 в 2004 г. до 4.28 на 1 м2 в 2005 г., соответственно, и ювенильных особей – с 2 до 4.52; имматурных – с 2 до 3. Число генеративных особей довольно стабильно: 9.33 на 1 м2 в 2004 г. и 9.28 на 1 м2 в 2005 г. (Marko, Shevchenko, 2005). Изменения могут быть вызваны неравномерным семенным размножением и элиминацией прегенеративных и генеративных особей. Относительно вегетативного размножения A. vernalis вопрос дискутируется. Так, А.П. Пошкурлат (Poshkurlat, 1969а, 1975a, 1975b), которая изучала возможности распространения, онтогенез и особенности строения вегетативных органов A. vernalis в течение ряда лет, утверждает, что этот вид способен размножаться только семенным путем. Другие исследователи говорят о возможности вегетативного размножения (Zhivotenko, 2000; Yudin, 2002). М.А. Цибанова (Cibanova, 1960) очень подробно описывает процессы развития, старения и распада дерновины A. vernalis, но как таковой вегетативный способ размножения не выделяет. Анализ литературных данных (Saidova, Lyubarsky, 2009) и наших исследований подземных побегов A. vernalis свидетельствует о том, что во время перехода растения в генеративное состояние наблюдается процесс ветвления корневищ, наиболее активный по периферии куста, и образование дерновины, из небольших, соединенных одно с другим корневищ (рис. 10). По мере старения куста дерновина разрушается на отдельные партикулы, способные к самостоятельному существованию живые части. По мнению А.П. Пошкурлат (Poshkurlat, 1969b, 1975b), обособившиеся стареющие партикулы не имеют омоложения и не могут служить исходным вегетативным материалом для развития новых особей, поэтому партикуляцию A. vernalis нельзя рассматривать как приспособление к вегетативному размножению. А.П. Пошкурлат (Poshkurlat, 1969b, 1975b) интерпретирует партикуляцию A. vernalis как чисто возрастное явление, не имеющее существенного биологического значения, и как один из признаков старческого распада растения. В.Н. Голубев (Golubev, 1967) считает, что биологический смысл партикуляции заключается в продолжении жизни клона и сохранении за видом определенной территории.
Рис. 10.
Дерновина Adonis vernalis (r b – почка возобновления).
Fig. 10. Turf of Adonis vernalis (r b – recovery bud).

По нашему мнению, партикуляцию у A. vernalis можно отнести к вегетативному размножению в широком смысле слова, как увеличение числа особей данного вида посредством естественного отделения от материнского растения жизнеспособных частей (без учета степени их омоложения), способных к самостоятельному существованию и развитию. Аналогичное мнение было высказано Смирновой О.В. (Smirnova, 1974) относительно ряда травянистых растений. Кроме того, биологический смысл партикуляции мы видим и в увеличении количества генеративных побегов на единицу площади, а соответственно, и семян, что приведет, согласно терминологии Ю.А. Злобина (Zlobin, 1993), к увеличению репродуктивного давления популяции на ценоз.
ЗАКЛЮЧЕНИЕ
Анализируя полученные результаты, можно заключить, что Adonis vernalis свойственно наличие полового процесса и отсутствие апомиксиса. Основной тип опыления – аллогения, перенос пыльцы осуществляется посредством энтомофилии, поэтому его успешность в значительной степени зависит от достаточного количества насекомых-опылителей в период цветения (первая-вторая декада апреля). Эндоспермо- и эмбриогенез проходят, в основном, без нарушений. Эндосперм нуклеарный, основные органы зародыша формируются из производных клетки са, подвесок и гипофиз – из производных клетки сb, что соответствует Solanad-типу эмбриогенеза. Сформированное семя содержит мощный эндосперм и мелкий, недифференцированный или слабо дифференцированный на органы зародыш, для которого характерно доразвитие во внутрисеменной период. Самоподдержание ценопопуляций A. vernalis осуществляется за счет семенного и вегетативного размножения в форме партикуляции, что в определенной степени и обусловливает его стратегию выживания. Для растений A. vernalis характерна значительная продолжительность жизни, которая и позволяет виду долгое время удерживать занимаемую территорию. Невыраженный сенильный период в анализированных ценопопуляциях свидетельствует об их относительной молодости. Причинами редкости вида A. vernalis наряду с отрицательным антропогенным воздействием (заготовка сырья для медицинской промышленности, оформление букетов в период цветения, распашка степей), вероятно, являются: критические показатели реальной семенной продуктивности (не превышает 40%); низкая всхожесть семян, наличие в семенах на стадии диссеминации недифференцированного зародыша, обусловливающего необходимость определенного периода и условий его доразвития; небольшое количество проростков в ценопопуляциях. Основными особенностями возобновления ценопопуляций A. vernalis в Крыму являются довольно слабое семенное размножение и медленное вегетативное разрастание корневища с последующей боковой партикуляцией, что способствует длительному удержанию видом территории, но не обеспечивает его значительное расселение.
Список литературы
[Adolf] Адольф Н.А. 1930. Многолетние виды рода Adonis Dill. (Подрод Consiligo ДС) в СССР. – Труды по прикладн. ботан. генетике и селекции. 23 (1): 291–356.
[Artyushenko] Артюшенко З.Т. 1986. Атлас по описательной морфологии высших растений: Плод. М. 392 с.
[Axerold] Аксерольд Д.М. 1961. Биологические особенности и приемы возделывания горицвета весеннего. – В кн.: Дикорастущие и интродуцируемые растения в Башкирии. Уфа. С. 74–86.
Bhandari N.N. 1962. Studies in the family Ranunculaceae. III. Development of the female gametophyte in Adonis annua L. – Phytomorphology. 12 (1): 70–74.
Bhandari N.N. 1966. Studies in the family Ranunculaceae. IX. Embryology of Adonis Dill ex Linn. – Phytomorphology. 16 (4): 578–587.
[Cibanova] Цибанова Н.А. 1960. К биологии горицвета (Adonis vernalis L.). – Труды Центр.-Черноземн. гос. заповедника им. В.В. Алехина. 4: 209–222.
[Demyanova] Демьянова Е.И. 2000. Аллогамия. – В кн.: Эмбриология цветковых растений: Терминология и концепции. Т. 3. СПб. С. 115–119.
[Efremova] Ефремова Л.Д. 1971. Состояние репродуктивных органов у Adonis vernalis L. на различных этапах генеративного развития. – Растит. ресурсы. 7 (2): 200–204.
[Fedorov] Федоров А.А., Артюшенко З.Т. 1975. Атлас по описательной морфологии высших растений: Цветок. Л. 352 с.
[Florya, Kretsu] Флоря В.Н., Крецу Л.Г. 1981. Биологическая характеристика горицветов Молдавии при интродукции. – Растит. ресурсы. 17 (5): 381–385.
Gawlowska J. 1958. Ochrona naturalnych zasobów a mozliwści uprawy miĺka wiosennego Adonis vernalis L. – Ochrona Przyrody. 25: 111–140.
[Golubev] Голубев В.Н. 1962. Основы биоморфологии травянистых растений центральной лесостепи. – Труды Центр. Чернозем. гос. заповедника им. В.В. Алехина: Основы биоморфологии травянистых растений центральной лесостепи. 7: 318–359.
[Golubev] Голубев В.Н. 1967. Основы биоморфологии травянистых растений Центральной лесостепи. – Журнал общ. биологии. 28 (2): 239–243.
[Golubev] Голубев В.H. 1996. Биологическая флора Крыма. 2-е изд. Ялта. 86 с.
Johansen D.A. 1950. Plant Embryology. – Chronica Botanica. Waltham MA. 305 p.
[Golubev, Volokitin] Голубев В.Н., Волокитин Ю.С. 1986. Методические рекомендации по изучению антэкологических особенностей цветковых растений: Функционально-экологические принципы организации репродуктивной структуры. Ялта. 37 с.
[Kordum] Кордюм Е.Л. 1959. Сравнительно-эмбриологическое исследование семейства Лютиковых: Автореф. дис. ... канд. биол. наук. К. 15 с.
[Kordyum] Кордюм Е.Л. 1961. Сравнительно-эмбриологическое исследование семейства Лютиковых. – В кн.: Морфогенез растений. М. Ч. 2. С. 473–477.
[Kordyum] Кордюм Е.Л. 1978. Эволюционная цитоэмбриология покрытосеменных растений. Киев. 220 с.
[Kryukova et al.] Крюкова И.В., Лукс Ю.А., Привалова Л.А. 1980. Заповедные растения Крыма: справочник. Симферополь. 96 с.
[Larin et al.] Ларин И.В., Агабабян Ш.М., Работнов Т.А., Любская А.Ф., Ларина В.К., Касименко М.А. 1952. Кормовые растения сенокосов и пастбищ СССР. Т. 2. М.–Л. 877 с.
[Levina] Левина Р.Е. 1967. Плоды. Морфология, экология, практическое значение. Саратов. 215 с.
[Lux, Kryukova] Лукс Ю.А., Крюкова И.В. 1973. Ценные, редкие и исчезающие растения флоры Крыма, подлежащие заповедной охране. – Бот журн. 58 (1): 97–106.
[Marko] Марко Н.В. 2005. Женская генеративная сфера Adonis vernalis L. (Ranunculaceae). – Бюлл. Никит. бот. сада. 91: 82–86.
[Marko] Марко Н.В. 2009. Особенности антэкология Adonis vernalis L. (Ranunculaceae). – Природничий альманах. Біологічні науки. 13: 106–113.
[Marko, Shevchenko] Марко Н.В., Шевченко С.В. 2005. О естественном возобновлении Adonis vernalis L. и Paeonia tenuifolia L. в Крыму. – Сб. науч. трудов Никит. бот. сада. 125: 88–98.
[Mate] Мате Акош 1977. Индекс цветения и индекс репродуктивности Аdonis vernalis L. – Растит. ресурсы. 13 (4): 686–692.
[Melnik, Parubok]] Мельник В.І., Парубок М.І. 2000. Проблеми охорони та раціонального використання горицвіту весняного (Adonis vernalis). – Фітотерапія в Україні. 3–4: 58–60.
[Melnik, Parubok] Мельник В.І., Парубок М.І. 2004. Горицвіт весняний (Adonis vernalis L.) в Україні. Київ. 163 с.
[Mukovkina] Муковкина З.П. 1974. К вопросу о всхожести семян горицвета весеннего. – В кн.: Биологические основы семеноведения и семеноводства интродуцентов. Новосибирск. С. 212–214.
[Mukovkina et al.] Муковкина З.П., Киричев А.Н., Карташова Л.М. 1985. Ранневесенние дикорастущие декоративные растения в Воронежском ботаническом саду. – Бюлл. Глав. бот. сада. 136: 29–32.
[Nikolaeva et al.] Николаева М.Г., Разумова М.В., Гладкова В.Н. 1985. Справочник по проращиванию покоящихся семян. Л. 347 с.
[Parubok] Парубок М.І. 2002. Горицвіт весняний (Adonis vernalis L) в Україні (еколого-ценотичні особливості та охорона): Автореф. дис. ... канд. біол. наук. Київ. 24 с.
[Pausheva] Паушева З.П. 1988. Практикум по цитологии растений. М. 271 с.
[Ponomarev] Пономарев А.Н. 1960. Изучение цветения и опыления растений. – В кн.: Полевая геоботаника. Т. 2. С. 9–19.
[Poshkurlat] Пошкурлат А.П. 1969а. Семенное размножение весеннего горицвета (Adonis vernalis L.). – Научн. докл. высш. школы. Биол. Науки. 7: 54–59.
[Poshkurlat] Пошкурлат А.П. 1969b. Развитие подземных органов виргинильных растений горицвета весеннего. – Бюл. МОИП. Отд. биол. 74 (5): 118–128.
[Poshkurlat] Пошкурлат А.П. 1970. Распространение горицвета весеннего и задача охраны его ресурсов. – Растит. ресурсы. 4 (2): 72–80.
[Poshkurlat] Пошкурлат А.П. 1975a. Урожайность семян Adonis vernalis в географическом и возрастном аспекте. – Бот. журн. 60 (4): 578–582.
[Poshkurlat] Пошкурлат А.П. 1975b. Большой жизненный цикл горицвета весеннего. – Растит. ресурсы. 11 (4): 483–492.
[Red Book…] Красная книга Республики Крым. Растения водоросли и грибы. 2015. Симферополь. 480 с.
[Red Book…] Красная книга города Севастополя. 2018. Калининград; Севастополь. 432 с.
[Romeis] Ромейс Б. 1954. Микроскопическая техника. М. 718 с.
[Saidova, Lyubarsky] Саидова Н.В., Любарский Е.Л. 2009. Диагнозы и ключи онтогенетических состояний Аdonis vernalis L. на территории республики Татарстан. – Учен. записки Казанского гос. ун-та. Естественные науки. 151 (2): 224–230.
[Shevchenko et al.] Шевченко С.В., Ругузов И.А., Ефремова. Л.М. 1986. Методика окраски постоянных препаратов метиловым зеленым и пиронином. – Бюл. Никит. бот. сада. 66: 99–101.
[Shevchenko, Chebotar] Шевченко С.В., Чеботарь А.А. 1992. Особенности эмбриологии маслины европейской (Olea europeaе). – Труды Никит. бот. сада. 113: 52–61.
[Shevchenkо et al.] Шевченко С.В., Кузьмина Т.Н., Марко Н.В., Ярославцева А.Д. 2010. Репродуктивная биология некоторых редких видов флоры Крыма. К. 392 с.
[Shevchenko] Шевченко Т.Г. 1986. Размножение горицвета весеннего в природе и в культуре. – В кн.: Материалы научно-практич. конф. “Редкие и исчезающие виды растений и животных, флористические и фаунистические комплексы Северного Кавказа, нуждающиеся в охране”. Ставрополь. С. 75–76.
[Shlangena] Шлангена З.Е. 1979. Хромосомные числа некоторых видов рода Adonis L. СССР (Ranunculaceae). – Бот журн. 61 (11): 1603–1608.
[Smirnova] Смирнова О.В. 1974. Особенности вегетативного размножения травянистых растений дубрав в связи с вопросом самоподдержания популяции. – В кн.: Возрастной состав популяций цветковых растений в связи с их онтогенезом. М. С. 168–195.
[Starshova] Старшова Н.П. 2000. Популяционные аспекты детерминации пола. – Эмбриология цветковых растений: Терминология и концепции. Т. 3. СПб. С. 88–93.
Vijayaraghavan M.R. 1970. Ranunculaceae. – Bulletin of the Indian National Science Academy. 41: 45–52.
[Yudin] Юдин С.И. 1997. Особенности семенного размножения Adonis vernalis L. в культуре. – В кн.: Тези ІV Міжнар. конф. з медичної ботаніки. Київ. С. 268–270.
[Yudin] Юдін С.І. 2002. Біологічні особливості насіннєвого розмноження Adonis vernalis L. – Інтродукція рослин. 3–4: 63–67.
[Zaitsev] Зайцев Г.Н. 1978. Фенология травянистых многолетников. М. 150 с.
[Zakharov] Захаров А.А. 1991. Организация сообществ у муравьев. М. 272 с.
[Zaugolnova et al.] Заугольнова Л.Б., Никитина С.В., Денисова Л.В. 1992. Типы функционирования популяций редких видов растений. – Бюлл. МОИП: отд. биол. 97 (3): 80–91.
[Zhinkina, Voronova] Жинкина Н.А., Воронова О.Н. 2000. К методике окраски эмбриологических препаратов. – Бот. журн. 85 (6): 168–170.
[Zhivotenko] Животенко Л.Ф. 1997. Состояние ресурсов адониса весеннего в предгорном Крыму. – В кн.: Тезисы докл. ІV Міжнародна конференція з медичної ботаніки. Киев. С. 90–91.
[Zhivotenko] Животенко Л.Ф. 2000. Распространение Adonis vernalis L. в условиях антропогенных нагрузок в предгорном Крыму. – В кн.: Тез. докл. Міжнар. конф. Проблеми сучасної екології. Запоріжжя. С. 64.
[Ziman] Зиман С.М. 1985. Морфология и филогения семейства Лютиковых. Киев. 248 с.
[Zlobin] Злобин Ю.А. 1993. Популяционное и ценотическое регулирование репродукции у цветковых растений. – Труды Бот. ин-та им. В.Л. Комарова РАН: Проблемы репродуктивной биологии семенных растений. 8: 8–15.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал