

Ботанический журнал, 2021, T. 106, № 6, стр. 540-555
ПРОЯВЛЕНИЕ ПОЛОВОГО ПОЛИМОРФИЗМА В СОЦВЕТИЯХ HERACLEUM SIBIRICUM (APIACEAE) В МОСКОВСКОЙ ОБЛАСТИ
В. Н. Годин 1, *, Т. В. Архипова 1, Ж. И. Яламова 1
1 Московский педагогический государственный университет
129164 Москва, ул. Кибальчича, 6, корп. 3, Россия
* E-mail: vn.godin@mpgu.su
Поступила в редакцию 15.01.2021
После доработки 21.01.2021
Принята к публикации 16.02.2021
Аннотация
Половой полиморфизм и структура синфлоресценций Heracleum sibiricum изучены в 2017–2019 годах в трех ценопопуляциях на территории Московской области. Выявлено четыре типа особей H. sibi-ricum по степени разветвленности соцветий (двойные зонтики на побегах I, I–II, I–III и I–IV порядков ветвления), которым соответствуют три типа синфлоресценций: единственный терминальный двойной зонтик, кисть из двойных зонтиков и метелка из двойных зонтиков. При возрастании степени разветвленности синфлоресценций происходит увеличение размеров двойных зонтиков и их структурных компонентов, а также уменьшение числа тычиночных цветков на побегах одного порядка ветвления. Простые зонтики могут включать как обоеполые, пестичные и тычиночные цветки (последние всегда занимают центральное положение), только обоеполые (у особей в терминальных двойных зонтиках с высокой степенью разветвленности синфлоресценций), обоеполые и тычиночные цветки (чаще всего в двойных зонтиках на побегах III и IV порядков), только пестичные (у редко встречающихся исключительно женских особей на побегах только I–II порядков ветвления). У H. sibiricum наблюдается сочетание андромоноэции и гинодиэции на популяционном уровне, что выражается в наличии трех типов особей: андромоноэцичных (обоеполые и тычиночные цветки; доля таких особей в популяциях варьирует от 88.9 до 92.1%), полигамомоноэцичных (обоеполые, пестичные и тычиночные цветки; доля этих особей колеблется от 6.8 до 8.4%) и женских (только пестичные цветки; частота таких особей изменяется от 0.9 до 3.4%). Обсуждаются причины и значение полового полиморфизма у цветковых растений.
Род Heracleum L. – один из крупных родов сем. Apiaceae (Downie et al., 2010). Согласно данным Plunkett et al. (2018) в роде Heracleum насчитывается от 120 до 130 видов. Изучение половой дифференциации представителей рода Heracleum имеет длительную историю. P. Knuth (1898) сообщает, что в состав зонтиков разных видов борщевиков входят обоеполые и тычиночные цветки. J. Hegi (1926) выделяет у борщевиков одно- и обоеполые цветки. И.П. Манденова (Mandenova, 1951) отметила, что цветки в зонтиках на боковых побегах некоторых видов данного рода часто бесплодны. И.Б. Сандина (Sandina, 1959) у H. sosnowskyi Manden. выделила четыре половых типа цветков: обоеполые, функционально тычиночные, функционально пестичные, бесполые (без функционирующих элементов андроцея и гинецея), которые могут встречаться в зонтиках в шести разных комбинациях. I тип – обоеполые и функционально тычиночные цветки (андромоноэция), встречается у 67.5% растений; II тип – обоеполые и функционально пестичные цветки (гиномоноэция), составляет 5% от общего числа особей; III тип – обоеполые, функционально тычиночные и бесполые цветки (андромоноэция) – 7.5%; IV тип – обоеполые, функционально пестичные и бесполые цветки (гиномоноэция) – 5%; V тип – только обоеполые цветки (гермафродиты) – 7.5% и VI тип – функционально пестичные и бесполые цветки (женские особи) – 7.5%.
Обоеполые, тычиночные и пестичные цветки в зонтиках разных кругов были отмечены у следующих видов рода Heracleum: H. sosnowskyi Manden. (Sandina, 1959), H. antasiaticum Manden., H. asperum (Hoffm.) M. Bieb., H. carpaticum Porc., H. dissectum Ledeb., H. ponticum (Lipsky) Schischk. ex Grossh. и др. (Tkachenko, 1989). При изучении большого фактического материала Г.К. Ткаченко (Tkachenko, 1989) удалось убедительно показать, что соотношение цветков разных половых типов в зонтиках на побегах разного порядка неодинаково у разных видов рода Heracleum. Терминальные зонтики многих изученных видов состояли в основном из обоеполых цветков, при этом число тычиночных и пестичных цветков было незначительным (от 1 до 30%). Зонтики на побегах второго порядка образованы, как правило, обоеполыми и тычиночными цветками, число последних колеблется в зависимости от вида от 25 до 100%. Пестичные цветки в зонтиках на побегах второго порядка либо отсутствуют, либо их число незначительно (0.3–2.3%). Если у особей развивались зонтики на побегах третьего порядка, то они состояли исключительно из тычиночных цветков.
В качестве объекта наших исследований выбран Heracleum sibiricum L. (борщевик сибирский) – двулетнее или многолетнее полурозеточное длительно вегетирующее травянистое растение со стержнекорневой системой, моно- или поликарпик (Sazyperova, 1984). H. sibiricum обладает евро-западносибирским ареалом, который охватывает Скандинавию, всю Восточную Европу, Предкавказье, Западную Сибирь, Монголию. H. sibiricum – лугово-лесной мезофит, в горах поднимается до лесного пояса.
Согласно данным G. E. Grosset (1974) у H. sibi-ricum в окрестностях Москвы по особенностям половой дифференциации цветков в терминальных двойных зонтиках особи распределены на четыре группы: 1) морфологически женские, тычинки которых полностью редуцированы (1.8% от состава изученных особей); 2) функционально пестичные с очень короткими, дугообразно согнутыми внутрь тычинками, пыльники которых не раскрываются (8.3%); 3) интерсексы – растения с менее длинными, часто дугообразно согнутыми внутрь тычинками, пыльники которых раскрываются только частично и содержат мало пыльцы (9.9%); 4) гермафродитные растения, имеющие длинные прямые тычинки и обильную пыльцу (80.0%).
Согласно данным A. Wróblewska (1992) на востоке Польши H. sibiricum образует синфлоресценции, в которых двойные зонтики располагаются на побегах I–III порядков, доля тычиночных цветков в зонтиках на побегах II порядка составляет в среднем 40.7%, на побегах III порядка – 56.7%. Терминальные двойные зонтики в анализируемых условиях состояли исключительно из обоеполых цветков.
Е.I. Dem’yanova et al. (2000) показали, что в популяциях H. sibiricum в Приуралье (Кунгурский лесной заказник) отмечаются немногочисленные (3.3%) женские особи, образующие только пестичные цветки.
Наши исследования (Godin, Ialamova, 2020) подтвердили, что в Московской области у H. sibi-ricum формируются три типа цветков – обоеполые, тычиночные и пестичные. Выделенные три половых типа цветков различаются размерными показателями, когда в отдельном простом зонтике на побеге одного порядка отмечается закономерное уменьшение размеров в следующем ряду: обоеполые – пестичные – тычиночные.
Следовательно, по данным разных авторов и нашим личным наблюдениям, половой полиморфизм этого вида варьирует от строгой андромоноэции до гинодиэции, что связано с формированием как минимум трех половых типов цветков – обоеполых, пестичных и тычиночных. Последние могут встречаться на особях H. sibiricum в разных сочетаниях. Поэтому целью данной работы было выявление полового полиморфизма на организменном и популяционном уровне у H. sibiricum в Московской области.
МАТЕРИАЛ И МЕТОДЫ
Выявление полового полиморфизма Heracleum sibiricum проводили в естественных условиях Московской области с 2017 по 2019 гг. Полученные материалы обрабатывали в камеральных условиях. Основные исследования проведены в трех ценопопуляциях. Ценопопуляция 1 описана в окр. п. Павловская Слобода (Инстринский р-н Московской области) на разнотравно-злаково-купыревом лугу (проективное покрытие H. sibiricum – 5%). Среди других видов травяного покрова наиболее часто встречались Anthriscus sylvestris (L.) Hoffm., Alopecurus pratensis L., Phleum pratense L., Vicia cracca L., Trifolium pratense L. Ценопопуляция 2 расположена в Одинцовском р-не Московской области, окр. г. Одинцово в составе разнотравно-злакового луга на центральной пойме (проективное покрытие H. sibiricum – 5%), в травяном покрове доминировали Festuca pratensis Huds., Poa pratensis L., Agrostis gigantea Roth, Geranium pratense L., Ranunculus polyanthemos L. Ценопопуляция 3 расположена в окрестностях п. Павловская Слобода (Инстринский р-н Московской области) в составе низинного сероольшатника (проективное покрытие Heracleum sibiricum – 10%), среди видов травяного покрова наиболее часто встречались Urtica dioica L., Lappa tomentosa (Mill.) Lam., Anthriscus sylvestris (L.) Hoffm., Filipendula ulmaria (L.) Maxim., Cirsium oleraceum (L.) Scop., Campanula trachelium L., Dactylis glomerata L. и Festuca gigantea (L.) Vill.
При описании особенностей строения синфлоресценции Heracleum sibiricum использована единая терминология и система обозначения для зонтиков, применяемая для представителей сем. Apiaceae: простой зонтик; двойной зонтик, состоящий из простых зонтиков; терминальный двойной зонтик, заканчивающий главный побег (Kuznetzova, 1982; Kuznetzova, Timonin, 2017).
Изучение половой дифференциации Heracleum sibiricum проводили на трансектах шириной 1 м в период массового цветения вида несколько раз за вегетационный сезон в течение трех лет наблюдений (2017–2019). Первый раз во время активного цветения цветков терминальных зонтиков отмечали характер распределения разных половых типов цветков и маркировали каждую особь, указывая ее половую дифференциацию. Через неделю уже во время цветения цветков двойных зонтиков на побегах II порядка вновь обследовали эти особи, отмечая особенности распределения в них обоеполых, тычиночных и пестичных цветков. Аналогичные наблюдения повторяли во время цветения цветков в двойных зонтиках на побегах III и IV порядков ветвления. После этого анализировали спектр и пространственное расположение трех половых типов цветков на каждой особи. Такие наблюдения проводили в каждой из трех исследованных ценопопуляций в течение трех лет. При этом в каждой отдельной ценопопуляции в один год исследования брали не менее чем по 100 особей данного вида средневозрастного онтогенетического состояния.
В 2018 г. во всех анализируемых ценопопуляциях в конце июня в фазе молочно-восковой спелости семян закладывали трансекты шириной 1 м и подсчитывали число особей H. sibiricum с разной степенью разветвленности синфлоресценций. В это время уже закончили цветение цветки в простых и двойных зонтиках на побегах самых высоких порядков, и происходит интенсивное развитие формирующихся семян и плодов. На трансекте изучено 100 особей данного вида в средневозрастном онтогенетическом состоянии. Выявлено, что двойные зонтики у H. sibiricum могут формироваться на побегах I–IV порядков, то есть существует четыре типа особей, различающиеся по степени разветвления их синфлоресценций. В каждой ценопопуляции по 10 особей с двойными зонтиками на побегах I, I–II, I–III и I–IV порядков были отобраны для камеральной обработки. В лабораторных условиях у этих особей исследованы диаметр двойного зонтика (на побегах всех порядков ветвления), число двойных зонтиков, длина лучей двойного зонтика, число простых зонтиков в составе отдельного двойного зонтика, диаметр простого зонтика, длина цветоножек, число обоеполых, пестичных и тычиночных цветков. В каждом двойном зонтике в анализ включены все простые зонтики. В дальнейшем полученные данные для особей с одним вариантом разветвленности синфлоресценций усредняли для получения обобщенной характеристики. Аналогичным образом поступали для всех четырех выявленных вариантов особей. Сравнение средних арифметических показателей всех изученных признаков зонтиков и их структурных компонентов показало, что не обнаруживаются достоверные отличия особей из разных ценопопуляций по анализируемым признакам. Поэтому все данные из трех исследованных ценопопуляций были усреднены для обобщенной характеристики синфлоресценций и их элементов у особей H. sibiricum в Московской обл. и представлены в табл. 1.
Таблица 1.
Морфометрические показатели простых и двойных зонтиков на побегах I–IV порядков у Heracleum sibiricum разной степени разветвленности Table 1. Values of Heracleum sibiricum morphological features of umbels and umbellets on shoots of different orders
| Признак/Morphological feature | Min–max | M ± m |
|---|---|---|
| Особи с зонтиками на побегах I порядка/Individuals with umbels on shoots of first order | ||
| Терминальные двойные зонтики/Terminal umbels | ||
| Диаметр двойного зонтика, см Diameter of umbel, cm |
9.0–11.5 | 10.3 ± 0.9 |
| Длина лучей двойного зонтика, см Length of umbel rays, cm |
3.5–8.6 | 5.6 ± 1.1 |
| Число двойных зонтиков, шт. Number of umbels |
1 | 1 |
| Число простых зонтиков в двойном зонтике, шт. Number of umbellets |
13–19 | 16.0 ± 2.1 |
| Диаметр простого зонтика, см Diameter of umbellet, cm |
1.6–2.6 | 2.1 ± 0.2 |
| Длина цветоножек, см Length of pedicels, cm |
0.5–1.5 | 0.9 ± 0.2 |
| Число цветков в простых зонтиках, шт. Number of flowers in umbellets |
19–33 | 26.3 ± 3.1 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellets, % |
16.1–47.6 | 32.7 ± 6.1 |
| Доля пестичных цветков в простых зонтиках, % Share of female flowers in umbellets, % |
0.0–3.1 | 1.5 ± 0.8 |
| Особи с зонтиками на побегах I–II порядков/Individuals with umbels on shoots of first and second orders | ||
| Терминальные двойные зонтики/Terminal umbels | ||
| Диаметр двойного зонтика, см Diameter of umbel, cm |
9.0–14.5 | 12.5 ± 0.6 |
| Длина лучей двойного зонтика, см Length of umbel rays, cm |
2.6–8.3 | 4.9 ± 0.4 |
| Число двойных зонтиков, шт. Number of umbels |
1 | 1 |
| Число простых зонтиков в двойном зонтике, шт. Number of umbellets |
16–23 | 19.1 ± 1.0 |
| Диаметр простого зонтика, см Diameter of umbellet, cm |
1.2–3.0 | 1.9 ± 0.1 |
| Длина цветоножек, см Length of pedicels, cm |
0.2–1.5 | 0.8 ± 0.1 |
| Число цветков в простых зонтиках, шт. Number of flowers in umbellets |
14–36 | 21.7 ± 1.5 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellets, % |
0.0–47.7 | 17.8 ± 3.0 |
| Доля пестичных цветков в простых зонтиках, % Share of female flowers in umbellets, % |
0.0–5.6 | 2.5 ± 0.5 |
| Двойные зонтики на побегах II порядка/Umbels on shoots of second order | ||
| Диаметр двойного зонтика, см Diameter of umbel, cm |
6.2–12.5 | 9.0 ± 0.6 |
| Длина лучей двойного зонтика, см Length of umbel rays, cm |
1.5–6.7 | 3.6 ± 0.3 |
| Число двойных зонтиков, шт. Number of umbels |
1–4 | 2.4 ± 0.1 |
| Число простых зонтиков в двойном зонтике, шт. Number of umbellets |
12–21 | 16.0 ± 0.8 |
| Диаметр простого зонтика, см Diameter of umbellet, cm |
0.9–2.2 | 1.4 ± 0.1 |
| Длина цветоножек, см Length of pedicels, cm |
0.3–1.1 | 0.5 ± 0.03 |
| Число цветков в простых зонтиках, шт. Number of flowers in umbellets |
12–31 | 20.6 ± 1.1 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellets, % |
33.3–81.3 | 52.3 ± 3.2 |
| Доля пестичных цветков в простых зонтиках, % Share of female flowers in umbellets, % |
0.0–3.6 | 0.7 ± 0.3 |
| Доля тычиночных цветков на особи, % Share of male flowers in individuals, % |
28.9–41.0 | 37.6 ± 2.3 |
| Особи с зонтиками на побегах I–III порядков/Individuals with umbels on shoots of first, second and third orders | ||
| Терминальные двойные зонтики/Terminal umbels | ||
| Диаметр двойного зонтика, см Diameter of umbel, cm |
18–20 | 18.8 ± 0.5 |
| Длина лучей двойного зонтика, см Length of umbel rays, cm |
3.5–10.8 | 7.0 ± 1.3 |
| Число двойных зонтиков, шт. Number of umbels |
1 | 1 |
| Число простых зонтиков в двойном зонтике, шт. Number of umbellets |
17–21 | 19.3 ± 1.0 |
| Диаметр простого зонтика, см Diameter of umbellet, cm |
2.2–4.0 | 2.9 ± 0.3 |
| Длина цветоножек, см Length of pedicels, cm |
0.5–1.8 | 1.0 ± 0.2 |
| Число цветков в простых зонтиках, шт. Number of flowers in umbellets |
17–36 | 25.5 ± 3.1 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellets, % |
0.0–18.8 | 3.7 ± 2.8 |
| Доля пестичных цветков в простых зонтиках, % Share of female flowers in umbellets, % |
0.0–6.7 | 4.9 ± 0.9 |
| Двойные зонтики на побегах II порядка/Umbels on shoots of second order | ||
| Диаметр двойного зонтика, см Diameter of umbel, cm |
14–16 | 14.8 ± 0.5 |
| Длина лучей двойного зонтика, см Length of umbel rays, cm |
2.9–8.1 | 5.7 ± 1.0 |
| Число двойных зонтиков, шт. Number of umbels |
2–5 | 3.0 ± 0.1 |
| Число простых зонтиков в двойном зонтике, шт. Number of umbellets |
16–19 | 17.7 ± 0.7 |
| Диаметр простого зонтика, см Diameter of umbellet, cm |
1.8–3.0 | 2.3 ± 0.2 |
| Длина цветоножек, см Length of pedicels, cm |
0.4–1.4 | 0.8 ± 0.1 |
| Число цветков в простых зонтиках, шт. Number of flowers in umbellets |
20–38 | 26.2 ± 2.6 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellets, % |
0.0–65.2 | 26.7 ± 9.3 |
| Доля пестичных цветков в простых зонтиках, % Share of female flowers in umbellets, % |
0.0–3.8 | 1.6 ± 0.7 |
| Двойные зонтики на побегах III порядка/Umbels on shoots of third order | ||
| Диаметр двойного зонтика, см Diameter of umbel, cm |
5.2–7.0 | 6.3 ± 0.5 |
| Длина лучей двойного зонтика, см Length of umbel rays, cm |
1.5–3.5 | 2.4 ± 0.3 |
| Число двойных зонтиков, шт. Number of umbels |
1–6 | 2.5 ± 0.2 |
| Число простых зонтиков в двойном зонтике, шт. Number of umbellets |
12–16 | 14.0 ± 1.0 |
| Диаметр простого зонтика, см Diameter of umbellet, cm |
0.7–1.1 | 0.9 ± 0.1 |
| Длина цветоножек, см Length of pedicels, cm |
0.2–0.7 | 0.4 ± 0.04 |
| Число цветков в простых зонтиках, шт. Number of flowers in umbellets |
8–21 | 15.8 ± 1.9 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellets, % |
27.8–100.0 | 55.8 ± 12.9 |
| Доля пестичных цветков в простых зонтиках, % Share of female flowers in umbellets, % |
0 | 0 |
| Доля тычиночных цветков на особи, % Share of male flowers in individuals, % |
8.1–46.8 | 25.8 ± 10.4 |
| Особи с зонтиками на побегах I–IV порядков/Individuals with umbels on shoots of first, second, third and fourth orders | ||
| Терминальные двойные зонтики/Terminal umbels | ||
| Диаметр двойного зонтика, см Diameter of umbel, cm |
20.0–21.0 | 20.5 ± 0.2 |
| Длина лучей двойного зонтика, см Length of umbel rays, cm |
4.5–11.5 | 8.0 ± 1.3 |
| Число двойных зонтиков, шт. Number of umbels |
1 | 1 |
| Число простых зонтиков в двойном зонтике, шт. Number of umbellets |
20–21 | 20.7 ± 0.3 |
| Диаметр простого зонтика, см Diameter of umbellet, cm |
2.2–4.5 | 3.3 ± 0.4 |
| Длина цветоножек, см Length of pedicels, cm |
0.7–2.0 | 1.4 ± 0.2 |
| Число цветков в простых зонтиках, шт. Number of flowers in umbellets |
19–40 | 29.3 ± 3.6 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellets, % |
0 | 0 |
| Доля пестичных цветков в простых зонтиках, % Share of female flowers in umbellets, % |
0–8.1 | 4.6 ± 0.9 |
| Двойные зонтики на побегах II порядка/Umbels on shoots of second order | ||
| Диаметр двойного зонтика, см Diameter of umbel, cm |
16–17 | 16.5 ± 0.2 |
| Длина лучей двойного зонтика, см Length of umbel rays, cm |
3.5–9.7 | 6.7 ± 1.1 |
| Число двойных зонтиков, шт. Number of umbels |
3–4 | 3.7 ± 0.3 |
| Число простых зонтиков в двойном зонтике, шт. Number of umbellets |
19–20 | 19.3 ± 0.3 |
| Диаметр простого зонтика, см Diameter of umbellet, cm |
2.0–3.5 | 2.7 ± 0.2 |
| Длина цветоножек, см Length of pedicels, cm |
0.6–1.7 | 1.0 ± 0.1 |
| Число цветков в простых зонтиках, шт. Number of flowers in umbellets |
17–35 | 25.3 ± 2.7 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellets, % |
0–4.0 | 1.5 ± 0.6 |
| Доля пестичных цветков в простых зонтиках, % Share of female flowers in umbellets, % |
0–1.0 | 0.5 ± 0.2 |
| Двойные зонтики на побегах III порядка/Umbels on shoots of third order | ||
| Диаметр двойного зонтика, см Diameter of umbel, cm |
8.3–10.2 | 9.4 ± 0.5 |
| Длина лучей двойного зонтика, см Length of umbel rays, cm |
2.4–7.2 | 4.5 ± 0.8 |
| Число двойных зонтиков, шт. Number of umbels |
4–5 | 4.3 ± 0.3 |
| Число простых зонтиков в двойном зонтике, шт. Number of umbellets |
13–16 | 14.7 ± 0.7 |
| Диаметр простого зонтика, см Diameter of umbellet, cm |
0.9–2.1 | 1.4 ± 0.2 |
| Длина цветоножек, см Length of pedicels, cm |
0.4–0.9 | 0.6 ± 0.1 |
| Число цветков в простых зонтиках, шт. Number of flowers in umbellets |
16–28 | 22.0 ± 1.7 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellets, % |
21.7–33.3 | 27.6 ± 1.8 |
| Доля пестичных цветков в простых зонтиках, % Share of female flowers in umbellets, % |
0 | 0 |
| Двойные зонтики на побегах IV порядка/Umbels on shoots of fourth order | ||
| Диаметр двойного зонтика, см Diameter of umbel, cm |
4.3–5.8 | 4.9 ± 0.4 |
| Длина лучей двойного зонтика, см Length of umbel rays, cm |
1.4–4.5 | 2.6 ± 0.5 |
| Число двойных зонтиков, шт. Number of umbels |
1–4 | 2.3 ± 0.7 |
| Число простых зонтиков в двойном зонтике, шт. Number of umbellets |
10–12 | 11.0 ± 0.5 |
| Диаметр простого зонтика, см Diameter of umbellet, cm |
0.6–1.4 | 1.0 ± 0.1 |
| Длина цветоножек, см Length of pedicels, cm |
0.4–0.9 | 0.6 ± 0.1 |
| Число цветков в простых зонтиках, шт. Number of flowers in umbellets |
13–24 | 18.2 ± 1.8 |
| Доля тычиночных цветков в простых зонтиках, % Share of male flowers in umbellets, % |
68.8–79.2 | 74.6 ± 1.7 |
| Доля пестичных цветков в простых зонтиках, % Share of female flowers in umbellets, % |
0 | 0 |
| Доля тычиночных цветков на особи, % Share of male flowers in individuals, % |
18.1–25.1 | 22.7 ± 1.3 |
Все полученные данные размерных показателей особей обработаны методами вариационной статистики (Sokal, Rohlf, 2012). Для каждого изучаемого признака определяли пределы варьирования (min–max), среднее значение (М), его ошибку (m). Сравнение средних арифметических проводили с помощью t-критерия Стьюдента. Для оценки степени отклонения фактических численностей от теоретически ожидаемых и сопоставления частот половых фенотипов в ценопопопуляциях использован критерий G (G-test). Результаты вычислений представлены в табл. 1 и 2.
Таблица 2.
Соотношение половых форм в ценопопуляциях Heracleum sibiricum в Московской области Table 2. Sex ratio in Heracleum sibiricum populations in Moscow Region
| Ценопопуляция Population |
Год Year |
Соотношение половых форм, % Sex ratio, % |
G | P | ||
|---|---|---|---|---|---|---|
| АМ | ПГМ | ГД | ||||
| CP 1 | 2017 | 90.8 | 7.3 | 1.9 | 0.497 | 0.220 |
| 2018 | 91.4 | 6.8 | 1.8 | |||
| 2019 | 89.5 | 8.4 | 2.1 | |||
| CP 2 | 2017 | 92.1 | 6.8 | 1.1 | 0.789 | 0.326 |
| 2018 | 90.8 | 8.3 | 0.9 | |||
| 2019 | 91.6 | 7.1 | 1.3 | |||
| CP 3 | 2017 | 88.9 | 8.0 | 3.1 | 0.145 | 0.070 |
| 2018 | 90.1 | 6.9 | 3.0 | |||
| 2019 | 89.4 | 7.2 | 3.4 | |||
Примечание. CP 1 – CP 3 – изученные ценопопуляции, АМ – андромоноэцичные особи, ПГМ – полигамомоноэцичные особи, ГД – гинодиэцичные особи, G – критерий сравнения частоты половых форм по годам исследования, P – достоверность различий.
Note. CP 1 – CP 3 – studied populations, AM – andromonoecious plants, ПГМ – polygamomonoecious plants, ГД – gynodioecious plants, G – test for comparing the frequency of sexual forms by investigated years, P – significance of differences.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
По степени разветвленности синфлоресценций выделяются четыре типа особей: 1) особи, у которых формируется только терминальный двойной зонтик на главном побеге; 2) особи, у которых двойные зонтики заканчивают главный побег и побеги II порядка, в этом случае синфлоресценция представляет собой закрытую кисть из двойных зонтиков; 3) особи, у которых двойные зонтики заканчивают главный и боковые побеги II и III порядков, формируя метелку из двойных зонтиков; 4) у самых разветвленных особей двойные зонтики образуются на главном побеге и боковых побегах II, III и IV порядков, синфлоресценция в данном случае представлена также метелкой из двойных зонтиков.
Как убедительно показала И.Ф. Сацыперова (Sazyperova, 1984), для всех видов рода Heracleum характерно наличие двойных зонтиков, заканчивающих главный побег, и двойных зонтиков на побегах II–III (реже IV–V) порядков.
У H. sibiricum возможно формирование двух флоральных единиц: двойной открытый зонтик, состоящий в свою очередь из открытых простых зонтиков, или двойной закрытый зонтик, включающий сближенные друг к другу двойные зонтики на побегах I и II порядка (реже II–III порядков). Лучи двойных зонтиков ребристые и опушенные. В каждом двойном зонтике все простые зонтики располагаются в нескольких кругах. Длина лучей двойного зонтика от периферии к центру всегда уменьшается. До начала цветения двойные зонтики у данного вида имеют уплощенную форму в центральной части, которая чашевидно вогнута. Такая форма сохраняется на протяжении периодов цветения и плодоношения. Особенности строения обертки двойных и простых зонтиков у данного вида сильно варьируют. У H. sibiricum встречаются особи как с отсутствием листочков обертки двойного зонтика, так и с их наличием в числе до 4–8 в зависимости от положения в синфлоресценции. Как правило, с возрастанием порядка побега число листочков обертки увеличивается у двойных зонтиков. Необходимо отметить, что листочки обертки у данного вида опадают полностью.
Простые зонтики в составе двойных зонтиков образуют несколько кругов цветков, у которых длина цветоножек всегда уменьшается от периферии к центру. Простые зонтики периферических кругов всегда крупнее центральных, и число цветков в них всегда больше, чем в последних. Все простые зонтики несут листочки оберточки, число которых в пределах каждого круга варьирует от 4 до 12. Листочки оберточки различаются по длине – наиболее длинные из них направлены наружу, наиболее короткие – к центру двойного зонтика.
В изученных ценопопуляциях крайне редко встречаются особи с самой низкой и самой высокой степенью разветвленности синфлоресценций. Чаще всего ценопопуляции представлены особями с двойными зонтиками на побегах I–II и I–III порядков ветвления (рис. 1). Рассмотрим морфологические особенности выделенных типов особей по степени разветвленности их синфлоресценций (табл. 1).
Рис. 1.
Соотношение особей с разной степенью разветвленности синфлоресценций у Heracleum sibiricum в трех ценопопуляциях.
По оси абсцисс – изученные ценопопуляции и типы особей с разной степенью разветвленности синфлоресценций, по оси ординат – доля (в %) особей с разной степенью разветвленности синфлоресценций.
CP 1 – CP 3 – изученные ценопопуляции, 1 – особи с двойными зонтиками на побегах I порядка, 2 – особи с двойными зонтиками на побегах I–II порядков ветвления, 3 – особи с двойными зонтиками на побегах I–III порядков ветвления, 4 – особи с двойными зонтиками на побегах I–IV порядков ветвления.
Fig. 1. The ratio of individuals with different degrees of synflorescence branching in three populations of Heracleum sibiricum.
X-axis – the studied populations and types of individuals with different degrees of synflorescence branching; Y-axis – the share (%) of individuals with different degrees of synflorescence branching.
CP 1 – CP 3 – studied populations, 1 – individuals with umbels on the shoots of I order, 2 – individuals with umbels on the shoots of I and II orders, 3 – individuals with double umbels on the shoots of I, II and III orders, 4 – individuals with double umbels on the shoots of I, II, III, and IV orders.
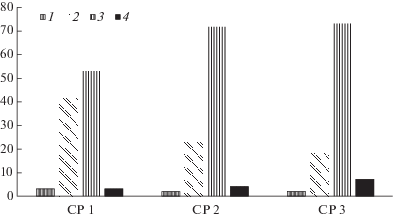
У особей с неразветвленными синфлоресценциями единственный терминальный двойной зонтик довольно крупный, до 11.5 см в диаметре (табл. 1). Доля таких особей в разных ценопопуляциях варьировала от 1.9 до 3.0% (рис. 1). У этих особей единственно возможная флоральная единица – двойной зонтик, состоящий из простых зонтиков. У особей с неразветвленными синфлоресценциями могут формироваться все три половых типа цветков – обоеполые, тычиночные и пестичные. Однако их пространственное расположение и соотношение в пределах двойного зонтика различно. Подавляющее большинство (67.3% в среднем) цветков в простых зонтиках представлено обоеполыми цветками, которые располагаются в нескольких кругах: периферических и срединных. Единичные пестичные цветки, как правило, отмечаются в периферических кругах цветков простых зонтиков. Наиболее часто формирование пестичных цветков у срединных простых зонтиков в пределах двойного зонтика. Иногда пестичные цветки совсем не встречаются в терминальных двойных зонтиках у таких особей. Следовательно, выделяется два типа особей по наличию пестичных цветков: у одних особей они не выявлены, у других они крайне немногочисленны. Тычиночные цветки всегда располагаются в центре простых зонтиков, при этом их доля постепенно увеличивается при переходе от периферических простых зонтиков к центральным в пределах двойного зонтика. Следует отметить, что доля тычиночных цветков в периферических простых зонтиках обычно составляет 16–20%, а в центральных простых зонтиках достигает почти 48%.
Хотя особи с двойными зонтиками на побегах I и II порядков встречаются в изученных условиях довольно часто, их доля в ценопопуляциях может варьировать от 18.0 до 71.7% (рис. 1). Число двойных зонтиков на боковых побегах при таком варианте строения синфлоресценций варьирует от 1 до 4. Наиболее частый вариант – наличие 2 или 3 двойных зонтиков на побегах II порядка. Довольно часто у таких особей отмечается укорочение междоузлий и сближение узлов, несущих верхние боковые побеги II порядка. В результате этого терминальный двойной зонтик оказывается окруженным несколькими двойными зонтиками, образующимися на побегах II порядка. При этом цветки всех сближенных в пространстве двойных зонтиков располагаются практически в одной плоскости. Формируется новая флоральная единица – закрытый двойной зонтик из двойных зонтиков. Следовательно, у особей с синфлоресценцией, представленной кистью из двойных зонтиков, возможно существование двух флоральных единиц: двойного зонтика из простых зонтиков и двойного зонтика из двойных зонтиков.
Терминальный двойной зонтик у таких особей отличается самыми крупными общими размерами (диаметр зонтика), а также его составных частей (длина лучей, число простых зонтиков, число цветков в простых зонтиках) (табл. 1). Как и у предыдущего типа особей с отсутствием боковых побегов, по направлению к центру терминального двойного зонтика отмечается уменьшение размеров всех структурных его элементов (длина лучей, диаметр простых зонтиков, число цветков) и одновременно увеличение доли тычиночных цветков. При этом необходимо отметить, что у наиболее мощно развитых особей, как правило, с четырьмя двойными зонтиками на побегах II порядка довольно часто в терминальном двойном зонтике тычиночные цветки вообще не встречаются даже у самых внутренних простых зонтиков (табл. 1). Проведено сравнение терминальных двойных зонтиков у особей с единственным двойным зонтиком в синфлоресценции и особей с кистью из двойных зонтиков. В терминальном двойном зонтике у особей с боковыми побегами II порядка доля тычиночных цветков ниже, а доля пестичных цветков наоборот выше, чем у особей с неразветвленным главным побегом. Необходимо отметить, что иногда встречаются особи, у которых в терминальных двойных зонтиках нет пестичных цветков, что также отмечается у вышеописанного типа особей по степени разветвления синфлоресценций. Следует упомянуть, что довольно редко встречаются особи H. sibiricum, у которых в терминальном двойном зонтике не формируются тычиночные цветки даже у самых внутренних простых зонтиков.
Двойные зонтики на побегах II порядка у таких особей отличаются уменьшением размеров всех структурных элементов по сравнению с терминальными двойными зонтиками. При этом значительно увеличивается число тычиночных цветков, доля которых у самых внутренних простых зонтиков может достигать 81.3%. С возрастанием порядка побега также уменьшается доля пестичных цветков в простых и двойных зонтиках. Иногда встречались особи с полным отсутствием пестичных цветков в двойных зонтиках на побегах II порядка, при этом в терминальных двойных зонтиках такие цветки могли присутствовать.
Исходя из всего выше сказанного, можно прийти к заключению, что у особей в случае формирования кисти из двойных зонтиков отмечается: 1) закономерное уменьшение размерных показателей двойных зонтиков и их элементов с увеличением порядка побега; 2) в двойных зонтиках в направлении к его центру происходит миниатюризация простых зонтиков и их структурных элементов. Однако одновременно доля тычиночных цветков возрастает, а доля пестичных – снижается. Крайне редко встречались особи, у которых полностью отсутствовали пестичные цветки в двойных зонтиках на побегах любого порядка. Довольно часто формируется новая флоральная единица – двойной зонтик, состоящий из двойных зонтиков.
У особей с разветвленными боковыми побегами II порядка формируется метелка из двойных зонтиков. Это один из самых распространенных вариантов особей по степени разветвления синфлоресценции в изученных нами условиях Московской обл. (рис. 1). Наиболее частый вариант у таких особей, когда формируется 3–4 двойных зонтика на побегах II порядка и 2–3 двойных зонтика на побегах III порядка. При таком варианте строения синфлоресценций также часто формируется своеобразная флоральная единица – двойной зонтик из двойных зонтиков. Следует отметить, что такая новая флоральная единица может включать только терминальный двойной зонтик и двойные зонтики на побегах II порядка. Случаи пространственного сближения двойных зонтиков на побегах II и III порядков не отмечены. Это связано с тем, что боковые побеги III порядка отличаются небольшой длиной и образуемые ими двойные зонтики никогда не располагаются в одной плоскости с двойным зонтиком на побеге II порядка.
Перечисленные выше закономерности изменения параметров как простых зонтиков в составе двойного зонтика, так и двойных зонтиков на побегах разных порядков, выявленные для особей с двойными зонтиками на побегах I–II порядков ветвления, характерны и для особей с более высокой степенью разветвленности синфлоресценций (табл. 1). Отметим лишь имеющиеся различия: 1. Доля тычиночных цветков в терминальном двойном зонтике у таких особей минимальна по сравнению с менее разветвленными особями. У большинства исследованных особей двойной зонтик, формирующийся на главном побеге, вообще может не образовывать тычиночные цветки. 2. Пестичные цветки не входят в состав двойных зонтиков на побегах III порядка, даже если они формируются в двойных зонтиках на побегах предыдущих порядков. 3. Наблюдались случаи, когда цветки всего двойного зонтика на побеге III порядка были исключительно тычиночными.
Особи с еще более разветвленными синфлоресценциями, когда двойные зонтики развиваются на побегах IV порядка, встречаются в изученных условиях крайне редко (их доля варьировала от 3 до 7% от общего числа исследованных особей) (рис. 1). Для них также характерно формирование флоральных единиц, представленных двойными зонтиками из двойных зонтиков, когда терминальный двойной зонтик окружен несколькими двойными зонтиками на побегах II порядка. Число двойных зонтиков на побегах II порядка варьирует от 3 до 4, на побегах III порядка – от 4 до 5. Число двойных зонтиков на побегах IV порядка колеблется от 1 до 4. При этом у наиболее развитых особей возможна пространственная агрегация двойных зонтиков на побегах III порядка вокруг двойного зонтика на побеге II порядка. Иными словами, возможно образование флоральных единиц не только на основе терминального двойного зонтика, но и на побегах следующих порядков ветвления.
Для особей с двойными зонтиками на побегах I–IV порядков характерны те же закономерности изменения размеров как простых, так и двойных зонтиков на побегах разного порядка, которые указаны выше для особей с менее разветвлёнными синфлоресценциями. Отметим различия:
1. Терминальный зонтик у таких особей никогда не образует тычиночных цветков, поэтому они включают только обоеполые и пестичные цветки (последние могут отсутствовать).
2. Двойные зонтики на побегах II порядка могут не содержать тычиночных цветков, а если они образуются, то их доля минимальна в сравнении с аналогичными зонтиками у менее разветвленных особей.
3. В двойных зонтиках на побегах IV порядка всегда преобладают тычиночные цветки (их доля варьирует от 68.8 до 79.2%), а обоеполые цветки немногочисленны.
4. Пестичные цветки не входят в состав двойных зонтиков на побегах IV порядка, даже если они формируются в двойных зонтиках на побегах I или II порядков.
Сравнение показателей двойных зонтиков, формирующихся на побегах разных порядков, у всех четырех типов особей по степени их разветвленности синфлоресценций позволило выявить следующее (табл. 1):
1. Общие размеры двойных зонтиков и их структурных элементов на побегах одного порядка ветвления последовательно уменьшаются с увеличением степени разветвленности синфлоресценций. Так, например, самые крупные терминальные двойные зонтики характерны для самых разветвленных особей, а наиболее мелкие – у неразветвленных особей.
2. Доля тычиночных цветков в двойных зонтиках на побегах одного порядка ветвления уменьшается с возрастанием степени разветвленности побеговой системы. Если у неразветвленных особей в терминальном зонтике всегда встречаются тычиночные цветки, и их доля в простых зонтиках варьирует от 16.1 до 47.6%, то у самых разветвленных особей в терминальном двойном зонтике никогда не встречаются цветки такого типа.
3. Изменение числа пестичных цветков у разных типов особей не носит какого-либо закономерного характера. Пестичные цветки, если они образуются, встречаются только в двойных зонтиках на побегах I и II порядков.
Отдельно необходимо остановиться на участии тычиночных цветков у разных типов особей, различающихся по особенностям строения их синфлоресценций. Так, J. Lovett-Doust (1980) при изучении некоторых представителей сем. Apiaceae показал, что наблюдается определенное соотношение обоеполых и тычиночных цветков (1 : 4) у особей андромоноэцичных видов. Такое соотношение, по мнению данного автора, обеспечивает успешное завязывание семян. Тем не менее, дальнейшие исследования других авторов показали, что соотношение обоеполых и тычиночных цветков у разных представителей этого семейства варьирует в очень широких пределах: доля тычиночных цветков колеблется от 10% у Conium ma-culatum L. до 98% у Myrrhis odorata (L.) Scop. (Reuther, Claßen-Bockhoff, 2010).
Теоретически можно ожидать, что у Heracleum sibiricum с увеличением степени разветвленности общего соцветия доля тычиночных цветков, образуемых особью, будет закономерно увеличиваться, поскольку доля тычиночных цветков значительно возрастает и становится максимальной у двойных зонтиков на побегах IV порядка. Однако, наши исследования показали, что характер изменения этого показателя не совпадает с теоретически ожидаемым (табл. 1). Оказалось, что с увеличением степени разветвленности синфлоресценций доля тычиночных цветков сначала незначительно увеличивается и достигает максимальных значений у особей с двойными зонтиками на побегах I–II порядков (кисть из двойных зонтиков), а затем начинает постепенно снижаться (у особей с метелками из двойных зонтиков). В результате у особей с самыми разветвленными синфлоресценциями участие тычиночных цветков минимальное из всех морфологических вариантов особей. Такой характер изменения доли тычиночных цветков у особей с разными синфлоресценциями объясняется следующим. С увеличением степени разветвленности синфлоресценций происходят два параллельных процесса: увеличение размеров терминальных двойных зонтиков и числа цветков в их составе и значительное сокращение числа тычиночных цветков. Это приводит к тому, что в терминальных двойных зонтиках самых разветвленных синфлоресценций такой тип цветков никогда не образуется. Несмотря на увеличение числа двойных зонтиков на боковых побегах разных порядков и одновременного возрастания участия тычиночных цветков в них, тем не менее это не оказывает существенного влияния на общую долю тычиночных цветков у особей. Таким образом, у H. sibiricum наблюдается примерно одинаковое соотношение обоеполых и тычиночных цветков (от 1 : 3 до 1 : 4) у особей, различающихся по степени разветвленности синфлоресценций.
У H. sibiricum возможно образование обоеполых, пестичных и тычиночных цветков. Данные половые типы цветков могут встречаться на особях в различных комбинациях, образуя следующие варианты (табл. 2):
1) андромоноэцичные особи, в составе которых отмечается наличие обоеполых и тычиночных цветков. Доля таких особей в разных ценопопуляциях варьирует от 88.9 до 92.1%. Все четыре выявленных типа строения синфлоресценций по степени разветвленности были обнаружены у особей с таким половым набором цветков;
2) полигамомоноэцичные особи, на которых формируются все три типа цветков. Частота встречаемости таких особей в разные годы исследования в трех исследованных ценопопуляциях колебалась от 6.8 до 8.4%. У таких особей были выявлены три типа синфлоресценций, за исключением самого разветвленного варианта, когда двойные зонтики располагаются на побегах I–IV порядков;
3) женские особи, все цветки которых представлены исключительно пестичными, распространены крайне редко – от 0.9 до 3.4% от общего числа исследованных особей в трех ценопопуляциях. Поскольку последний вариант особей обладает низкой частотой, было невозможно собрать обширный материал для структурного анализа синфлоресценций у женских особей. Все обнаруженные женские особи формировали двойные зонтики только на побегах I и II порядков. Других вариантов строения синфлоресценций с исключительно пестичными цветками нами не выявлено в изученных условиях.
Сравнение изменений частот выявленных половых форм по годам в каждой отдельной ценопопуляции (табл. 2) показывает стабильность полового спектра ценопопуляций (0.145 < G < 0.789 при 0.07 < P < 0.326). Однако, за счет более высокой доли женских особей в ценопопуляции 3 выявлены достоверные отличия по частоте половых форм между изученными ценопопуляциями (G = = 1.431, P = 0.006).
Анализ литературных сведений о половом полиморфизме Heracleum sibiricum и полученных нами данных показывает следующее. Во-первых, в Московской обл. нами выявлен более широкий спектр особей по степени разветвленности синфлоресценций и формирование четырех типов особей по этому признаку. Так, в исследованиях A. Wróblewska (1992) H. sibiricum обладал двойными зонтиками, формирующимися на побегах только I–III порядков ветвления. Во-вторых, литературные и наши данные не совпадают при анализе доли тычиночных цветков в двойных зонтиках. Согласно сведениям A. Wróblewska (1992), у H. sibiricum на востоке Польши не выявлены тычиночные цветки в терминальных двойных зонтиках, тогда как в условиях Московской области такие зонтики формировали, хотя и в небольшом числе, тычиночные цветки у особей с разветвлениями синфлоресценции до III порядка. В-третьих, наши исследования подтвердили существование чисто женских особей у H. sibiricum, доля которых в популяциях сопоставима с приведенными в литературе данными (Dem’yanova et al., 2000).
Согласно данным литературы и собственным исследованиям, представители сем. Apiaceae характеризуются широким спектром вариантов половой дифференциации: андромоноэция, полигамомоноэция, гинодиэция, андродиэция, диэция. 12 видов обладают андродиэцией (неопубликованные данные), которая наиболее часто встречается в роде Centella L. (Schubert, van Wyk, 1987). Гинодиэция отмечена у 36 видов из 18 родов (Godin, 2019, 2020). Диэция распространена гораздо реже и выявлена у четырех родов (Aciphylla J.R. Forst. et G. Forst., Arctopus L., Anisotome Hook.f., Trinia Hoffm.) (Plunkett et al., 2018). Полигамомоноэция, как половая форма, довольно редко отмечается у цветковых растений, изредка появляясь в таких семействах, как Amaranthaceae, Apiaceae, Araliaceae, Chenopodiaceae, Cucurbitaceae, Hippocastanaceae, Poaceae, Polygonaceae (Yampolsky, Yampolsky, 1922). Сведения литературы по ней крайне фрагментарны. Тем не менее, полигамомоноэция, по данным Сацыперовой (Sazyperova, 1984), характерна для всех представителей рода Heracleum. Самый распространенный вариант половой экспрессии в сем. Apiaceae – андромоноэция (Schlessmann, 2010). К сожалению, в настоящее время отсутствуют точные данные о представленности данной половой формы в семействе, но мы можем судить о частоте ее встречаемости в разных географических районах. Например, во флоре Московской обл. 72.7% видов обладают андромоноэцией (Godin, Perkova, 2017), а во флоре Троицкого лесостепного заказника (Урал) – 95% (Dem’yanova, 2011). Несмотря на наличие трех типов цветков у особей H. sibiricum, пестичные цветки встречаются достаточно редко в сравнении с тычиночными, поэтому андромоноэцичная составляющая будет преобладать в половой дифференциации данного вида.
Вопрос о причинах широкого распространения андромоноэции у представителей сем. Apiaceae интересовал многих исследователей (Knuth, 1898; Lovett-Doust, 1980; Webb, 1981; Spalik, 1991). Выявление особенностей распространения разных форм половой дифференциации во всем порядке Apiales позволило M.A. Schlessman (2010) наметить некоторые возможные причины столь большой сосредоточенности андромоноэции в этой группе. Он убедительно показал, что общий предок всех представителей порядка Apiales характеризовался диэцией, поскольку данной формой половой дифференциации обладают таксоны, входящие в состав трех базальных семейств этого порядка: Pennantiaceae, Toricelliaceae и Griseliniaceae. В дальнейшем, по мнению этого автора, эволюция привела к формированию обоеполых цветков, часть которых далее дала начало тычиночным. По предположению M.A. Schlessman (2010) и ряда других авторов (Pellmyr, 1987; Brunet, Charlesworth, 1995), превращению обоеполых цветков в тычиночные благоприятствовало две причины – специфическая структура соцветий у представителей Araliaceae и Apiaceae (разные варианты зонтиков, как простых, так и сложных, и их агрегация в синфлоресценциях) и наличие дихогамии в форме протандрии и протогинии. Согласно представлениям M.A. Schlessman (2010), в синфлоресценциях Araliaceae и Apiaceae превращение более поздно зацветающих обоеполых цветков в тычиночные необходимо для достижения большего успеха особи, поскольку существует ограничение пластических ресурсов, которое не позволяет формировать таким цветкам полноценные плоды и семена на побегах самых высоких порядков ветвления. Именно для многих видов семейств Araliaceae и Apiaceae характерна внутри- и межцветковая дихогамия и крайне своеобразный порядок цветения цветков в зонтиках на побегах разного порядка, когда все цветки особи в целом сначала находятся в тычиночной стадии цветения (в случае внутрицветковой протандрии), затем синхронно переходят в рыльцевую. Такая закономерная смена тычиночной и рыльцевой стадий цветения может наблюдаться несколько раз во время цветения синфлоресценции за один вегетационный сезон (Reuther, Claßen-Bockhoff, 2010; Enderss, 2020). При этом из-за асинхронности цветения разных особей в популяции одни из них являются донорами пыльцы в определенный момент времени, другие – реципиентами. Иными словами, андромоноэцичные особи при такой последовательности цветения цветков в синфлоресценции функционально подобны особям диэцичных видов, для которых перекрестное опыление – единственно возможный вариант переноса пыльцы. По нашему мнению, такие особенности биологии цветения способствуют успеху не только отдельной особи в популяции, но и вида в целом. Наличие в зонтиках представителей рода Heracleum пестичных цветков, хотя и немногочисленных, способствует увеличению числа семян, завязавшихся в результате перекрестного опыления. Такое возможно, во-первых, благодаря особенностям цветения цветков в зонтиках на побегах разного порядка у H. sibiricum (неопубликованные данные), когда рыльцевая стадия цветения пестичных цветков полностью заканчивается к моменту начала тычиночной стадии цветения обоеполых и тычиночных цветков в зонтиках на побегах следующего порядка. Следовательно, единственно возможный способ опыления пестичных цветков – перекрёстный. Во-вторых, пестичные цветки у H. sibiricum располагаются в основном на побегах I или II порядков ветвления, поэтому ограниченность ресурсов не будет препятствовать формированию полноценных семян и плодов из пестичных цветков.
Наличие трех половых типов цветков – необходимая основа для формирования разных их сочетаний в пределах отдельной особи у H. sibiricum. Фактически у данного вида наблюдается одновременное существование трех половых форм особей – андромоноэцичных, полигамомоноэцичных и женских. Сочетание и встречаемость нескольких половых форм в пределах одной популяции у цветковых растений отмечается довольно часто в литературе. У таких представителей сем. Apiaceae, как Carum carvi L., Daucus carota L., Heracleum sphondylium L., Seseli libanotis (L.) W.D.J. Koch и др. выявлено одновременное существование в популяциях андромоноэцичных и женских особей (Dem’yanova et al., 2000). Cucumis melo L. (Cucurbitaceae) характеризуется значительным половым полиморфизмом: наряду с моноэцичными встречаются полигамомоноэцичные, андро- и гиномоноэцичные особи (Pangalo, 1943). Большинство исследователей полового полиморфизма у цветковых растений (Lloyd, Webb, 1977; Barrett, Harder, 1996; Charlesworth, 2006; Dem’yanova, 2011) считают, что наличие нескольких половых форм особей в популяциях благоприятствует перекрестному опылению, повышающему гетерозиготность популяций, а также, по всей видимости, обеспечивает дифференциацию половых форм по экологически нишам, что уменьшает внутривидовую конкуренцию и повышает общую конкурентоспособность вида. Кроме наличия структурных (или морфологических) различий трех половых форм особей, они еще различаются и функционально во времени, что обусловлено спецификой последовательности цветения цветков в зонтиках всей синфлоресценции, когда одна и та же особь за один вегетационный сезон несколько раз выступает то как “мужская” (тычиночная стадия цветения цветков), то как “женская” (рыльцевая стадия). Несомненно, такие структурные и функциональные особенности полового полиморфизма особей H. sibiricum носят явный адаптивный характер.
ВЫВОДЫ
1. По степени разветвленности побеговой системы у Heracleum sibiricum в Московской области формируются четыре типа особей, которым соответствуют три типа синфлоресценций: единственный терминальный двойной зонтик, кисть из двойных зонтиков и метелка из двойных зонтиков.
2. У H. sibiricum флоральная единица может быть представлена открытым двойным зонтиком, состоящим из простых зонтиков (у неразветвленных синфлоресценций) или двойным зонтиком из двойных зонтиков (у разветвленных вариантов синфлоресценций).
3. С увеличением степени разветвленности синфлоресценций происходит увеличение размеров двойных зонтиков и их структурных компонентов, а также уменьшение доли в них тычиночных цветков на побегах одного порядка ветвления.
4. Возможно образование разных вариантов соотношения половых типов цветков в простых и двойных зонтиках на побегах разного порядка. Простые зонтики могут включать как обоеполые, пестичные и тычиночные цветки (последние всегда занимают центральное положение), только обоеполые (у особей в терминальных двойных зонтиках с высокой степенью разветвленности синфлоресценций), обоеполые и тычиночные цветки (чаще всего в двойных зонтиках на побегах III и IV порядков), только пестичные (у редко встречающихся исключительно женских особей).
5. У H. sibiricum наблюдается сочетание андромоноэции и гинодиэции на популяционном уровне, что выражается в наличии трех типов особей: андромоноэцичных (обоеполые и тычиночные цветки), полигамомоноэцичных (обоеполые, пестичные и тычиночные цветки) и женских (только пестичные цветки).
Список литературы
Barrett S.C.H., Harder L.D. 1996. Ecology and evolution of plant mating. – Trends Ecol. Evol. 11 (2): 73–79. https://doi.org/10.1016/0169-5347(96)81046-9
Brunet J., Charlesworth D. 1995. Floral sex allocation in sequentially blooming plants. – Evolution. 49 (1): 23–44. https://doi.org/10.2307/2410293
Charlesworth D. 2006. Evolution of plant breeding systems. – Curr. Biol. 16 (17): R726–735. https://doi.org/10.1016/j.cub.2006.07.068
[Dem’yanova] Демьянова Е.И. 2011. Спектр половых типов и форм в локальных флорах Урала (Предуралья и Зауралья). – Бот. журн. 96 (10): 1297–1315.
[Dem’yanova et al.] Демьянова Е.И., Шестакова О.М., Деткова А.Г. 2000. О половой структуре популяций гинодиэцичных зонтичных Приуралья. – Вестник Пермского ун-та. Биология. 2: 58–61.
Downie S.R., Spalik K., Katz-Downie D.S., Reduron J.-P. 2010. Major clades within Apiaceae subfamily Apioideae as inferred by phylogenetic analysis of nrDNA ITS sequences. – Plant Div. Evol. 128 (1–2): 111–136. https://doi.org/10.1127/1869-6155/2010/0128-0005
Endress P.K. 2020. Structural and temporal modes of heterodichogamy and similar patterns across angiosperms. – Bot. J. Linn. Soc. 193 (1): 5–18. https://doi.org/10.1093/botlinnean/boaa001
[Godin] Годин В.Н. 2019. Распространение гинодиэции в системе APG IV. – Бот. журн. 104 (5): 669–683. https://doi.org/10.1134/S0006813619050053
[Godin] Годин В.Н. 2020. Распространение гинодиэции у цветковых растений. – Бот. журн. 105 (3): 236–252. https://doi.org/10.31857/S0006813620030023
Godin V.N., Ialamova J.I. 2020. Sexual types of flowers morphology in Heracleum sibiricum (Apiaceae). – BIO Web of Conferences. 24: 00025. https://doi.org/10.1051/bioconf/20202400025
[Godin, Perkova] Годин В.Н., Перкова Т.В. 2017. Биология цветения и половой полиморфизм у видов семейства Apiaceae (Московская область). – Бот. журн. 102 (1): 35–47. https://doi.org/10.1134/S0006813617010033
[Grosset] Гроссет Г.Э. 1974. Модификационная изменчивость Seseli peucedanoides (Bieb.) K.-Pol. и половой полиморфизм растений этого вида и Heracleum sibiricum L. – Бюллетень МОИП. Отд. Биологии. 79 (6): 57–77.
Hegi J. 1926. Illustrierte Flora von Mittel-Europa. Munchen. Bd. 5. H. 2. S. 1415–1457.
Knuth P. 1898. Handbuch der Blütenbiologie. Bd 2. T. 1. Leipzig. 705 s.
[Kuznetsova] Кузнецова Т.В. 1982. Морфология соцветий семейства зонтичных (Umbelliferae Juss. – Apiaceae Lindl.): Автореф. дис. … канд. биол. наук. М. 17 с.
[Kuznetzova, Timonin] Кузнецова Т.В., Тимонин А.К. 2017. Соцветие: морфология, эволюция, таксономическое значение (применение комплементарных подходов). М. 183 с.
Lloyd D.G., Webb C.J. 1977. Secondary sex characters in plants. – Bot. Rev. 43 (2): 177–216. https://doi.org/10.1007/BF02860717
Lovett-Doust J. 1980. Floral sex ratio in andromonoecious Umbelliferae. – New Phytol. 85 (2): 265–273. https://doi.org/10.1111/j.1469-8137.1980.tb04468.x
[Mandenova] Манденова И.П. 1951. Род Борщевик – Heracleum L. – В кн.: Флора СССР. Т. 17. М.; Л. С. 223–259.
[Pangalo] Пангало К.И. 1943. Пол и цветение у возделываемых Cucurbitaceae. – Бот. журн. 28 (1): 10–23.
Pellmyr O. 1987. Multiple sex expressions in Cimicifuga simplex: dichogamy destabilizes hermaphroditism. – Biol. J. Linn. Soc. 31 (2): 1161–1174. https://doi.org/10.1111/j.1095-8312.1987.tb01987.x
Plunkett G.M., Pimenov M.G., Reduron J.P., Kljukov E.V., van Wyk B.E., Ostroumova T.A., Henwood M.J., Tilney P.M., Spalik K., Watson M.F., Lee B.Y., Pu F.D., Webb C.J., Hart J.M., Mitchell A.D., Muckensturm B. 2018. Apiaceae. – In: Flowering Plants. Eudicots. 15: 9–206. https://doi.org/10.1007/978-3-319-93605-5_2
Reuther K., Claßen-Bockhoff R. 2010. Diversity behind uniformity – inflorescence architecture and flowering sequence in Apiaceae-Apioideae. – Plant Div. Evol. 128 (8): 181–220. https://doi.org/10.1127/1869-6155/2010/0128-0009
[Sandina] Сандина И.Б. 1959. Борщевик Сосновского (Heracleum sosnowskyi Manden.), его биология и опыт выращивания в Ленинградской области: Автореф. дис. … канд. биол. наук. Л. 18 с.
[Sazyperova] Сацыперова И.Ф. 1984. Борщевики флоры СССР – новые кормовые растения. Л. 223 с.
Schlessmann M.A. 2010. Major events in the evolution of sexual systems in Apiales: ancestral andromonoecy abandoned. – Plant Div. Evol. 128 (1–2): 233–245. https://doi.org/10.1127/1869-6155/2010/0128-0011
chubert M.T.R., van Wyk B.E. 1997. A revision of Centella series Capenses (Apiaceae). – Nord. J. Bot. 17 (3): 301–314. 10.1111/j.1756-1051.1997.tb00318.
Sokal R.R., Rohlf F.J. 2012. Biometry: the principles and practice of statistics in biological research. New York. 937 p.
Spalik K. 1991. On evolution of andromonoecy and “overproduction” of flowers: A resource allocation model. – Biol. J. Linn. Soc. 42 (3): 325–336. https://doi.org/10.1111/j.1095-8312.1991.tb00566.x
[Tkachenko] Ткаченко К.Г. 1989. Особенности цветения и семенная продуктивность некоторых видов Heracleum L., выращенных в Ленинградской области. – Раст. ресурсы. 25 (1): 52–61.
Webb C.J. 1981. Andromonoecism, protandry, and sexual selection in Umbelliferae. – New Zealand J. Bot. 19 (4): 335–338. https://doi.org/10.1080/0028825X.1981.10426389
Wróblewska A. 1992. Flowering and pollen value of selected species of umbelliferous family – Umbelliferae Juss. (Apiaceae Lindl.) in Lublin region. Part 1. Biology and abundance of flowering. – Acta agrobotanica. 45 (1–2): 5–24. https://doi.org/10.5586/aa.1992.001
Yampolsky C., Yampolsky H. 1922. Distribution of sex forms in the phanerogamic flora. – Bibliotheca Genetica. 3: 1–62.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал