

Ботанический журнал, 2022, T. 107, № 11, стр. 1027-1038
ФОРМИРОВАНИЕ ТРАВЯНИСТОЙ ПОДУШКИ И СТРУКТУРА ЦЕНОПОПУЛЯЦИИ NEPETA LIPSKYI (LAMIACEAE) НА ТУРКЕСТАНСКОМ ХРЕБТЕ
А. Ю. Асташенков 1, *, В. А. Черемушкина 1
1 Центральный сибирский ботанический сад СО РАН
630090 Новосибирск, ул. Золотодолинская, 101, Россия
* E-mail: astal@bk.ru
Поступила в редакцию 19.06.22
После доработки 23.08.2022
Принята к публикации 20.09.2022
- EDN: LHQJMX
- DOI: 10.31857/S0006813622110023
Аннотация
Изучен механизм формирования травянистой подушки в онтогенезе и дана оценка состояния ценопопуляции Nepeta lipskyi Kudrjasch. в высокогорных условиях Туркестанского хребта (Таджикистан). Установлено, что онтогенез особей полный, сложный, растения развиваются по варианту симподиальной верхнерозеточной модели побегообразования. За счет выровненной длины резидов и одинаковой высоты плотно прижатых к субстрату интенсивно ветвящихся верхнерозеточных побегов, а также ярусного ветвления близкорасположенных осей с незначительным годичным приростом формируется приземистая плоская травянистая подушка. В побегообразовании участвуют как почки регулярного возобновления, так и спящие почки. Начиная с имматурного состояния формируется каудекс, в результате ветвления особей и сохранения резидов он становится разветвленным. В течение всего онтогенеза его базальная часть разрастается, апикальная часть надстраивается резидами побегов, возникших из почек регулярного возобновления. Тонкие ветви каудекса (каудикулы) определяют подземную многолетнюю структуру особей. С помощью архитектурного анализа установлено, что многолетние оси (каудикулы) строятся в результате линейного последовательного симподиального сочленения резидов вегетативных модулей. Первый модуль любой оси формируется из спящей почки, в дальнейшем ось строится за счет развития модулей из почек регулярного возобновления. Последний годичный прирост всегда генеративный, после чего ось разрушается. Онтогенетический спектр изученной ценопопуляции центрированный, полночленный, совпадает с характерным спектром. Индексы возрастности и эффективности (Δ = 0.47; ω = 0.69) указывают на ценопопуляцию как переходную к зрелой. Онтогенетический состав и демографические показатели отражают устойчивое нормальное состояние ценопопуляции.
Подушковидные растения занимают особое место в системе жизненных форм высших растений (Raunkiær, 1934; Serebryakov, 1962). Они широко распространены среди цветковых растений, отличаются обширным ареалом, приурочены к горным областям и высоким поясам от Арктики до субантарктических островов (Hauri, Schröter, 1914; Körner, 2003; Butterfield et al., 2013; Aubert et al., 2014). Все местообитания подушек характеризуются крайними неблагоприятными условиями для роста надземных осей (Rauh, 1939; Stanyukovich, 1949; Steshenko, 1965; Udalova, 1978). В различных местообитаниях ограничивающими рост могут быть низкие температуры и влажность воздуха и почвы, штормовые ветра. Влияние экологических факторов сказывается на подавлении роста надземных частей побегов (Serebryakov, 1962; Khokhryakov, 1994; Körner, 2016). Развитие подушковидных растений в различных систематических группах привело к многообразию их типов и форм (Rauh, 1939; Zajczeva, 1949; Richter, 1981; Moore, 1983). В настоящее время выделено и изучено морфологическое разнообразие основных типов подушек: травянистых, полудревесных, древесных, истинных и ложных, дерновинных и розеточных (Rauh, 1939; Serebryakov, 1962; Kul’tiasov, 1962; Mazurenko, Khokhryakov, 1977; Volkov, 2007; Körner et al., 2011; Kolegova, Cheryomushkina, 2014; Talovskaya, 2017). Если в литературе существует много сведений по биологии и механизмам формирования полудревесных и древесных подушек, то для травянистых растений имеются частные работы (Borisova, 1962; Antropova, 1983; Nukhimovskii, 1997; Pichugina, Savinykh, 2007; Zhmylev, 2000). Травянистые подушки относятся к хамефитам и/или гемикрипофитам, почки возобновления которых находятся выше поверхности или на уровне субстрата. Для трав-подушек в общем плане их строения и развития, так же как для прочих подушек, характерно акротонное интенсивное ветвление, незначительный годичный прирост, этажированность многолетних структур, компактное расположение осей и образование придаточных корней. Разнообразие и форма трав-подушек связаны главным образом с особенностями побегообразования, типом и цикличностью побегов возобновления. Так, у различных видов рода Saxifraga L. травянистые подушки (Zhmylev, 2004) строятся за счет развития моно-, ди-, или полициклических побегов розеточного или полурозеточного типов побегов с орто- и/или анизотропным направлением роста.
Биоморфологический анализ травянистых растений-подушек, с одной стороны, расширит представление об этой жизненной форме и ее эволюции (Boucher et al., 2016), с другой – позволит выявить механизм адаптации этих растений к суровым природным условиям. Наши исследования сосредоточены на изучении морфологического разнообразия представителей крупного рода Nepeta L. (Lamiaceae), виды которого в основном формируют травянистую, реже полудревесную жизненные формы (Astashenkov, 2015; Astashenkov et al., 2021). Среди изученных к настоящему времени видов только особи Nepeta pamirensis Franch. в высокогорных условиях Памира образуют подушковидную биоморфу (Astashenkov, 2015). Сравнительный анализ растений-подушек в системе биоморф близкородственных таксонов поможет выявить основные морфофилогенетические связи и пути соматической эволюции видов рода. Современные работы, связанные с изучением растений-подушек в условиях высокогорий, остаются без анализа их популяционной организации и оценки их состояния. Комплексное изучение популяционной биологии позволит заполнить этот пробел. В связи с этим цель работы – изучение особенностей формирования травянистой подушки и онтогенетической структуры ценопопуляции Nepeta lipskyi Kudrjasch. Образование подушки у представителей Nepeta на базе травянистого растения описывается впервые.
МАТЕРИАЛЫ И МЕТОДЫ
Материал по онтогенезу и структуре ценопопуляции Nepeta lipskyi собран на перевале Оббурдон (хр. Туркестанский, Таджикистан) в седловине гребня хребта (39°30'40.6"N 69°08'03.6"E, h = 3905 м над ур. м.) на сухом сильно ощебненном субстрате. Расположенная в криоксерофитных условиях на осевой пологой части хребта изученная популяция подвержена сильному иссушающему ветру и высокой инсоляции. Особи N. lipskyi образуют монодоминантную растительную группировку в составе формации альпийских низкотравных пустошей, проективное покрытие вида составляло 30%. В целом климат района исследования характеризуется как очень холодный (средняя годовая температура +0.35–1.01°С) умеренный (годовая амплитуда месячных температур 21–22°С) и с небольшим количеством осадков в течение года. Максимум осадков (не более 800 мм в год) выпадает в апреле, мае и июне. В разгар вегетации (июль, август) количество осадков резко уменьшается, и вследствие сильной инсоляции, частых ветров, сильной испаряемости и высокого нагревания каменистых почв создаются условия значительной сухости. Климат горных склонов на высотах около 4000 м сменяется суровым климатом нивальной области (Afanas’ev, 1956).
При описании жизненной формы использованы подходы, разработанные отечественными морфологами (Serebryakov, 1962; Serebryakova, 1977). Онтогенез изучен согласно представлениям Т.А. Работнова (Rabotnov, 1950), А.А. Уранова (Uranov, 1975). Организация побега охарактеризована с позиции его структурно-функциональной дифференциации, предложенной W. Troll (1964). При описании структуры растений применен архитектурный подход (Hallé, Oldeman, 1970; Serebryakova, 1977; Barthélémy, Caraglio, 2007). Для описания архитектуры подушки в работе использованы термины: модуль (Barthélémy, Caraglio, 2007), резид (Nukhimovskii, 1997). Календарный возраст особей, когда это представлялось возможным, установлен с помощью прямого подсчета сохранившихся ежегодных резидов и годичных колец на анатомическом срезе главного корня. Онтогенетическая структура ценопопуляции изучена по общепринятой методике (Tsenopopulyatsii…, 1988), применено представление о характерном онтогенетическом спектре (Zaugol’nova, 1994). В качестве счетных единиц выбраны особь семенного происхождения (p–g3) и парциальные образования (кусты и побеги) (g3–s). В ценопопуляции определено соотношение особей разных онтогенетических состояний. Онтогенетический спектр построен на основе учета особей на 45 площадках размером 1 м2, заложенных регулярным способом на трансектах шириной 1 м. Трансекты располагались вдоль и поперек осевой части хребта. При характеристике ценопопуляции использованы классификации по абсолютному максимуму онтогенетических групп (Uranov, Smirnova, 1969) и “дельта-омега” Л.А. Животовского (Zhivotovskii, 2001). Показатели экологической плотности (Pэкол) даны согласно Ю. Одуму (Odum, 1986).
РЕЗУЛЬТАТЫ
Nepeta lipskyi – симподиально нарастающее многолетнее травянистое растение, эндемик Средней Азии. Гемикриптофит. Произрастает на осыпях в составе криофильной растительности в границах альпийского и субальпийского поясов (3800–4000 м над ур. м.) горной системы Гиссаро-Алая (хр. Алайский, Туркестанский, Гиссарский) (Pojarkova, 1954; Budantzev, 1993) (рис. 1).
Рис. 1.
Травянистая подушка Nepeta lipskyi в условиях Туркестанского хребта.
Fig. 1. Herbaceous cushion of Nepeta lipskyi in the conditions of Turkestan Range.

ОНТОГЕНЕЗ
Онтогенез особей Nepeta lipskyi схематично изображен на рис. 2.
Рис. 2.
Онтогенез особей Nepeta lipskyi.
Онтогенетические состояния: p – проросток, j – ювенильное, im – имматурное, v – виргинильное, g1 – молодое генеративное, g2 – зрелое генеративное, g3 – старое генеративное, ss – субсенильное, s – сенильное; черта – уровень почвы, стрелки – переход из одного онтогенетического состояние в другое.
Fig. 2. Ontogeny of Nepeta lipskyi individuals.
Ontogenetic states: p – seedling, j – juvenile, im – immature, v – virginal, g1 – young generative, g2 – mature generative, g3 – old generative, ss – subsenile, s – senile; line – ground level, arrows – transition from one ontogenetic state to another.
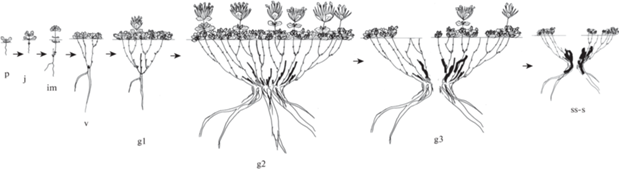
Прорастание семян надземное. Проросток имеет две семядоли и два настоящих супротивных листа. За счет контрактильной деятельности корня происходит втягивание семядольного узла с коротким гипокотилем в мелкоземистый субстрат. При этом эпикотиль вытягивается (0.3–0.4 см). В год прорастания семян особи, продолжая нарастать моноподиально, переходят в ювенильное состояние. Первичный побег ортотропный, верхнерозеточный, высотой не более 1.5 см. За вегетационный период развивается 2–3 пары накрест-супротивно расположенных настоящих черешковых листьев с городчатым краем. В пазухах всех листьев закладываются почки. Обычно на одном метамере супротивные почки имеют неодинаковое число зачатков, одна всегда наиболее развита. Длина главного корня не превышает 2 см. После вегетации розеточная часть побега отмирает, сохраняется семядольный узел с пазушными почками.
На второй год растения переходят в имматурное состояние. Особи однопобеговые. Моноподиальное нарастание сменяется на симподиальное. Из одной почки семядольного узла развертывается моноциклический ортотропный верхнерозеточный побег. Вторая почка становится спящей. У побега текущего прироста геофильная часть длиной 1.5 см. Она ориентирована ортотропно, состоит из коротких 2–3 метамеров с чешуевидными и 1–2 длинных метамеров с переходными листьями. Переходные листья сидячие, бурого цвета, листовые пластинки цельнокрайние, только верхушки слегка волнистые. Почки регулярного возобновления закладываются в пазухе верхнего чешуевидного или нижнего переходного листа. Остальные почки не реализуются и становятся спящими. Надземная часть побега несет 3–4 метамера со сближенными междоузлиями. Верхнерозеточный побег прижат к поверхности субстрата, его высота не превышает 1.5 см. Побег в надземной части интенсивно ветвится. Из каждой пазушной почки развивается побег обогащения длиной 0.5–0.9 см. В конце вегетации разветвленный верхнерозеточный побег засыхает, втягивается в субстрат и постепенно отмирает до почки регулярного возобновления. Сохранившийся ортотропный резид длиной не более 0.4 см дает начало формированию каудекса и становится основой для построения его главной оси. Главный корень начинает ветвиться. Длина главного корня достигает 5 см. Длительность состояния 1 год.
На следующий год особи переходят в виргинильное состояние. В этом состоянии формируется первичный куст. Он состоит из двух вегетативных моноциклических верхнерозеточных побегов. Побеги текущего прироста развертываются из почки, сохранившейся в семядольном узле, и почки регулярного возобновления, расположенной на резиде прошлогоднего побега. Геофильные части побегов возобновления 2.0–2.2 см дл, состоят из 1–3 коротких и 2–3 длинных метамеров с чешуевидными и переходными листьями. Начиная с этого состояния, почка регулярного возобновления формируется в пазухе нижнего переходного листа. Остальные почки не реализуются, они могут тронуться в рост при повреждении растущего побега. Розеточная часть побегов высотой 1.3–1.5 см состоит из 5–6 коротких метамеров. Из каждого узла развиваются боковые побеги обогащения длиной 1–1.5 см. После вегетации все листья засыхают. В субстрат втягивается разветвленная осевая часть каждого материнского побега, которая сохраняется в течение следующего года, затем отмирает до почки возобновления. Сохранившиеся резиды удлиненные, они имеют относительно одинаковую длину (0.8–1.0 см). Резид побега, возникшего из почки возобновления, надстраивает главную ось каудекса. Резид побега, возникшего из почки семядольного узла, дает начало боковой оси (каудикула I порядка). Базальная часть каудекса разрастается до 0.4–0.8 см в диаметре. Главный и боковой корни утолщаются (0.4–0.5 см), достигают длины 8–10 см. Покровные ткани каудекса и главного корня начинают слущиваться. Длительность онтогенетического состояния 1–2 года.
Первое цветение приходится на 4–5 год, особи переходят в молодое генеративное состояние. Начинает формироваться подушка. В ее основе лежат выровненная длина резидов, одинаковая высота плотно прижатых к субстрату ветвящихся верхнерозеточных побегов, тесное расположение друг к другу ортотропных осей с незначительным годичным приростом. В молодой подушке различают вегетативные побеги разного происхождения: 1) побеги, развившиеся из почки возобновления, расположенные на каудикуле I порядка; 2) побеги, развившиеся из спящих почек, сохранившихся на первом резиде главной оси и каудикуле I порядка. У особей вегетативные верхнерозеточные побеги имеют облик побегов имматурных и виргинильных особей. Каждый вегетативный побег, развернувшийся из спящей почки, дает начало новой боковой оси каудекса (каудикула n+1 порядка). Так, после отмирания надземных частей таких побегов сохраняются резиды с почками регулярного возобновления. Нижележащие почки становятся спящими. На следующий год из одной перезимовавшей почки каждого резида формируется вегетативный побег. После его вегетации сохранившийся удлиненный ортотропный резид продолжает симподиально надстраивать ось каждой каудикулы. В кусте первым зацветает побег, развившийся из почки регулярного возобновления верхнего резида главной оси. Надземная часть генеративного побега высотой (вместе с соцветием) 5.8–6.0 см, она состоит из метамеров с удлиненными междоузлиями в отличие от вегетативных побегов. В результате этого соцветие выносится высоко над поверхностью подушки. Соцветие представляет собой отрытый головчатый тирс из 4–5 близкорасположенных дихазиев, составленных из монохазиев. Геофильная часть генеративного побега не превышает 2.5 см дл., несет 2–3 коротких и 2–3 длинных метамера с чешуевидными и переходными листьями. Почки, расположенные в геофильной части побега, слабо развиты и не участвуют в побегообразовании. В конце вегетационного сезона генеративный побег засыхает и отмирает. В результате акронекроза каудикула начинает постепенно разрушаться. Ежегодно отмирает каждый последующий нижележащий резид каудикулы, живым остается только первый резид оси. Базальная часть каудекса за счет сохранения первых резидов разных каудикул со спящими почками увеличивается в диаметре (2.5–3.0 см). На ней появляются многолетние придаточные корни. Главный корень достигает длины 15 см, интенсивно ветвится, образуются однолетние эфемерные корни. В этом состоянии начинается процесс расщепления главного корня. Он разрушается с апикального конца вдоль сосудов ксилемы. В общей подземной части подушки из-за одинакового метамерного расположения почки регулярного возобновления (всегда первый метамер с переходными листьями) и выровненной длины сохраняющихся резидов (1.4–1.5 см), строится многолетняя этажированная структура. Диаметр подушки не превышает 10 см. Длительность состояния 3–4 года.
В зрелом генеративном состоянии особи N. lipskyi представляют собой плоскую, округлую в очертании подушку диаметром 40–50 см. Побегообразование осуществляется за счет почек возобновления и спящих почек. В подушке развиваются 30–50 генеративных и большое число вегетативных (100–150) побегов, розеточные части которых приземистые и плотно прижаты друг к другу. Развертывание спящих почек, расположенных в базальной части каудекса, приводит к появлению большого числа слаборазвитых вегетативных побегов, которые заполняют надземную сферу плоской подушки и приводят к ее радиальному разрастанию. Фотосинтезирующая часть таких побегов состоит из 3–5 коротких метамеров с зелеными листьями, геофильная часть – из 2–3 коротких и 1–3 длинных метамеров. Большая часть вегетативных побегов осенью отмирает и в дальнейшем они не участвуют в построении многолетних осей. Вегетативные побеги, появившиеся из почек возобновления, более развиты, длина их геофильной и надземной частей варьирует от 1.5 до 2.0 см и 3.0–4.0 см соответственно. Генеративные побеги развиваются только из почек возобновления последних годичных приростов каждой каудикулы. Высота побегов – 6.5–7.5 см, длина геофильной части – 2–2.5 см. Сохранившиеся ортотропные резиды имеют выровненную длину (1.6–1.7 см), они продолжают строить ярусную подземную структуру подушки. Некроз многолетних осей приводит к накоплению в кусте большого числа отмерших остатков каудикул. Базальная часть разрастается до 4–7 см. Главный корень длиной до 25 см, он веретенообразно перекручен, его базальная часть расширенная. Многолетние боковые и придаточные корни утолщаются до 1.0–1.5 см и достигают длины 20 см. На поверхности корней образуется большое число эфемерных корней разного порядка. Длительность состояния – 10–12 лет.
В старом генеративном состоянии формируется клон. Отмирание первых резидов старых осей и расщепление главного корня приводят к полной партикуляции особи. В результате этого обособляются первичный и парциальные кусты, имеющие собственную корневую систему. Подушка становится рыхлой и имеет неправильную в очертании форму. Клон состоит из первичного куста и неомоложенных парциальных образований, которые медленно стареют вместе с первичным кустом. Побегообразование осуществляется как за счет почек возобновления, так и спящих почек. В первичном и парциальных кустах формируется большое число вегетативных (более 50) и незначительное число генеративных (не более 10) побегов. В конце этого состояния возможно образование только одного генеративного и 5–15 вегетативных побегов. У некоторых парциальных образований последний годичный прирост каудикулы заканчивается побегом с неполным циклом развития, в связи с этим особи оказываются временно не цветущими. Большая часть побегов, возникших из спящих почек, не выходит на поверхность субстрата. В течение короткого вегетационного периода у них образуется только геофильная часть, несущая чешуевидные листья. Эти побеги отмирают и не участвуют в построении новых каудикул. Из-за разрушения целостности особи в надземной сфере подушки образуются свободные пространства. Отдельные парциальные кусты могут располагаться друг от друга на расстоянии более 40–50 см. В результате полной партикуляции базальная часть каудекса также разрушается. Длительность состояния не превышает 4–6 лет.
Особи постгенеративного периода представляют собой рыхлую подушку (клон), состоящую из первичного и одного парциального куста с небольшим числом (5–7) побегов виргинильного облика. Побегообразование осуществляется исключительно из сохранившихся спящих почек. За счет большого числа спящих почек и медленного разрушения особи долгое время могут находиться в этом состоянии. У неомоложенных парциальных образований возможна повторная партикуляция. Разрушение происходит в центральной части куста в результате отмирания многолетних резидов каудекса и повторного расщепления придаточного корня. Длительность состояния 3–4 года.
Таким образом, онтогенез особей N. lipskyi полный, сложный. Процесс партикуляции растянутый, полный распад особи приходится на старое генеративное состояние. В результате медленного распада образуются неомоложенные долгоживущие парциальные образования. По темпам развития средневозрастное онтогенетическое состояние самое продолжительное.
Архитектурный анализ
Используя архитектурный подход, мы выявили особенности строения и развития многолетней структуры Nepeta lipskyi в условиях высокогорий Туркестанского хребта. В качестве конструкционных единиц выбраны: модуль (Barthélémy, Caraglio, 2007) и резид (Nukhimovskii, 1997). Так, после отмирания первичного побега, возникшего из семени, из одной почки семядольного узла формируется замещающий разветвленный вегетативный модуль (рис. 3). После отмирания его надземной части сохраняется удлиненный резид, который становится основой для построения первой оси (главная ось). Первая ось развивается за счет симподиального линейного сочленения удлиненных резидов разветвленных в надземной части модулей. Каждый год формируется один модуль замещения. Развитие модуля происходит из почки длинного метамера, несущего переходные листья. Нижележащие почки (в пазухах коротких метамеров) становятся спящими. Особенность структуры многолетней оси заключается в том, что многолетняя ось строится резидами только вегетативных модулей. На 4–5 год из почки верхнего резида развивается один генеративный модуль. После его развития вся симподиальная ось начинает постепенно разрушаться. Многолетняя структура оси разрушается до первого резида с расположенными на нем спящими почками.
Рис. 3.
Архитектурный анализ особей Nepeta lipskyi.
1 – разветвленный в надземной части вегетативный модуль, 2 – разветвленный в надземной части генеративный модуль, 3 – сохраняющаяся подземная часть модуля (резид), 4 – почка возобновления, 5 – почка спящая. Число почек условное, узлы расставлены условно.
Fig. 3. Architectural analysis of Nepeta lipskyi individuals.
1 – vegetative module branched in the above-ground part, 2 – generative module branched in the above-ground part, 3 – preserved underground part of the module (residue), 4 – renewal bud, 5 – dormant bud. The number of buds is conditional, the nodes are arranged conditionally.
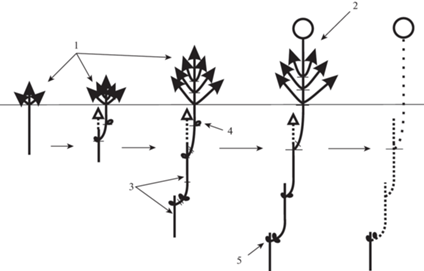
Основой построения боковых осей (каудикул) выступают резиды модулей, возникших из спящих почек. Механизм формирования новых осей происходит также по линейному симподиальному способу. Как и главная, любая боковая ось строится резидами только вегетативных побегов. На 3–4 год из одной (реже двух) почки последнего верхнего резида развивается один генеративный модуль. В конце вегетационного сезона, после отмирания модуля, боковая ось начинает разрушаться до первого резида. Такой паттерн построения боковых осей сохраняется в течение всего онтогенеза особей. С возрастом у растений сокращается число резидов в составе оси.
Онтогенетическая структура ценопопуляции
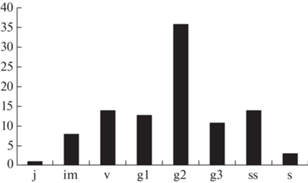
Исходя из особенностей онтогенеза и семенного способа возобновления, характерный онтогенетический спектр Nepeta lipskyi центрированный, полночленный. В условиях высокогорий Туркестанского хребта особи N. lipskyi образуют монодоминантную группировку с полным составом всех онтогенетических состояний. Онтогенетический спектр ценопопуляции совпадает с характерным спектром. Спектр центрированный, полночленный, с абсолютным максимумом на средневозрастных генеративных растениях (36%) (рис. 4). Это связано с долгим нахождением особей в зрелом генеративном состоянии и является биологической особенностью этого вида. Распределение онтогенетических фракций в левой части спектра отражает успешное семенное возобновление, выровненные темпы развития особей и перехода из одного онтогенетического состояния в другое. Правая часть спектра отражает процесс старческой партикуляции особей в конце генеративного периода. Замедленный процесс разрушения, который сопровождается образованием клона, с одной стороны, обуславливает появление старых жизнеспособных особей в ценопопуляции (g3–11; ss–14%), с другой, приводит к растянутым темпам развития старых особей и, как следствие, их накоплению. По классификации “дельта-омега” Л.А. Животовского (Zhivotovskii, 1991) ценопопуляция переходная, приближающаяся к зрелой (Δ = 0.47; ω = 0.69). В изученной ценопопуляции пространственное распределение особей вдоль осевой части хребта носит равномерный характер. Средняя экологическая плотность составляла 4.7 ± 2.4 экз./м2. Наибольшая плотность (9–11 экз./м2) отмечена на площадках, где преобладали клоны, состоящие из большого числа неомоложенных парциальных образований (ss) и старых особей семенного происхождения (g3). Низкая плотность (1–3 экз./м2) отмечалась на выдуваемых буграх и складывалась преимущественно за счет растений прегенеративного периода и одиночных растений зрелого генеративного состояния.
Рис. 4.
Онтогенетический спектр ценопопуляции Nepeta lipskyi.
По оси Х – онтогенетические состояния, по оси Y – количество особей на единицу площади, % от общего числа.
Fig. 4. Ontogenetic spectrum of Nepeta lipskyi coenopopulation.
X-axis – ontogenetic states; Y-axis – the number of individuals per area unit, % of total number.
Таким образом, онтогенетический состав, индексы возрастности и эффективности отражают успешный оборот поколений, выровненные темпы онтогенеза и нормальное устойчивое развитие ценопопуляции N. lipskyi в сложившихся высокогорных условиях.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Развитие особей Nepeta lipskyi существенно отличается от известных на сегодняшний день вариантов побегообразования многолетних каудексообразующих трав (Serebryakovа, 1977, Nukhimovskii, 1997; Aktual’nye…, 2012) и описанных механизмов формирования травянистых подушек. Так, в отличие от большинства описанных каудексовых растений и трав-подушек, в побегообразовании которых обычно участвует почки возобновления, у особей N. lipskyi, помимо почек регулярного возобновления, в побегообразовании участвуют спящие почки. Пробуждение спящих почек у особей N. lipskyi дает начало построению новых многолетних осей каудекса. Такое побегообразование характерно для полудревесных и некоторых древесных растений (кустарников, кустарничков) (Mazurenko, Khokhryakov, 1977; Cheryomushkina, Guseva, 2015; Talovskaya et al., 2018), но не для трав (Serebryakov, 1962). Отличие в побегообразовании и в механизме формирования подушки отмечается у близкородственного вида N. pamirensis, произрастающего в условиях высокогорий Восточного Памира. Так, в отличие от особей N. lipskyi, подушка особей N. pamirensis развивается на основе полукустарничка в результате силлептического ветвления базальной части материнского побега. Побеги возобновления развертываются из почек, сохранившихся на базальных частях побегов II порядка прошлогоднего прироста. Интенсивное ветвление близкорасположенных побегов и ежегодное симподиальное надстраивание одинаковыми базальными частями приводит к формированию плотной выпуклой подушки. Развертывание побегов из почек удлиненного метамера, находящегося выше поверхности почвы, приводит к росту подушки в высоту, а из почек укороченных метамеров – к ее разрастанию по периферии.
Помимо этого, механизм формирования травянистых и большей части древесных и полудревесных подушковидных жизненных форм связан с розеточным или полурозеточным типом побега (Rauh, 1939, Serebryakov, 1962; Körner, 2016). Для особей N. lipskyi, как и для большинства представителей рода Nepeta, характерен удлиненный тип побега. Однако влияние низких температур и сильного иссушающего ветра привели к сокращению длины междоузлиев надземной части, что отразилось на структуре побегов особей N. lipskyi и обусловило “сжатие” верхушки побегов до розетки. Такой тип побега можно характеризовать как верхнерозеточный, и мы его рассматриваем как следствие аббревиации длины метаметров с листьями срединной формации под влиянием высокогорных условий.
В целом механизм формирования подушек у изученных ранее травянистых видов растений подобен подушкам древесных и полудревесных жизненных форм (Borisova, 1962; Nukhimovskii, 1997; Zhmylev, 2004; Pichugina, Savinykh, 2007). Построение плоской подушки у особей N. lipskyi – это результат выровненной длины резидов; одинаковой высоты плотно прижатых к субстрату интенсивно ветвящихся верхнерозеточных побегов; этажированного ветвления многолетних структур за счет одинакового метамерного расположения почки регулярного возобновления; близкого расположения осей с незначительным годичным приростом. В отличие от известных травянистых подушек, у которых накапливается ветошь от годичных приростов внутри подушки, у особей N. lipskyi вся надземная часть ежегодно отмирает и сдувается ветром, а ветошь накапливается в субстрате между каудикулами.
В побегообразовании у N. lipskyi установлен эффект “усиления побегов”, описанный Т.И. Серебряковой (Serebryakovа, 1977). Она отмечала, что усиление побегов у растений с удлиненным типом побега наблюдается только на начальных этапах онтогенеза особи, и после первого цветения у растений ежегодно формируются хорошо развитые генеративные побеги замещения. У особей N. lipskyi эффект “усиления побегов” наблюдается в процессе формирования многолетней оси (каудикулы) (рис. 3). Такой эффект повторяется каждый раз при образовании новой боковой оси и сохраняется в течение всего онтогенеза особей. Так, каждый последующий побег строящейся оси более развит, чем предыдущий. Таким образом, архитектура подушки N. lipskyi определяется наличием многолетних осей, формирующихся в результате линейного симподиального сочленения резидов вегетативных модулей.
Онтогенетическая структура ценопопуляции N. lipskyi отражает устойчивое ее развитие в условиях монодоминантного котовникового сообщества на Туркестанском хребте. Онтогенетический спектр ценопопуляции совпадает с характерным. В отсутствие фитоценотического давления динамические процессы, происходящие в ценопопуляции, обусловлены выровненными темпами развития семенных особей и парциальных образований, сбалансированным оборотом поколений в результате растянутого процесса партикуляции и постоянной инспермации. Подобное популяционное поведение отмечено у некоторых видов рода Allium L., образующих монодоминантные группировки на открытых перевеваемых песчаных барханах (Cheryomushkina, 2004), у которых онтогенетическая структура, как и у Nepeta lipskyi определяется только особенностями онтогенеза, способом возобновления и влиянием экологических факторов.
ЗАКЛЮЧЕНИЕ
Онтогенез особей N. lipskyi сложный, полный. Каудекс формируется на начальных этапах онтогенеза в результате симподиального сочленения резидов. По темпам развития наиболее продолжительное состояние средневозрастное. В побегообразовании принимают участие почки регулярного возобновления и спящие почки. В течение онтогенеза у особей N. lipskyi формируются моноциклические вегетативные верхнерозеточные побеги и генеративные побеги с удлиненными междоузлиями.
Плотно расположенные друг к другу надземные части верхнерозеточных побегов из-за одинакового метамерного расположения почки регулярного возобновления имеют выровненную высоту (3.0–4.0 см) и прижаты к субстрату, это приводит к образованию плоской травянистой подушки. Ежегодный незначительный годичный прирост (длина резида 1.6–1.7 см) многолетних осей (каудикул) и одинаковое расположение почки регулярного возобновления сопровождается этажированным ветвлением. Удлиненные междоузлия генеративных побегов выносят верхушечное соцветие над поверхностью подушки.
Многолетние оси каудекса строятся по монохазиальному типу. Длительность жизни каждой оси не превышает 3–5 лет. В построении оси участвуют резиды вегетативных побегов, заканчивается ось всегда генеративным побегом, после чего ось разрушается. Онтогенетический спектр ценопопуляции центрированный, полночленный, совпадает с характерным спектром. Индексы возрастности и эффективности (Δ = 0.47; ω = = 0.69) характеризуют ценопопуляцию как переходную к зрелой. На основании онтогенетического состава и демографических показателей изученная ценопопуляция характеризуется как устойчивая, с нормальным типом развития.
Список литературы
[Afanas’ev] Афанасьев К.С. 1956. Растительность Туркестанского хребта. М., Л. 277 с.
[Aktual’nye…] Актуальные проблемы современной биоморфологии. 2012. Киров. 610 с.
[Antropova] Антропова Г.Л. 1983. Трансформация формы роста травянистых подушковидных лапчаток северо-востока СССР. – В кн.: Тезисы докл. VII делегатского съезда всесоюзного ботанического общества. Л. С. 225.
Astashenkov A.Yu. 2015. Morphological adaptation of Nepeta pamirensis Franch. (Lamiaceae) to the conditions of the Pamir mountains. – Contemporary Problems of Ecology 8 (5): 636–646. https://doi.org/10.1134/S1995425515050029
Astashenkov A.Yu., Karpova E.A., Cheryomushkina V.A. 2021. Diversity patterns of life forms and phenolic profiles of endemic Nepeta plants along an aridity gradient of a high-mountain zone in Central Asia. – Taiwania. 66 (4): 541–556. https://doi.org/10.6165/tai.2021.66.541
Aubert S., Boucher F., Lavergne S., Renaud J., Choler P. 2014: A revised worldwide catalogue of cushion plants 100 years after Hauri and Schröter. – Alpine Botany. 124 (1): 59–70. https://doi.org/10.1007/s00035-014-0127-x
Barthélémy D., Caraglio Y. 2007. Plant architecture: a dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. –Ann. Bot. 99 (3): 375–407. https://doi.org/10.1093/aob/mcl260
[Borisova] Борисова И.В. 1962. Биолого-морфологическая характеристика травянистых подушковидных растений Северного Казахстана. – Проблемы ботаники. 6: 336–345.
Boucher F.C., Lavergne S., Basile M., Choler P., Aubert S. 2016. Evolution and biogeography of the cushion life form in angiosperms. – Perspectives in Plant Ecology, Evolution and Systematics. 20: 22–31. https://doi.org/10.1016/j.ppees.2016.03.002
[Budantzev] Буданцев А.Л. 1993. Триба Nepeteae Benth. семейства Lamiaceae Lindl. (систематика, география, возможности использования): Автореф. дис. … док. биол. наук. СПб. 33 с.
Butterfield B.J., Cavieres L.A., Callaway R.M., Cook B.J., Kikvidze Z., Lortie C.J., Michalet R. et al. 2013. Alpine cushion plants inhibit the loss of phylogenetic diversity in severe environments. – Ecol. Lett. 16: 478–486. https://doi.org/10.1111/ele.12070
[Cheryomushkina] Черемушкина В.А. 2004. Биология луков Евразии. Новосибирск. 279 с.
Cheryomushkina V.A., Guseva A.A. 2015. Life forms of Scutellaria supina L. (Lamiaceae). – Contemporary Problems of Ecology. 8 (5): 624–635. https://doi.org/10.1134/S1995425515050054
Hallé F., Oldeman R.A.A. 1970. Essai sur l architecture et la dynamique de croissance des arbres tropicaux. Paris. 178 p.
Hauri H., Schröter C. 1914. Versuch einer Ubersicht der siphonogamen Polsterpflanzen. – Bot. Jahrb. Syst. Pflanzengesch. Pflanzengeogr. 50:618–656.
[Khokhryakov] Хохряков А.П. 1994. “Подушки” для экстремальных условий. – Природа. 6: 36–41.
[Kolegova, Cheryomushkina] Колегова Е.Б., Черемушкина В.А. 2014. Онтогенез подушковидной жизненной формы Thymus baicalensis (Lamiaceae). – Бот. журн. 99 (10): 1109–1118.
Körner C. 2003. Alpine plant life: functional plant ecology of high mountain ecosystems, 2nd ed. Springer, New York. 345 p.
Körner C., Paulsen J., Spehn E.M. 2011. A definition of mountains and their bioclimatic belts for global comparisons of biodiversity data. – Alpine Botany. 121: 73–78. https://doi.org/10.1007/s00035-011-0094-4
Körner C. 2016. Plant adaptation to cold climates [version 1; peer review: 2 approved]. F1000Research 5 (F1000 Faculty Rev): 2769. https://doi.org/10.12688/f1000research.9107.1
[Kul’tiasov] Культиасов И.М. 1962. Эколого-морфологические особенности подушковидных эспарцетов Средней Азии. – Бот. журн. 47 (5): 645–656.
[Mazurenko, Khokhryakov] Мазуренко М.Т., Хохряков А.П. 1977. Структура и морфогенез кустарников. М. 158 с.
Moore D.M. 1983. Flora of Tierra del Fuego. Missouri Botanical Garden, St-Louis. 396 p.
[Nukhimovskii] Нухимовский Е.Л. 1997. Основы биоморфологии высших растений. Т. 1. М. 630 с.
[Odum] Одум Ю. 1986. Экология. Т. 2. М. 376 с.
[Pichugina, Savinykh] Пичугина Е.В., Савиных Н.П. 2006. Особенности онтогенеза Jurineae cyanoides (Asteraceae) на северной границе ареала. – Раст. ресурсы. 2006. 3: 10–25.
[Pojarkova] Пояркова А.И. 1954. Семейство Lamiaceae. – В кн.: Флора СССР. Т. 20. М., Л. С. 286–360.
[Rabotnov] Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах. – Труды БИН АН СССР. Геоботаника. 3 (6): 176–196.
Rauh W. 1939. Uber polsterförmigen Wuchs. Ein Beitrag zur Kenntis der Wuchsformen der hoheren Planzen. – Nova Acta Leopold. 7: 268–508.
Raunkiær C. 1934. The life forms of plants and statistical plant geography, being the collected papers of C. Raunkiær. Oxford. 632 p.
Richter M. 1981. Klimagegensätze in Südperu und ihre Auswirkungen auf die Vegetation. Erdkunde. 35 (1): 12–30. https://doi.org/10.3112/erdkunde.1981.01.02
[Serebryakov] Серебряков И.Г. 1962. Экологическая морфология растений. М. 378 с.
[Serebryakova] Серебрякова Т.И. 1977. Об основных “архитектурных моделях” травянистых многолетников и модусах их преобразования. – Бюл. МОИП. Отд. биол. 5 (82): 112–128.
[Stanyukovich] Станюкович К.В. 1949. Растительный покров Восточного Памира. М. 159 с.
[Steshenko] Стешенко А.П. 1965. Ритм развития растений Памира в связи с различиями условий среды. – Проблемы современной ботаники. 2: 111–115.
[Talovskaya] Таловская Е.Б. 2017. Поливариантность онтогенеза Thymus mugodzharicus (Lamiaceae) подушковидной жизненной формы. – Вестник Томского гос. ун-та. Биология. 40: 88–101. https://doi.org/10.17223/19988591/40/5
Talovskaya E.B, Cheryomushkina V.A., Denisova G.R. 2018. Morphological adaptation of species of the genus Thymus (Lamiaceae) in Yakutia. – Contemporary Problems of Ecology. 6: 624–634. https://doi.org/10.1134/S1995425518060094
Troll W. 1964. Die Infloreszenzen, Typologie und Stellung im Aufbau des Vegetationskörpers. Bd. 1. Jena. 615 p.
[Tsenopopulyatsii…] Ценопопуляции растений (очерки популяционной биологии). 1988. М. 182 с.
[Udalova] Удалова Р.А. 1978. Морфологический анализ некоторых подушковидных кактусов. – Бот. журн. 63 (2): 256–263.
[Uranov] Уранов А.А. 1975. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов. – Биол. науки. 2: 7–34.
[Uranov, Smirnova] Уранов А.А., Смирнова О.В. 1969. Классификация и основные особенности развития популяций многолетних растений. – Бюл. МОИП. Отд. биол. 74 (2): 119–134.
[Volkov] Волков И.В. 2007. Биоморфологические адаптации высокогорных растений. Томск. 412 с.
[Zajczeva] Зайцева М.Г. 1949. О развитии формы растений-подушек. – Сообщ. Таджик. Фил. АН СССР. Сталинобад. Т. 17. С. 16–20.
[Zaugol’nova] Заугольнова Л.Б. 1994. Структура популяций семенных растений и проблемы их мониторинга: Автореф. дис. … док. биол. наук. СПб. 70 с.
[Zhmylev] Жмылев П.Ю. 2000. Жизненные формы камнеломок в связи с эволюцией рода Saxifraga L. (Saxifragaceae). – Бюл. МОИП. Отд. биол. 105 (6): 32–37.
[Zhmylev] Жмылев П.Ю. 2004. Род Saxifraga L. (Saxifragaceae): Биоморфология, систематика и эволюция жизненных форм: Автореф. дис. … док. биол. наук. М. 45 с.
[Zhivotovskii] Животовский Л.А. 2001. Показатель сходства популяций по полиморфным признакам. – Журн. общ. биол. (40) 4: 587–602.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал