

Ботанический журнал, 2022, T. 107, № 3, стр. 211-236
СВЯЗЬ ФИТОЦЕНОТИЧЕСКОГО РАЗНООБРАЗИЯ СЕВЕРОВОСТОЧНО-ЗАБАЙКАЛЬСКОГО ОРОБИОМА С БИОКЛИМАТИЧЕСКИМИ ПОКАЗАТЕЛЯМИ
М. В. Бочарников *
Московский государственный университет имени М.В. Ломоносова
119991 Москва, Ленинские горы, 1, Россия
* E-mail: maxim-msu-bg@mail.ru
Поступила в редакцию 19.03.2021
После доработки 05.12.2021
Принята к публикации 14.12.2021
- EDN: QANCIL
- DOI: 10.31857/S0006813622030024
Аннотация
Важнейшие пространственные закономерности растительного покрова Северного Забайкалья, региональные особенности которого отражаются в структуре высотной поясности Северовосточно-Забайкальского оробиома, определены в связи с климатическим фактором. Дана региональная климатическая оценка высотно-поясной структуры растительного покрова оробиома для типологических подразделений высшего иерархического уровня (фратрии классов растительных формаций) с использованием глобальной климатической модели BioClim. Метод главных компонент, дискриминантный и кластерный анализ определили за летним омбротермическим индексом, индексами континентальности и увлажненности климата, средним многолетним годовым количеством осадков наибольшую значимость среди биоклиматических показателей, определяющих пространственную структуру растительного покрова горнотаежного, подгольцового, горнотундрового и гольцового поясов. Фоновые сообщества поясов оробиома развиваются в условиях континентального климата (индекс континентальности от 36 до 50). Показатели теплообеспеченности, прежде всего, вегетационного периода, являются определяющими в пространственной дифференциации высотных подпоясов, характеризуя смену редколесных и стланиковых сообществ в подгольцовом поясе, лиственничных, лиственнично-сосновых и темнохвойных лесов в горнотаежном поясе. С влагообеспеченностью связаны региональные различия типологического разнообразия поясов, выраженные в развитии более требовательных к увлажнению пихтово-еловых лесов в низкогорьях Патомского нагорья (количество осадков более 450 мм в год), и сосновых лесов в межгорных котловинах Станового нагорья, существующих в условиях с наибольшей годовой амплитудой температур и небольшого количества осадков (до 400 мм в год).
Уровни флористического и фитоценотического разнообразия, характеризуя региональное своеобразие биомов как совокупности природных экосистем, являются интегральным выражением эколого-географических факторов его формирования, на современном этапе поддерживаемого, прежде всего, климатическими условиями. Определение ведущих климатических градиентов и анализ отклика различных характеристик растительного покрова на изменение биоклиматических показателей является предметом фундаментальных исследований как отдельных видов растений (Popov, 2018), так типологических подразделений растительности (Gavilan, 2005) в разных масштабах и с использованием широкого спектра методов при проведении климатической ординации.
Разнообразие климатических условий и их отражение в пространственной организации фитоценотического разнообразия горных территорий связано со сложным сочетанием широтных и долготных факторов дифференциации среды в системе высотных градиентов условий. Совокупное отражение комплекса эколого-географических закономерностей представлено в биоклиматических моделях растительности (Rivas-Martinez, 1996; Nazimova et al., 2004), типологических схемах высотной поясности растительности (Ogureeva, 1999), в районировании территории с учетом связей растительности и климата (Polikarpov et al., 1986; Rivas-Martinez et al., 2004). Климатические условия выступают ключевыми факторами формирования региональной структуры биоразнообразия равнин и горных территорий (Ogureeva, Bocharnikov, 2017; Map…, 2018).
Пространственная структура фитоценотического разнообразия растительного покрова Северного Забайкалья в мелкомасштабных исследованиях прослеживается через дифференциацию ключевых биоклиматических показателей: средней годовой температуры, среднего годового количества осадков (Tuhkanen, 1984), индексов континентальности климата (Nakamura, Krestov, 2007). С показателями продуктивности сообществ находят корреляционные зависимости показатели суммы активных температур (Buks, 1976), радиационного баланса (Golubyatnikov, Denisenko, 2004). Региональный уровень исследования, на котором выделяются оробиомы, требует особого подхода к оценке разнообразия и структуры биоты с учетом высотных градиентов биоклиматических показателей. Под их воздействием формируется структура поясности, с пространственной дифференциацией которой связано выделение типов высотной поясности растительности, их подтипов и вариантов, а также формирование специфики состава и структуры конкретных высотных поясов, что находит отражение в разнообразии региональных оробиомов и их географических вариантов.
Разнообразие растительного покрова Северного Забайкалья определяется развитием нескольких фратрий классов растительных формаций (Sochava, 1964) в условиях выраженной высотно-поясной организации, свойственной Забайкальской группе типов поясности растительности (Ogureeva, 1999). Разнообразие и структура растительного покрова региона находит интегральное выражение в Северовосточно-Забайкальском оробиоме, представленном совокупностью экосистем, адаптированных к сформированной высотно-поясной структуре, и имеющем сложившийся уровень ботанического разнообразия в пределах поясов. Специфика исторического развития и современных условий, прежде всего, климатических, обуславливает единство оробиома (Map…, 2018).
Для обоснования географии фитоценотического разнообразия Северовосточно-Забайкальского оробиома проведен комплексный анализ климатических условий развития типологических подразделений растительности.
МАТЕРИАЛЫ И МЕТОДЫ
В основу исследования положена концепция биомного разнообразия. При многочисленных трактовках термина “биом” как крупных экосистем регионального уровня и подходах к их выделению и оценке биоразнообразия (Mucina, 2019) мы основываемся на классификационной схеме биомного разнообразия, наиболее полно раскрывающей ее структуру для равнин – через зональную структуру, и для горных территорий – с учетом высотно-поясных закономерностей организации биоты (Walter, Breckle, 1991). В соответствии с данными представлениями составлена карта “Биомы России” (Map…, 2018). Единство биомов гор (оробиомов) определяется разнообразием и структурой растительного покрова в пределах полного высотно-поясного спектра, который сформирован в результате длительного эволюционного развития и испытывает воздействие современных высотных градиентов, прежде всего, климатического (Ogureeva, Bocharnikov, 2017; Bocharnikov et al., 2018). В качестве модельного взят Северовосточно-Забайкальский оробиом, ботаническое разнообразие которого, связанное с его формированием в пределах Байкало-Джугджурской природной области (Sochava, 1980), характеризуется высотно-поясными закономерностями Забайкальской группы типов поясности растительности (Ogureeva, 1991).
Типологическое разнообразие растительного покрова представлено с применением географо-генетической классификации, на высших иерархических уровнях раскрывающей региональную специфику растительности с учетом ее происхождения и географической общности (Sochava, 1964). Оценка пространственной организации растительности в связи с климатическими условиями проведена на основе обзорной среднемасштабной карты растительности юга Восточной Сибири (М. 1 : 1500000), составленной на географо-генетической основе (Belov, 1973). Использована цифровая версия фрагмента карты на область развития Северовосточно-Забайкальского оробиома, созданная с помощью полиномиальной модели 6-й степени и географической привязки в системе UTM WGS-84. В ее легенде нашли отражение типологические единицы уровня растительных формаций, классов и групп ассоциаций. Объединения классов формаций в рамках фратрий как флороценогенетических подразделений растительности регионального уровня (Sochava, 1979, 1980) и высотных поясов и подпоясов, отражающих эколого-географические связи растительности по высотному градиенту, послужили опорными единицами анализа растительного покрова оробиома в связи с климатическими условиями. Это позволило дать современное обоснование типологического разнообразия растительности, которое формировалось в ходе длительного сопряженного исторического развития.
Для оценки пространственной структуры растительного покрова Северовосточно-Забайкальского оробиома предложен алгоритм, который направлен на выявление ее связей с биоклиматическими показателями, характеризующими общие пространственные закономерности развития биоты и эколого-географическое разнообразие растительного покрова в связи с комплексом градиентов факторов. Анализ включает в себя несколько этапов:
1) выявление градиентов климатических условий в связи со сложным орографическим строением горной территории Северного Забайкалья, ее высотно-поясной и секторной дифференциацией;
2) выявление связей типологического разнообразия растительности оробиома с климатическими условиями через конкретные биоклиматические показатели и определение среди них наиболее значимых для формирования растительности на региональном уровне;
3) оценка сопряженности пространственной дифференциации высотных градиентов и структуры растительного покрова оробиома.
В качестве основы для анализа пространственной дифференциации климатических условий взята глобальная цифровая климатическая модель BioClim (Hijmans et al., 2005), представленная набором растровых слоев ключевых биоклиматических показателей, характеризующих тепло- и влагообеспеченность, а также ряда комплексных показателей, основанных на соотношении тепла и влаги. Модель создана на основе данных метеостанций, осредненных за период 1970–2000 гг. и их интерполяции с учетом морфометрических показателей рельефа (Fick, Hijmans, 2017). Пространственное разрешение модели (30") послужило опорным уровнем анализа связей растровых слоев растительности и климата (статистический анализ в границах области развития оробиома проводился для 61203 пикселей).
Из биоклиматических показателей при анализе использованы средние многолетние, максимальные и минимальные средние многолетние значения температур воздуха, среднее многолетнее количество осадков по месяцам, сезонам и за год, а также комплексные показатели – изотермический коэффициент, показатель сезонности температур. На их основе был рассчитан ряд коэффициентов. Их выбор в качестве ключевых показателей определен опытом выявления географических закономерностей пространственной организации и разнообразия растительности, проявляющихся как на планетарном уровне (Rivas-Martinez et al., 1999; Walter, 1985; Box, 1995), так и для различных регионов, в том числе горных (Major, 1963; Yim, Kira, 1975; Valencia-Barrera et al., 2002; Krestov, 2007). Переменные модели использованы при расчете ряда ключевых в определении структуры растительности коэффициентов, которые применяются при ее климатической ординации на разных пространственных уровнях. Они успешно апробированы при биоклиматическом обосновании пространственной структуры растительного покрова и проведении мелко- и среднемасштабного биоклиматического и биогеографического районирования (Rivas-Martinez, Armaiz, 1984; Rivas-Martinez et al., 2004, 2011; Nakamura et al., 2007; Gopar-Merino et al., 2015). Перечень использованных биоклиматических индексов с формулами их расчета и некоторыми примерами их применения в исследованиях связей растительности и климата приведен ниже.
1. Индекс континентальности – simple continentality index (Ic) [Ic = Tmax – Tmin, где Tmax – средняя многолетняя температура самого теплого месяца (июль) (°C), Tmin – средняя многолетняя температура самого холодного месяца (январь) (°C)]. Индекс характеризует различия в континентальности климата, используется для выделения биоклиматических секторов (Rivas-Martinez et al., 2004, 2011).
2. Индекс континентальности Бодье – Baudiere’s index (IcBaud) [IcBaud = (100 × × Psummer)/(${\text{M}}_{{{\text{sum}}}}^{2}$– ${\text{m}}_{{{\text{win}}}}^{2}$), где Psummer – среднее многолетнее количество осадков теплой четверти года (летние месяцы) (мм), Msum – средняя многолетняя максимальная температура самого теплого месяца (июль) (°C), mwin – средняя многолетняя минимальная температура самого холодного месяца (январь) (°C)]. Индекс используется при анализе высотных градиентов пространственной дифференциации видов и сообществ (Carballeira et al., 1983; Retuerto, Carballeira, 1990).
3. Индекс океаничности Кернера – Kerner oceanity index (OCE) [OCE = (T10 – T4)/(Tmax – Tmin), где T10 – средняя многолетняя температура октября, T4 – средняя многолетняя температура апреля (°C), Tmax – средняя многолетняя температура самого теплого месяца (июль) (°C), Tmin – средняя многолетняя температура самого холодного месяца (январь) (°C)]. Индекс предложен для оценки амплитуды температур на основе показателей разных сезонов года и применяется при климатической ординации как видов растений, так и растительных сообществ (Retuerto, Carballeira, 1992; Stonevicius et al., 2018).
4. Индекс термичности (термический индекс) – thermicity index (It) [It = T + mwin + Mwin, где T – средняя многолетняя температура (°C), mwin – средняя многолетняя минимальная температура самого холодного месяца (январь) (°C), Mwin – средняя многолетняя максимальная температура самого холодного месяца (январь) (°C)]. Индекс используется для выделения термических вариантов в пределах биоклиматических поясов (Rivas-Martinez et al., 1999, 2004).
5. Омбротермические индексы – ombrothermic indices. Ios – летний омбротермический индекс (июнь, июль, август) (ombrothermic index of summer) [Ios = (Psummer/Tsummer) × 10, где Psummer – среднее многолетнее количество осадков теплой четверти года (летние месяцы) (мм); Tsummer – средняя многолетняя температура теплой четверти года (летние месяцы) (°C)]. Показатель характеризует степень увлажнения при определенном уровне теплообеспеченности и используется при выделении омброклиматических типов в пределах биоклиматических поясов (Rivas-Martinez et al., 1999, 2004).
6. Показатель увлажненности – rain factor (Rf) [Rf = P/T, где P – среднее многолетнее количество осадков (мм), T – средняя многолетняя температура воздуха (°C)]. Показатель характеризует соотношение средних годовых условий тепло- и влагообеспеченности (Lang et al., 1976).
7. Плювиотермический показатель – pluviothermic quotient (Qpluv) [Qpluv = 2000× P/[(Tmax + 273.2)2 – – (Tmin + 273.2)2], где P – среднее многолетнее количество осадков (мм), Tmax – средняя многолетняя температура самого теплого месяца (июль) (°C), Tmin – средняя многолетняя температура самого холодного месяца (январь) (°C)]. Показатель предложен для оценки соотношения тепло- и влагообеспеченности (Emberger, 1930), являясь важным критерием разграничения условий развития высотно-поясных подразделений растительности, что определено на примере Средиземноморья (Gavilan, 2005).
Перечисленные 7 коэффициентов, основанные на отношении средних многолетних показателей тепло- и влагообеспеченности в целом за год, за теплый и холодный сезоны, а также на средних многолетних экстремальных значениях температур самого теплого и самого холодного месяцев (для Северного Забайкалья – июль и январь соответственно), предоставляют количественные данные о климатических условиях, активно используемых в исследованиях и важных для характеристики пространственной структуры растительного покрова на региональном уровне.
Для интегральной оценки пространственной структуры климатических условий выбранные 7 биоклиматических коэффициентов преобразованы по методу главных компонент, используемому в географических исследованиях для определения основных векторов, на которые приходится максимальное варьирование исходных переменных – факторов среды (Greig-Smith, 1983). Значения всех коэффициентов имеют нормальное либо близкое к нормальному распределение. Для каждой из трех первых компонент, независимо отражающих варьирование климатических условий, на основе расчета коэффициента корреляции Пирсона определены показатели, имеющие с ними наиболее тесную связь (летний омбротермический индекс, индекс континентальности Бодье, показатель увлажненности). Эти показатели, наряду со средними многолетними значениями тепло- и влагообеспеченности, использованы в качестве опорных данных при климатической ординации растительности и эколого-географической интерпретации пространственной структуры растительности оробиома.
Оценка влияния климатических условий на региональную специфику фитоценотического разнообразия оробиома проведена для фоновых типологических единиц географо-генетической классификации растительности. В анализе использованы такие биоклиматические показатели, с которыми выявлена наиболее тесная связь с главными компонентами: летний омбротермический индекс, индекс континентальности Бодье, показатель увлажненности, а также индекс континентальности (Rivas-Martinez et al., 2004) и средние многолетние значения средних и максимальных температур июля, минимальных температур января и количества осадков июля. С учетом нормального и близкого к нормальному распределению данных показателей по ним проведен дискриминантный анализ типологических подразделений растительности. Он позволил выявить среди них ключевые показатели, которые вносят наибольший вклад в разделение на группы (статистики Wilk’s lambda, Partial lambda, а также Standardizes coefficients для каждой дискриминантной функции). При сравнительной биоклиматической оценке типологических подразделений использован непараметрический критерий Краскела–Уоллеса. Сравнение проведено с учетом диаграмм размаха по наиболее значимым показателям, определенным в дискриминантном анализе. Климатическая ординация растительности с построением климатических ареалов (Grebenschikov, 1974; Nazimova, 1995; Parfenova, Chebakova, 2009; Gorichev, Davydychev, 2010) позволила дать оценку ее пространственной организации с учетом высотно-поясной структуры в системе важнейших характеристик тепло- и влагообеспеченности. Она представлена в двумерных системах координат, на осях которых отложены значения ключевых биоклиматических показателей. На схемах нашли отражение оптимальные для растительности климатические условия, определенные на основе стандартного отклонения от средних значений соответствующих ее подразделениям показателей.
Для проведения сравнительного биоклиматического анализа растительности Северовосточно-Забайкальского оробиома апробирован метод кластерного анализа ее типологических подразделений по средним значениям биоклиматических показателей, который нашел успешное применение при сравнительной биоклиматической характеристике растительности Северо-Восточной Азии (Nakamura et al., 2007). Всего использовано 34 типологические единицы в соответствии с разнообразием, отображенным на карте растительности юга Восточной Сибири. Каждая из них занимает определенный климатоп, которому может быть дана количественная оценка по важнейшим биоклиматическим характеристикам. К ним отнесены 10 переменных: индекс континентальности, летний омбротермический индекс, индекс континентальности Бодье, показатель увлажненности, средние многолетние годовая и июльская температуры, среднее многолетнее годовое и июльское количество осадков, средняя многолетняя максимальная температура июля, средняя многолетняя минимальная температура января. Использован метод Уорда (Ward, 1963), в качестве меры сходства – Евклидово расстояние. Дополнительно проведена кластеризация методом k-средних на 4 кластера, соответствующих обособившимся группам при кластеризации методом Уорда. Кластеризация послужила основой анализа пространственной структуры растительности на основе климатопов сообществ, характеризующих оптимальные климатические условия развития через средние значения биоклиматических показателей и их стандартные отклонения. Показатели, вносящие наибольший вклад в разделение кластеров, определены при дисперсионном анализе кластеров по значениям F-статистики и уровня статистической значимости (p-value). В осях наиболее значимых показателей построены двумерные ординационные схемы, на которых нашла отражение пространственная организация растительности в соответствии с высотно-поясными подразделениями.
Составление интегральной биоклиматической матрицы растительности и определение оптимальных условий развития ее подразделений проведено в соответствии с принципами построения биоклиматической матрицы, которая является одним из инструментов, служащих для оптимального отображения и сравнительного анализа разнообразия растительности в связи с климатическими условиями (Korrelyatzionnaya…, 1977). Средние значения ключевых показателей и стандартные отклонения от средних послужили ориентирами при определении биоклиматических оптимумов произрастания растительных сообществ и их высотно-поясных подразделений.
Латинские названия сосудистых растений даны по С.К. Черепанову (1995).
Все операции с растровыми и векторными данными цифровой климатической модели, картографическими материалами проведены с помощью программ ArcGis (v. 10.2.2) (инструменты приложения ArcToolbox) и SAGA GIS (v. 2.1.4). Статистический анализ выполнен с помощью тематических инструментов приложения ArcCatalog, а также программ SPSS (v. 11.5) и Statistica (v. 12.5.192.5). Оформление карт проведено в среде ArcGis.
Климатические предпосылки формирования фитоценотического разнообразия в Северном Забайкалье
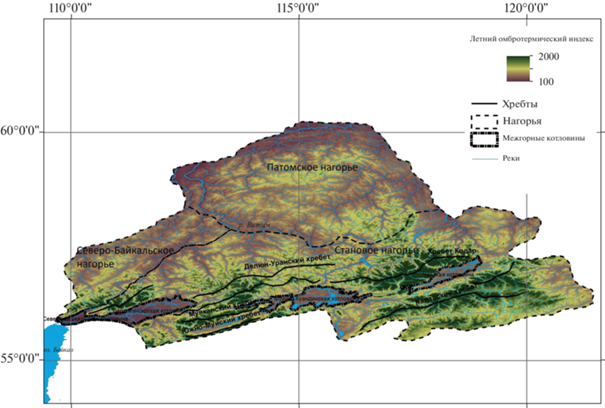
Северовосточно-Забайкальский оробиом связан с горными системами Северного Забайкалья – Становым, Северо-Байкальским и Патомским нагорьями с абсолютными высотами от 500–700 м н.у.м. у подножий межгорных котловин до 2500–3000 м н.у.м. на вершинах хребтов (рис. 1). Высотно-поясные закономерности в растительном покрове состоят в последовательной смене горнотаежного (500–900 м н.у.м.), подгольцового (900–1300 м н.у.м.), горнотундрового (1300–1900 м н.у.м.) и гольцового (1900–3000 м н.у.м.) поясов (Bocharnikov et al., 2018). В связи с выраженной высотной поясностью следует ожидать, что климатические предпосылки формирования структуры растительного покрова также тесно связаны с градиентом абсолютной высоты. При этом проявляются также широтные и долготные градиенты климатических показателей, которые прослеживаются, в общем виде, в направлениях с севера на юг и с запада на восток соответственно (Sochava, 1964; Hamet-Ahti, 1981; Tuhkanen, 1984; Krestov, Nakamura, 2007). Так, значения коэффициента континентальности Конрада (Conrad, 1946) увеличиваются примерно с 70 (юго-западная часть Северо-Байкальского нагорья) до 80 (хребты Калар, Удокан). Индекс континентальности, полученный на основе разницы между температурами самого теплого и самого холодного месяцев (Rivas-Martinez et al., 1999), варьирует в широких пределах (Krestov, Nakamura, 2007). Минимальные значения (около 30) приходятся на северо-восточное побережье Байкала, что сопоставимо со значениями индекса на юге Приморья и Сахалина и немного превышает минимальные значения для Азиатской части России значения на юге Курильских островов. Максимальные значения индекса континентальности (около 50) отмечаются на восточной периферии Станового и Патомского нагорий; области с большими значениями показателя связаны с ультраконтинентальным климатом Центральной Якутии.
Рис. 1.
Орографическая схема Северного Забайкалья.
Fig. 1. Orographic scheme of North Transbaikalia.

Распределение тепла и влаги в Северном Забайкалье связано с обширностью территории, градиентом абсолютной высоты, котловинным эффектом, проявляющимся в климатических различиях между межгорными котловинами Байкальской рифтовой зоны и окружающими их хребтами, воздействием озера Байкал, развитием долин крупных рек. Пространственная структура показателей согласуется с орографической структурой территории, маркируя разные высотные уровни горных хребтов и котловин, как, например, летний омбротермический индекс (рис. 2). Секторные различия, которые прослеживаются по вектору с запада на восток, выражаются через распределение показателя среднего многолетнего количества осадков января. Пространственная дифференциация климатических условий, рассматриваемая в целом для оробиома, находит отражение в климатическом районировании (Atlas…, 1967), определяя особенности структуры растительного покрова территории (Belov, 1973; Vladimirov et al., 2014).
Рис. 2.
Пространственное распределение значений летнего омбротермического индекса в Северном Забайкалье.
Fig. 2. Spatial distribution of ombrothermic index of summer in North Transbaikalia.
Пространственное распределение климатических условий на территории Северного Забайкалья
Интегральная оценка климатических условий, являющихся ключевыми в формировании структуры растительного покрова оробиома, на основе преобразования по методу главных компонент биоклиматических показателей характеризует различия в их вкладе в пространственную дифференциацию климата. На первые две компоненты, играющие наибольшую роль в соответствии со значениями собственных векторов (Eigenvalue), приходится 88% варьирования признаков, на первые три компоненты – 92%. Вклад климатических переменных в собственные вектора (Eigenvectors) оценен по значениям линейной корреляции. Наибольший вклад в первую компоненту (Eigenvalue – 55%) вносит летний омбротермический индекс (коэффициент корреляции Пирсона равен – 0.99, R2 = 0.9722), изменение значений которого тесно сопряжено с высотной дифференциацией орографической структуры (коэффициент корреляции Пирсона с абсолютной высотой равен 0.93, R2 = 0.8713) (рис. 2). Наибольшие значения средней многолетней температуры июля отмечаются в низкогорьях Патомского нагорья, котловинах Байкальской рифтовой зоны и в нижней части долины Витима (около +17…+18°С), закономерно понижаясь к среднегорьям и высокогорьям хребтов (до +3…+4°С). Это дает основания предположить, что соотношение осадков и температуры в летний период выступает в качестве одного из ключевых климатических критериев, обуславливающего высотно-поясную структуру растительного покрова оробиома в целом.
Вторая компонента тесно связана с показателем континентальности климата, определенной через индекс континентальности Бодье, который рассчитывается с учетом летней влагообеспеченности и экстремумов летних и зимних температур (коэффициент корреляции Пирсона равен 0.71, R2 = 0.5022) (рис. 3). В пространственной дифференциации данного показателя более выражены региональные различия. Четко обособляются межгорные котловины, для которых характерно наименьшее количество осадков в летний период (средние многолетние значения – около 70–90 мм в год). Среди них выделяются котловины восточной части Станового нагорья, имеющие максимальные значения индекса континентальности за счет снижения январского минимума температур (средние многолетние значения минимальной температуры января достигают ‒41…–43°С). Наветренные макросклоны наиболее высоких хребтов субширотного простирания характеризуются сниженной континентальностью климата.
Рис. 3.
Пространственное распределение значений индекса континентальности Бодье в Северном Забайкалье.
Fig. 3. Spatial distribution of Baudiere’s index in North Transbaikalia.

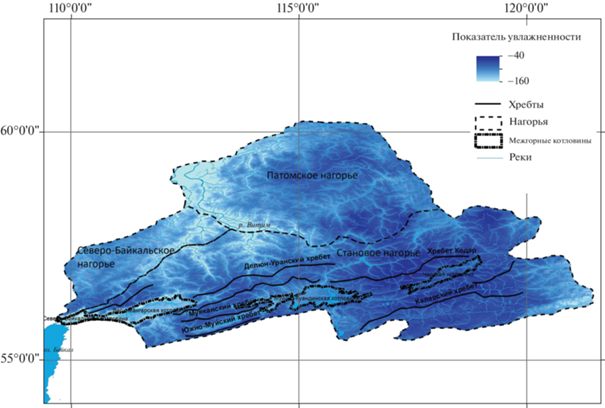
Третья компонента в наибольшей степени сопряжена с показателем увлажненности (rain factor), рассчитанным по отношению средних многолетних значений годовой суммы осадков и температур (коэффициент корреляции Пирсона равен –0.67, R2 = 0.4459) (рис. 4). Области с наименьшими значениями приходятся на Верхнеангарскую котловину, нижнюю часть бассейна Витима в пределах Патомского нагорья, область контакта хребта Удокан с Нюкжинским плато (юго-восточная часть Станового нагорья) с относительно невысокой влагообеспеченностью и повышенной теплообеспеченностью (средняя многолетняя температура достигает –3…–6°C). Наибольшие значения индекса отмечаются для хребтов Кодар, Калар с более высокой влагообеспеченностью и низкими температурами (среднее многолетнее количество осадков – 500 мм и выше).
Рис. 4.
Пространственное распределение значений показателя увлажненности в Северном Забайкалье.
Fig. 4. Spatial distribution of rain factor in North Transbaikalia.
Интегральный анализ биоклиматических показателей раскрывает ключевые пространственные закономерности климатических условий, с которыми связано формирование структуры растительного покрова оробиома. С высотным градиентом сопряжено изменение теплообеспеченности, что подтверждается высокой связью абсолютных высот как с температурными показателями, прежде всего, летнего периода, так и индексами, рассчитанными на их основе. Показатели наибольшей теплообеспеченности маркируют днища межгорных котловин, низкогорья хребтов, нижнюю часть долины реки Витим и его притоков в пределах Патомского нагорья.
Биоклиматические показатели, основанные на соотношении тепла и влаги, имеют меньшую связь с абсолютной высотой. На пространственную дифференциацию значений данных показателей оказывают влияние секторные закономерности, которые могут быть оценены через интегральное выражение региональной специфики растительного покрова. Абсолютная высота определяет общие тенденции в пространственной дифференциации степени континентальности климата, оцениваемой через индекс континентальности (IcBaud) и показатель увлажненности (Rf), причем пониженной континентальностью характеризуются высокогорья (Kolomytz, 1966).
Более существенные биоклиматические различия выражены в связи с развитием типов морфоструктур, спецификой орографического строения, выраженной через ориентацию горных хребтов в широтном и долготном отношении. На биоклиматических картосхемах (рис. 2–4) четко выделяются крупнейшие Верхнеангарская, Муйско-Куандинская, Чарская межгорные котловины. Разнообразие и пространственная структура их растительного покрова, выявляемая через абсолютные высоты, к которым приурочены котловины, и их геолого-геоморфологическую специфику (Aleksandrova, Preobrazhenskiy, 1964), может быть определена и климатическими условиями. Увеличение континентальности климата рассматривается в качестве одного из факторов, приводящих к уменьшению фитоценотического разнообразия и более равномерному его распределению по высотному градиенту в географических вариантах оробиома (Bocharnikov et al., 2018). Северные и южные макросклоны окружающих котловины хребтов, в наибольшей степени выраженные в связи с их субширотным простиранием на Становом нагорье, также дифференцированы в соответствии со степенью континентальности, что особенно характерно для крутосклонных, достигающих наибольших абсолютных высот хребтов.
Разнообразие фратрий классов растительных формаций Северовосточно-Забайкальского оробиома в связи с климатическими условиями
В растительном покрове Северного Забайкалья выражена трехчленная высотно-поясная структура. Высотный спектр сложен горнотаежным поясом лиственничных (Larix gmelinii) лесов, подгольцовым поясом березово-лиственничных (Betula lanata) редколесий и кедровостланиковых (Pinus pumila) сообществ, горнотундровым поясом, а также, на наиболее высоких хребтах, гольцовым поясом (Ogureeva, 1999; Bocharnikov et al., 2018). Исходя из опыта исследований по оценке влияния климата на растительность горных территорий (Polikarpov et al., 1986; Nazimova, 1995), смена высотных поясов и типологических подразделений растительности (типы растительности, классы и группы формаций), которые их характеризуют, хорошо согласуется с термическим градиентом, тогда как показатели влагообеспеченности определяют, преимущественно, характер высотных спектров, типы и подтипы высотной поясности, то есть различия в разнообразии поясов между высотно-поясными колонками. В целом, формирование структуры высотной поясности происходит под влиянием комплекса климатических условий, которые при оценке глобальных и региональных закономерностей разнообразия биоты являются главенствующими среди других факторов. Различия в высотном положении поясов в пределах одного типа поясности связаны, как правило, с особенностями перераспределения климатических условий в связи с орографической структурой в пределах конкретного хребта, например, оробореальной экспозиционной лесостепи (Ogureeva, 1980), пространственная структура сообществ которой обусловлена климатическими показателями (Hais et al., 2016). Дополнительную сложность в организацию растительного покрова и его разнообразие вносят литологические условия. Таким образом, пространственную структуру растительности в пределах высотных поясов необходимо рассматривать с учетом широкого спектра условий, воздействующих на топологическом уровне и определяющих разнообразие подразделений растительности более низкого ранга. В связи с этим базовой задачей является определение закономерностей структуры растительности и уровней организации биоты, роль климатических условий в которых является определяющей.
Растительность оробиома относится к пяти фратриям классов формаций. Они являются сложившимися в результате длительного исторического развития флороценогенетическими образованиями, современное разнообразие и совместное развитие которых на определенной территории определяет ее ботанико-географическую специфику.
С Восточносибирской фратрией горнотундровых формаций связано разнообразие высокогорий, прежде всего, горнотундрового и гольцового высотных поясов. В условиях гольцового типа высокогорий (Tolmachev, 1948) на высотах в среднем более 1300 м формируются кустарничковые (Dryas punctata, Cassiope ericoides), осоковые (Carex ensifolia), лишайниковые (Cladonia stellaris, C. arbuscula) тундры. С их участием связано фоновое разнообразие в структуре растительного покрова высокогорий Северного Забайкалья (Sofronov, 2015). Ограниченное распространение имеют сообщества Алтае-Тянь-Шанской фратрии альпийских формаций. Их наибольшее разнообразие приходится на альпийский тип высокогорий, характерный для гор Южной Сибири, где они широко представлены субальпийскими и альпийскими лугами (Sedel’nikov, 1988; Telyatnikov, 2015), субальпийскими темнохвойными и березовыми редколесьями (Shumilova, 1962; Zibzeev et al., 2015). На Становом нагорье ограниченное развитие получают альпинотипные и субальпинотипные луга на восточной границе своего распространения на высотах 900–1200 м н.у.м. Стланиковые сообщества Берингийской фратрии формаций являются фоновыми для подгольцового пояса (900–1300 м н.у.м.). Они представлены кустарниковыми сообществами с кедровым стлаником, редколесьями из березы шерстистой (Betula lanata), произрастающими в относительно широкой амплитуде экотопических условий в пределах, преимущественно, подгольцового пояса. Фитоценотическое разнообразие стланиковых сообществ в Северном Забайкалье уступает разнообразию вблизи центра их происхождения на Камчатке, сообщества только некоторых ассоциаций которых встречаются в рассматриваемом регионе (Neshataeva, 2011, Molozhnikov, 1986). С лиственничными (Larix gmelinii) лесами связано основное разнообразие Ангаридской фратрии. Наибольшее эколого-фитоценотическое разнообразие лиственничных лесов связано с горнотаежным поясом (600–900 м н.у.м.), фоновое участие которого выражено в пределах всего оробиома (Garaschenko, 1993; Vladimirov et al., 2014). Урало-Южносибирская фратрия представлена на восточном пределе распространения лишь небольшой частью своего разнообразия в нижней части горнотаежного пояса (500–600 м н.у.м.). Ее основное типологическое разнообразие и роль в структуре растительного покрова приходится на горы Южной Сибири (Sochava, 1980). Фоновое участие в сложении фитоценотического разнообразия Северовосточно-Забайкальского оробиома имеет растительность Байкало-Джугджурского регионального комплекса, включающего преобладающие в каждом высотном поясе сообщества, развитие которых связано с Северным Забайкальем (лиственничные леса и редколесья, сообщества кедрового стланика, высокогорные тундры).
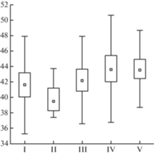
Дискриминантный анализ фратрий классов формаций для всего оробиома определил высокую значимость всех анализируемых биоклиматических переменных при p < 0.000 (табл. 1). Наибольший вклад в разделение фратрий вносят индекс континентальности, средняя многолетняя температура июля и показатель увлажненности. Каждая из выбранных переменных наилучшим образом характеризует климатические условия развития Ангаридской фратрии, фоновой для горнотаежного пояса, а также Восточносибирской фратрии, определяющей основное разнообразие высокогорий. Качество определения сообществ других фратрий значительно ниже (табл. 1). Диаграммы размаха значений трех показателей с наибольшим вкладом показывают соотношение оптимальных климатических условий развития растительности фратрий (табл. 2). Тест Краскела–Уоллиса показывает высокую достоверность их разделения в пространстве выбранных переменных (p = 0.000).
30. Ерниковые (Betula exilis) сфагновые болота.
31. Осоковые (Carex pseudocuraica, C. juncella, C. enervis) и вейниковые (Calamagrostis langsdorffii) луга.
32. Лиственничные травяные и аулакомниево-сфагновые мари.
Берингийская фратрия подгольцовых формаций.
33. Кедровостланиковые с лиственницей и ерниками (Betula divaricata, B. exilis) сообщества.
34. Березовые (Betula lanata) с кедровым стлаником и ольховником редколесья.
Fig. 5. Bioclimatic dendrogram of vegetation of the Northeast Transbaikal orobiome (cluster analysis based on the mean values of bioclimatic parameters for typological units of vegetation, Ward’s method, Euclidean distance).
Bioclimatic parameters: Ic – simple continentality index; It – thermicity index; Ios – ombrothermic index of summer; Rf – rain factor; Qpluv – pluviothermic quotient; OCE – Kerner’s oceanity index; IcBaud – Baudiere’s index; T_year – average annual temperature; T_july – average temperature of July; P_year – average annual precipitation; P_july – average precipitation of July; T_max_july – average maximum temperature of July; T_min_january – average minimum temperature of January.
Vegetation.
East Siberian phratry of montane tundra formations.
1. Shrub tundra (Betula rotundifolia, Rhododendron parvifolium, Salix glauca).
2. Dwarf shrub tundra (Cassiope ericoides, Empetrum nigrum, Salix sphenophylla) with crustose lichens.
3. Dwarf shrub (Ledum decumbens, Rhododendron aureum, Cassiope ericoides, Salix saxatilis, S. berberifolia) – moss-lichen tundra.
4. Meadow tundra (Festuca ovina, Diphasiastrum alpinum, Hierochloe alpina).
Altai-Tien Shan phratry of alpine formations.
5. Alpine (Trollius altaicus, Aquilegia glandulosa) and subalpine (Geranium albiflorum, Saussurea latifolia) meadows.
6. Kobresia (Kobresia myosuroides, K. simpliciuscula, Ptilagrostis mongholica) and fescue (Festuca airoides, F. ovina) meadows.
Ural-South Siberian phratry of montane taiga formations.
7. Fir-Siberian pine (Pinus sibirica, Abies sibirica) herb-bilberry-moss forests.
8. Fir-Siberian pine with Siberian dwarf pine (Pinus pumila) bergenia-bilberry-moss forests.
9. Pine (Pinus sylvestris) shrub herb forests.
10. Pine cowberry-bearberry lichen forests.
11. Spruce (Picea obovata) with Pinus pumila and Rhododendron aureum moss-lichen sparse forests.
12. Spruce-Siberian pine with fir and larch (Larix sibirica) herb-dwarf shrub (Vaccinium myrtillus, Ledum palustre) – moss forests.
13. Spruce-Siberian pine with Pinus pumila dwarf shrub-moss-lichen forests.
14. Spruce with fir, Siberian pine, larch and poplar (Populus suaveolens) shrub herb-moss forests.
15. Siberian pine-spruce herb-moss forests.
16. Larch and pine-larch herb forests.
17. Pine cowberry-bearberry-lichen forests.
18. Larch-pine with dark coniferous species cowberry-herb-moss forests.
Angarida phratry of montane taiga formations.
19. Larch (Larix gmelinii), with Siberian pine and spruce, alder (Duschekia fruticosa) herb-dwarf shrub-moss forests.
20. Larch with Pinus pumila, shrub birch and alder (Duschekia fruticosa) moss-lichen forests and sparse forests.
21. Larch with Pinus pumila dwarf shrub-moss forests and sparse forests.
22. Larch shrub birch (Betula divaricata, B. exilis) forests and sparse forests.
23. Larch with spruce shrub birch-alder (Duschekia fruticosa, Betula divaricata) dwarf shrub (Vaccinium vitis-idaea, Ledum palustre) – moss forests.
24. Larch with Siberian pine, fir, spruce, Pinus pumila and Rhododendron aureum herb-moss forests.
25. Larch dwarf shrub (Vaccinium uliginosum, Ledum palustre) – moss forests.
26. Larch with Rhododendron dauricum forests.
27. Larch-pine with alder and shrub birch (Betula divaricata, B. exilis) dwarf shrub-moss forests.
28. Larch-spruce and spruce-larch (Picea obovata) with Chosenia arbutifolia and poplar dwarf shrub-moss forests.
29. Shrub birch (Betula fruticosa) with larch and birch (Betula platyphylla) communities.
30. Shrub birch (Betula exilis) sphagnum bogs.
31. Sedge (Carex pseudocuraica, C. juncella, C. enervis) and small-reed (Calamagrostis langsdorffii) meadows.
32. Larch herb and Aulacomnium-Sphagnum mires. 33. Siberian dwarf pine with larch and shrub birch (Betula divaricata, B. exilis) communities.34. Birch (Betula lanata) with Pinus pumila and alder sparse forests.

Табл. 1.
Дискриминантный анализ сообществ фратрий классов растительных формаций Северовосточно-Забайкальского оробиома по биоклиматическим показателям Table 1. Discriminant analysis of vegetation communities of formation class phratries of Northeastern Transbaikal orobiome by bioclimatic parameters
| А | Фратрии классов формаций Phratries of formation classes |
||||||
|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | |||
| Верно классифицированные пиксели (%)/Correctly classified pixels (%) | 47.05 | 2.08 | 5.70 | 91.11k | 33.51 | ||
| Всего (%)/Total (%) | 66.95 | ||||||
| Б | N = 61203 Wilks’ Lambda: 0.58516; approx. F (32.225662) = 1102.9; p <0.000 |
||||||
| Биоклиматические показатели Bioclimatic parameters | Wilks’ Lambda | Partial Wilks’ Lambda | F-remove | p-value | |||
| Ic | 0.609930 | 0.959387 | 647.5870 | 0.000 | |||
| Ios | 0.596326 | 0.981273 | 291.9456 | 0.000 | |||
| Rf | 0.603266 | 0.969985 | 473.3752 | 0.000 | |||
| IcBaud | 0.592036 | 0.988383 | 179.8012 | 0.000 | |||
| T_july | 0.604745 | 0.967612 | 512.0417 | 0.000 | |||
| P_july | 0.598345 | 0.977961 | 344.7452 | 0.000 | |||
| T_max_july | 0.591318 | 0.989583 | 161.0360 | 0.000 | |||
| T_min_january | 0.597290 | 0.979689 | 317.1482 | 0.000 | |||
Примечание. А. Матрица классификации. Фратрии классов растительных формаций: I – Восточносибирская; II – Алтае-Тянь-Шанская; III – Берингийская; IV – Ангаридская; V – Урало-Южносибирская.
Б. Вклады переменных. Биоклиматические показатели: Ic – индекс континентальности; Ios – летний омбротермический индекс; Rf – показатель увлажненности; IcBaud – индекс континентальности Бодье; T_july – средняя многолетняя температура июля; P_july – среднее многолетнее количество осадков июля; T_max_july – средняя многолетняя максимальная температура июля; T_min_january – средняя многолетняя минимальная температура января.
Note. A. Classification matrix. Phratries of formation classes: I – East Siberian; II – Altai-Tien Shan; III – Beringian; IV – Angarid; V – Ural-South Siberian.
B. Variable contributions. Bioclimatic parameters: Ic – simple continentality index; Ios – ombrothermic index of summer; Rf – rain factor; IcBaud – Baudiere’s index; T_july – average temperature of July; P_july – average precipitation of July; T_max_july – average maximum temperature of July; T_min_january – average minimum temperature of January.
Таблица 2.
Диаграммы размаха значений ключевых биоклиматических показателей для фратрий классов формаций Table 2. Box plots of key bioclimatic indicators for phratries of formation classes
| Биоклиматические показатели Bioclimatic parameters |
Диаграммы размаха (Whisker box plots) (медианы,
квартили (25–75%), максимальные и минимальные значения) Whisker box plots (medians, quartiles (25–75%), maximum and minimum values) |
|---|---|
| Ic (индекс континентальности) Ic (simple continentality index) |
 |
| T_july (средняя многолетняя температура июля) T_july (average temperature of July) |
 |
| Rf (показатель увлажненности) Rf (rain factor) |
 |
Условия теплообеспеченности являются ключевыми показателями, определяющими развитие растительных сообществ фратрий, с фоновым участием сообществ которых связано формирование высотно-поясных подразделений оробиома (табл. 2). Индекс континентальности детерминирует биоклиматические различия растительности Ангаридской и Урало-Южносибирской фратрий, слагающей горнотаежный пояс, и Восточносибирской, Алтае-Тянь-Шанской и Берингийской фратрий, развитой в высокогорьях (подгольцовый, горнотундровый, гольцовый пояса). В условиях наименьшей континентальности климата развиваются сообщества Алтае-Тянь-Шанской фратрии, представленные формациями альпинотипных лугов, которые в высокогорьях Северного Забайкалья развиваются на северо-восточном пределе своего распространения (Tyulina, 1976; Bocharnikov et al., 2018). Соотношение тепла и влаги также играет важную роль в биоклиматической дифференциации фратрий. Наименьшие значения показателя увлажненности характерны для сообществ Урало-Южносибирской фратрии, развивающихся в условиях повышенного увлажнения (годовая сумма осадков – более 450 мм) при наиболее высоком уровне теплообеспеченности (средняя многолетняя температура –5°…–6°С). Показатель увлажненности определяет оптимальные в отношении соотношения тепла и влаги условия, с которыми связано развитие лиственнично-кедрово-пихтовых лесов в низкогорьях Патомского нагорья.
Биоклиматическая ординация растительности Северовосточно-Забайкальского оробиома и интегральная оценка пространственной организации ее разнообразия
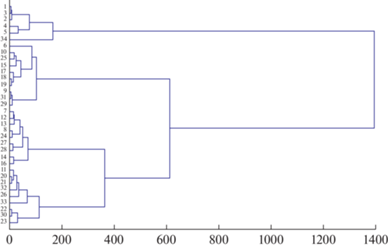
Анализ разнообразия растительности оробиома в условиях пространственной дифференциации климата как фактора его формирования проведен по 13 ключевым биоклиматическим показателям. Опорными единицами оценки послужили 34 типологические единицы растительности, связанные в пределах конкретного высотного пояса с определенным климатопом, оптимальные условия которого маркируются средними значениями биоклиматических показателей и стандартными отклонениями от них. Средние значения использованы при проведении кластерного анализа растительности. На построенной дендрограмме при близком уровне сходства выделяется 4 кластера, включающие в себя от 6 до 10 типологических подразделений растительности (рис. 5).
Рис. 5.
Биоклиматическая дендрограмма растительности Северовосточно-Забайкальского оробиома (кластерный анализ по средним значениям биоклиматических показателей для типологических единиц растительности, метод Уорда, мера сходства – Евклидово расстояние).
Растительность.
Восточносибирская фратрия горнотундровых формаций.
1. Кустарниковые (Betula rotundifolia, Rhododendron parvifolium, Salix glauca) тундры.
2. Кустарничковые (Cassiope ericoides, Empetrum nigrum, Salix sphenophylla) с господством накипных лишайников тундры.
3. Кустарничково (Ledum decumbens, Rhododendron aureum, Cassiope ericoides, Salix saxatilis, S. berberifolia) – мохово-лишайниковые тундры.
4. Олуговелые (Festuca ovina, Diphasiastrum alpinum, Hierochloe alpina) тундры.
Алтае-Тянь-Шанская фратрия альпийских формаций.
5. Альпинотипные (Trollius altaicus, Aquilegia glandulosa) и субальпинотипные (Geranium albiflorum, Saussurea latifolia) луга.
6. Кобрезиевые (Kobresia myosuroides, K. simpliciuscula, Ptilagrostis mongholica) и овсянициевые (Festuca airoides, F. ovina) луга.
Урало-Южносибирская фратрия горнотаежных формаций.
7. Пихтово-кедровые (Pinus sibirica, Abies sibirica) мелкотравно-чернично-зеленомошные леса.
8. Пихтово-кедровые кедровостланиковые (Pinus pumila) баданово-чернично-зеленомошные леса.
9. Сосновые (Pinus sylvestris) кустарниковые травяные леса.
10. Сосновые бруснично-толокнянковые лишайниковые леса.
11. Еловые (Picea obovata) с кедровым стлаником и золотистым рододендроном (Rhododendron aureum) мохово-лишайниковые редколесья.
12. Елово-кедровые с пихтой и лиственницей (Larix sibirica) мелкотравно-кустарничково (Vaccinium myrtillus, Ledum palustre) – зеленомошные леса.
13. Елово-кедровые кедровостланиковые кустарничково-мохово-лишайниковые леса.
14. Еловые с пихтой, кедром, лиственницей, тополем (Populus suaveolens) кустарниковые травяно-зеленомошные леса.
15. Кедрово-еловые мелкотравно-зеленомошные леса.
16. Лиственничные и сосново-лиственничные травяные леса.
17. Сосновые бруснично-толокнянково-лишайниковые леса.
18. Лиственнично-сосновые иногда с примесью темнохвойных пород бруснично-мелкотравно-зеленомошные леса.
Ангаридская фратрия горнотаежных формаций.
19. Лиственничные (Larix gmelinii) с кедром и елью ольховниковые (Duschekia fruticosa) мелкотравно-кустарничково-зеленомошные леса.
20. Лиственничные с кедровым стлаником, ерником и ольховником (Duschekia fruticosa) мохово-лишайниковые леса и редколесья.
21. Лиственничные с кедровым стлаником кустарничково-моховые леса и редколесья.
22. Лиственничные ерниковые (Betula divaricata, B. exilis) леса и редколесья.
23. Лиственничные с елью ерниково-ольховниковые (Duschekia fruticosa, Betula divaricata) кустарничково (Vaccinium vitis-idaea, Ledum palustre) – зеленомошные леса.
24. Лиственничные с кедром, пихтой, елью, с кедровым стлаником и золотистым рододендроном мелкотравно-моховые леса.
25. Лиственничные кустарничково (Vaccinium uliginosum, Ledum palustre) – моховые леса.
26. Лиственничные с рододендроном даурским (Rhododendron dauricum) леса.
27. Лиственнично-сосновые с ольховником и ерником (Betula divaricata, B. exilis) кустарничково-зеленомошные леса.
28. Лиственнично-еловые и елово-лиственничные (Picea obovata) с чозенией (Chosenia arbutifolia) и тополем кустарничково-зеленомошные леса.
29. Ерниковые (Betula fruticosa) с лиственницей и березой (Betula platyphylla) сообщества.
Кластер 1 включает в себя леса Ангаридской фратрии, представленные фоновыми для горнотаежного пояса сообществами. Основное разнообразие приходится на лиственничные (Larix gmelinii) кедровостланиковые (Pinus pumila), ольховниковые (Duschekia fruticosa), ерниковые (Betula divaricata, Betula exilis) багульниковые (Ledum palustre) и кустарничково (Vaccinium vitis-idaea) – зеленомошные леса. При большой занимаемой площади и высотной амплитуде (600–1200 м) они характеризуются близкими значениями для большинства биоклиматических показателей. При значениях индекса континентальности около 45 развиваются лиственничники в условиях наименьшей континентальности для области их распространения в Северо-Восточной Азии (Nakamura et al., 2007). Менее распространены лиственничные и ерниковые комплексы марей, развивающиеся в сходных климатопах, но имеющие эдафическую специфику формирования. В данный кластер также входят лиственничные редколесья и сообщества кедрового стланика, образующие подпояса подгольцового пояса. Для них характерно активное взаимопроникновение, в том числе, в соседние подпояса. Заросли кедрового стланика могут достигать низкогорий хребтов и днищ межгорных котловин, но оптимальные условия находят на высотах 1000–1400 м. Климатоп редколесных и стланиковых сообществ характеризуется более низкими средними годовыми (–7…–10°C) и июльскими (+12…+15°C) температурами воздуха при одинаковой влагообеспеченности. Уровень континентальности, в среднем, ниже по сравнению с горнотаежными лесами.
Кластер 2 включает в себя растительность высокогорий, относящихся к разным генетическим комплексам. Наиболее тесную связь по биоклиматическим показателям имеют высокогорные тундры, формирующие горнотундровый пояс на всех хребтах горной системы. Фоновую роль здесь играют дриадовые (Dryas punctata), овсяницевые (Festuca ovina, F. brachyphylla), мохово-лишайниковые (Cetraria islandica, C. cucullata, Alectoria ochroleuca) тундры, типичные для гольцовых систем Восточной Сибири. Оптимум их развития приходится на высоты 1200–1800 м н.у.м. и связан с низкой теплообеспеченностью, характеризуемой не столько средними многолетними годовыми показателями (например, значения индекса термичности около –68…–70 сопоставимы с аналогичными для сосново-лиственничных лесов котловин Станового нагорья), сколько c показателями вегетационного периода (средние многолетние температуры июля –9…–10°C). Сообщества южносибирского комплекса (альпинотипные и субальпинотипные луга) имеют максимальные для всего оробиома показатели плювиотермического коэффициента за счет сниженной разницы в амплитудах температур самого теплого и холодного месяцев и повышенного увлажнения (среднее годовое количество осадков – 500–550 мм). Они сосредоточены, преимущественно, в западной части Северо-Байкальского и Станового нагорий на высотах около 1500 м н.у.м., к востоку от них встречаясь только небольшими фрагментами, находя благоприятные экотопические условия, связанные с повышенной теплообеспеченностью. Наиболее обособленными в высокогорном кластере являются сообщества березовых (Betula lanata) редколесий. Они распространены в подгольцовом поясе наряду с лиственничными редколесьями и зарослями кедрового стланика. Климатоп сообществ характеризуется пониженными значениями показателя увлажненности, а по сравнению с горными тундрами, в среднем, более низкими средними годовыми (–10…–12°C) и июльскими (+8…+12°C) температурами воздуха.
Кластер 3 состоит из наиболее близких по биоклиматическим показателям подразделений. В его состав включены леса, преимущественно, нижней части горнотаежного пояса, типологически относящиеся к двум фратриям. Урало-Южносибирская фратрия представлена темнохвойными пихтово-кедровыми (Pinus sibirica, Abies sibirica) кустарничково (Vaccinium myrtillus, V. uliginosum) – зеленомошными (Pleurozium schreberi, Dicranum elongatum) лесами, занимающими небольшую площадь в низкогорьях Патомского и Северо-Байкальского нагорий (400–700 м н.у.м.). Еловые (Picea obovata) с участием темнохвойных видов и лиственницы (Larix sibirica, L. gmelinii) леса распространены по долинам крупных рек, в основном, на Патомском нагорье. Сообщества Ангаридской фратрии формаций, входящие в рассматриваемый кластер, имеют переходные черты к Урало-Южносибирским в отношении фитоценотического разнообразия и структуры. В составе их древостоя участвуют темнохвойные виды, а также сосна (Pinus sylvestris). Данные леса находят специфику и в своей географии в Северном Забайкалье. Лиственничные золотисторододендроновые (Rhododendron aureum) леса произрастают в среднегорьях (700–900 м н.у.м.) Северо-Байкальского нагорья при более высоком уровне влагообеспеченности в горнотаежном поясе (среднее годовое количество осадков – 450–500 мм). Лиственнично-еловые с чозенией (Chosenia arbutifolia) кустарничково-зеленомошные леса произрастают более широко, но только в нижних частях речных долин. Специфика кластера с биоклиматических позиций заключается в наименьшем среди всех кластеров среднем значении показателя увлажненности и максимальном – индекса термичности. Данные показатели характеризуют важные рубежи, ограничивающие развитие темнохвойных лесов оробиома. Для индекса термичности такой рубеж приходится на значение около –60, ниже которого темнохвойные леса вытесняются лиственничными. Данная региональная закономерность подтверждается сменой темнохвойных лесов на светлохвойные в связи с изменением годовой амплитуды температур, что выявлено для всей области распространения вечной мерзлоты в Сибири (Nazimova, 1995).
Кластер 4 в отношении типологического разнообразия наиболее разнородный, но в высотном распространении имеющий небольшую амплитуду. В него входят сообщества, с которыми связано формирование сложной пространственной структуры растительного покрова нижней части горнотаежного пояса, получающего развитие в низкогорьях хребтов и межгорных котловинах (400–700 м н.у.м.). Это сосновые кустарничковые (Vaccinium vitis-idaea), мелкотравно-зеленомошные (Rhytidium rugosum, Polytrichum juniperinum) и лишайниковые (Cladonia stellaris, C. ra.pngerina) леса, ерниковые (Betula fruticosa) сообщества, влажные осоковые (Carex pseudocuraica, C. enervis, C. cespitosa) и остепненные овсяницевые (Festuca ovina) луга. Они формируются в широкой амплитуде климатических условий, в частности, индекса термичности (от –52 до –69), средней годовой температуры (–3…–6°C) за счет участия в растительном покрове разных межгорных котловин, в значительной степени дифференцированных по биоклиматическим показателям (рис. 2–4). Пространственная дифференциация растительности, помимо климатических условий, определяется глубиной залегания вечной мерзлоты, составом и мощностью подстилающих горных пород, заболоченностью и другими факторами.
Интегральный анализ дендрограммы показывает, что классификация типологического разнообразия растительности, проведенная на основе биоклиматических показателей, отражает два ведущих комплексных градиента факторов, обуславливающих географию данного разнообразия. Градиентом первого порядка выступают высотно-поясные закономерности. С ними связано формирование смены высотных поясов растительности, находящих отражение в обобщенном виде в высотных спектрах растительности оробиома (Bocharnikov et al., 2018). Основное фитоценотическое разнообразие оробиома, фоновая роль которого подтверждается занимаемой типологическими единицами площадью, сопряжено с высотным градиентом. Градиент второго порядка характеризует провинциальные закономерности, которые находят свое выражение в пределах поясов в связи с участием их в разных типах, подтипах и географических вариантах типов поясности. С данными закономерностями связано развитие ограниченных в участии в растительном покрове сообществ, находящихся в условиях оробиома на географических рубежах своего распространения и пределе подходящих климатических условий.
Кластеризация методом K-средних определила высокую значимость выделенных групп сообществ по биоклиматическим показателям (табл. 3). Наибольший вклад вносят показатели среднего многолетнего количества осадков, летнего омбротермического индекса, средней многолетней температуры июля и плювиотермического показателя, оптимальные значения которых легли в основу климатической ординации кластеров (рис. 6). В двумерной системе координат в осях данных показателей занимаемые ими области, определенные средними и стандартными отклонениями от средних выбранных показателей, не пересекаются, маркируя биоклиматические градиенты, которыми обусловлена смена высотных поясов и подпоясов растительности оробиома (показатели тепло- и влагообеспеченности для всего года и вегетационного периода). Пространственная организация фоновых типологических подразделений растительности, образующих фитоценотическое разнообразие оробиома в соответствии с его высотно-поясной структурой растительности, нашла отражение на интегральной схеме биоклиматической ординации в осях наиболее значимых показателей (рис. 7). При максимальном количестве годовых осадков и наиболее низкой теплообеспеченности формируются гольцовый и горнотундровый пояса в верхней части высотного спектра. Область оптимального развития их сообществ приходится на значения летнего омбротермического индекса более 300. В пределах горнотундрового пояса при наибольшем количестве осадков и наименьших значениях омбротермического индекса развитие получают альпинотипные и субальпинотипные луга. Смена горнотундрового пояса на подгольцовый происходит в широкой амплитуде значений омбротермического индекса. Более четко данную смену детерминирует увлажнение (граничным является значение около 500 мм осадков в год). Активное взаимопроникновение в пределах высотного спектра сообществ кедрового стланика, березовых и лиственничных редколесий характеризует значительный интервал значений годового количества осадков (400–500 мм), в котором они произрастают. Граница между оптимальными условиями развития фоновых сообществ подпоясов подгольцового пояса определяется летним омбротермическим индексом (300–320). Преимущественно термические показатели определяют смену подпоясов горнотаежного пояса, которая происходит при значениях летнего омбротермического индекса 150–180. Уровень влагообеспеченности маркирует различия в типологическом разнообразии лесов в пределах нижнего подпояса горнотежного пояса. При годовом количестве осадков менее 400 мм формируются лиственнично-сосновые леса межгорных котловин Станового нагорья; для лиственнично-елово-пихтовых лесов низкогорий Патомского нагорья характерны значения 450–500 мм осадков в год. Их развитие происходит при близких значениях омбротермического индекса.
Табл. 3.
Биоклиматические показатели объединений типологических подразделений растительности Северовосточно-Забайкальского оробиома на основе их климатопов (значения средних, Mean, и стандартных отклонений, STD) Table 3. Bioclimatic parameters of typological units of vegetation of Northeastern Transbaikal orobiome by their climatiopes (values of Mean and STD – standard deviations)
| Биоклиматические показатели Bioclimatic parameters | Кластеры/Clusters | ||||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||||
| Статистика/Statistics | Типологические подразделения/Typological units | ||||||
| F | p-value | 11, 20, 21, 22, 23, 26, 30, 32, 33 | 1, 2, 3, 4, 5, 34 | 7, 8, 12, 13, 14, 16, 24, 27, 28 | 6, 9, 10, 15, 17, 18, 19, 25, 29, 31 | ||
| Ic | 8.0883 | 0.000428 | Mean | 43.7975 | 40.5889 | 43.9367 | 45.1465 |
| STD | 1.87879 | 1.58069 | 0.98748 | 2.35261 | |||
| It | 6.3104 | 0.001897 | Mean | –66.1739 | –68.6550 | –61.6057 | –62.8061 |
| STD | 3.32346 | 1.45741 | 1.96978 | 5.14905 | |||
| Ios | 89.1821 | 0.000000 | Mean | 209.9245 | 337.7616 | 151.2030 | 139.8534 |
| STD | 20.03914 | 48.33458 | 14.90073 | 17.01550 | |||
| Rf | 10.5201 | 0.000069 | Mean | –60.5588 | –51.7406 | –78.6397 | –71.1462 |
| STD | 5.90133 | 3.09485 | 7.12779 | 15.88916 | |||
| Qpluv | 60.4948 | 0.000000 | Mean | 38.4126 | 47.4892 | 38.1410 | 31.5184 |
| STD | 2.99964 | 2.84130 | 1.90891 | 1.39439 | |||
| OCE | 11.1630 | 0.000044 | Mean | –0.0224 | 2.2490 | –1.4339 | –1.6717 |
| STD | 0.99041 | 0.98708 | 0.77946 | 2.21607 | |||
| IcBaud | 7.1664 | 0.000910 | Mean | –28.4923 | –30.4761 | –27.0864 | –25.2152 |
| STD | 3.28233 | 1.26013 | 2.02038 | 1.96436 | |||
| T_year | 52.8132 | 0.000000 | Mean | –7.5676 | –9.9250 | –5.7881 | –5.5858 |
| STD | 0.79514 | 0.59162 | 0.54170 | 0.90500 | |||
| P_year | 100.4268 | 0.000000 | Mean | 443.5320 | 505.7146 | 446.7471 | 378.6479 |
| STD | 20.37161 | 12.39180 | 12.98788 | 9.97654 | |||
| T_july | 85.6152 | 0.000000 | Mean | 14.4948 | 11.2219 | 16.0278 | 16.5356 |
| STD | 0.75286 | 1.01715 | 0.57272 | 0.44493 | |||
| P_july | 43.7506 | 0.000000 | Mean | 94.5836 | 108.0280 | 74.2212 | 75.3132 |
| STD | 3.76503 | 3.41427 | 6.73349 | 9.59734 | |||
| T_max_july | 22.6563 | 0.000000 | Mean | 20.0633 | 16.9394 | 20.5373 | 21.5170 |
| STD | 1.16493 | 1.10867 | 0.62033 | 1.33267 | |||
| T_min_jan | 1.2169 | 0.320621 | Mean | –36.1938 | –35.2910 | –34.6215 | –35.7618 |
| STD | 1.79346 | 1.13811 | 1.17142 | 2.52244 | |||
Рис. 6.
Климатические оптимумы объединений типологических единиц (кластеров) Северовосточно-Забайкальского оробиома по осям: а – летнего омбротермического индекса (Ios) и среднего многолетнего количества осадков (P); б – плювиотермического коэффициента (Qpluv) и средней многолетней температуры июля (T_july).
Fig. 6. Climatic optima of typological units (clusters) of Northeastern Transbaikal orobiome by axes: a – ombrothermic index of summer (Ios) and average annual precipitation (P); b – pluviothermic quotient (Qpluv) and average temperature of July (T_july).

Рис. 7.
Обобщенная биоклиматическая схема фитоценотического разнообразия Северовосточно-Забайкальского оробиома.
Ось x – летний омбротермический индекс (Ios); ось y – годовое количество осадков (P). Границы высотно-поясных подразделений: а – высотные подпояса; б – высотные пояса.
Растительность.
Гольцовый пояс: 1 – разреженные группировки накипных лишайников на каменистых россыпях.
Горнотундровый пояс: 2 – кустарничковые и мохово-лишайниковые тундры; 3 – альпинотипные и субальпинотипные луга.
Подгольцовый пояс: 4 – кедровостланиковые (Pinus pumila) сообщества; 5 – березовые (Betula lanata) и лиственничные (Larix gmelinii) редколесья.
Горнотаежный пояс: 6 – лиственничные (Larix gmelinii) леса; 7 – лиственнично-сосновые (Pinus sylvestris, Larix sibirica, L. gmelinii) леса; 8 – сосновые леса; 9 – пихтово-кедрово-еловые (Picea obovata, Pinus sibirica, Abies sibirica) леса.
Fig. 7. General bioclimatic scheme of phytocoenotic diversity of Northeastern Transbaikal orobiome.
X-axis – ombrothermic index of summer (Ios); Y-axis – average annual precipitation (P). The borders of altitudinal subdivisions: a – sub-belts; б –belts.
Vegetation.
Upper-tundra belt: 1 – sparse crustose lichen communities on rocky sites.
Montane tundra belt: 2 – dwarf shrub and moss-lichen tundra; 3 – alpine and subalpine meadows.
Sub-tundra belt: 4 – Siberian dwarf pine (Pinus pumila) communities; 5 – birch (Betula lanata) and larch (Larix gmelinii) sparse forests.
Montane taiga belt: 6 – larch (Larix gmelinii) forests; 7 – larch-pine (Pinus sylvestris, Larix sibirica, L. gmelinii) forests; 8 – pine forests; 9 – fir-Siberian pine-spruce (Picea obovata, Pinus sibirica, Abies sibirica) forests.
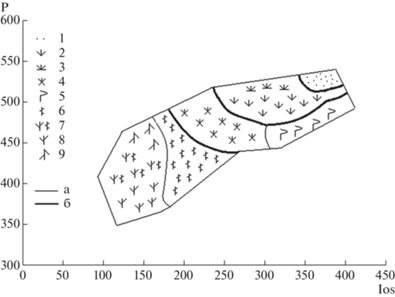
Выявленные климатические оптимумы фоновых для Северовосточно-Забайкальского оробиома типологических подразделений растительности послужили основой для построения интегральной биоклиматической матрицы в соответствии с принципами использования биоклиматических показателей в анализе пространственной структуры растительности (Rivas-Martinez et al., 2011, 2016). Показатели, взятые в его основу (прежде всего, индекс континентальности, индекс термичности, летний омбротермический индекс, средние многолетние температуры года и июля), использованы при биоклиматической классификации растительности оробиома. Фитоценотическое разнообразие растительного покрова связано, главным образом, с двумя группами биоклиматических показателей, характеризующих условия тепло- и влагообеспеченности, в соответствии с которыми все биоклиматическое пространство поделено на сектора. Уровень континентальности определяется значениями индекса континентальности и летнего омбротермического индекса. Уровень теплообеспеченности оценен по индексу термичности, средним многолетним годовым и июльским температурам. Выявленное соответствие данных показателей разнообразию растительности представлено в виде обобщенной биоклиматической матрицы (табл. 4). Ячейки в данной матрице отражают связь подразделений растительности с конкретными климатопами согласно высотно-поясной структуре растительности оробиома. В пределах секторов континентальности дифференциация растительности происходит по изменению характерных для нее значений теплообеспеченности.
Табл. 4.
Интегральная биоклиматическая матрица растительности Северовосточно-Забайкальского оробиома Table 4. Integral bioclimatic matrix of vegetation of Northeastern Transbaikal orobiome
| Макробиоклимат Macrobioclimate | Высотно-поясные подразделения Altitudinal subdivisions | Уровень теплообеспеченности Temperature supply levels | Сектора континентальности Continentality sectors | ||||
|---|---|---|---|---|---|---|---|
| ЭК | РК | К | |||||
| Пояса Belts | Подпояса Sub-belts | Сектора Sectors | Показатели (°C) Parameters (°C) | Ic = 46–50 | Ic = 40–46 | Ic = 36–40 | |
| Высокогорный гольцовый High mountain upper-tundra | I–II | Очень холодный Very cold | It = –75…–64 | 1 | 4 | ||
| T_year = –12…–8 | 2 | 5 | |||||
| T_july = +7… +14 | 3 | ||||||
| III | III.1 | Холодный Cold | It = –68…–62 | 33 | |||
| T_year = –10…–7 | |||||||
| T_july = +11… +15 | |||||||
| III.2 | It = –70…–62 | 11 | 34 | ||||
| T_year = –12…–7 | |||||||
| T_july = +7… +15 | |||||||
| Бореальный Boreal | IV | IV.1 | It = –74…–57 | 30 | 20 21 | ||
| T_year = –9…–5 | 32 | 22 23 | |||||
| T_july = +14… +17 | 25 26 | ||||||
| IV.2 | Умеренно-холодный Temperate cold | It = –64…–49 | 31 | 29 28 10 | 6 | ||
| T_year = –7…–3 | 9 | 17 27 24 | |||||
| T_july = +16… +18 | 19 13 8 | ||||||
| 14 12 15 | |||||||
| 18 16 7 | |||||||
Примечание. Типологические подразделения растительности и биоклиматические переменные – см. рис. 5.
Высотные пояса растительности: I – гольцовый; II – горнотундровый; III – подгольцовый (III.1 – кедровостланиковый подпояс; III.2 – редколесный подпояс); IV – горнотаежный (IV.1 – подпояс лиственничных лесов; IV.2 – подпояс лиственнично-сосновых и пихтово-кедрово-еловых лесов).
Сектора континентальности: ЭК – экстраконтинентальный; РК – резкоконтинентальный; К – континентальный.
Note. See Fig. 5 for typological subdivisions of vegetation and bioclimatic parameters.
Altitudinal belts of vegetation: I – upper-tundra; II – montane tundra; III – sub-tundra (III.1 – sub-belt of Siberian dwarf pine communities; III.2 – sub-belt of sparse forests); IV – montane taiga (IV.1 – sub-belt of larch forests; IV.2 – sub-belt of larch-pine and fir-Siberian pine-spruce forests).
Continentality sectors: ЭК – extracontinental; РК – strong continental; К – continental.
Растительность оробиома, составляющая основу его фитоценотического разнообразия, формируется в условиях резкоконтинентального климата. В интервале значений индекса континентальности от 40 до 46 развита растительность всех высотных поясов. Дифференциация между поясами маркируется разными показателями теплообеспеченности при выраженных пересечениях на осях с их значениями. В соответствии с этим выделено три сектора теплообеспеченности. В очень холодном секторе при наиболее низких средних годовых и июльских температурах формируется растительность горнотундрового пояса. С увеличением средних показателей теплообеспеченности происходит смена на подгольцовый и горнотаежный пояса, которые отнесены к холодному сектору. Нижний подпояс горнотаежного пояса формируется в условиях максимальных для оробиома температур и относится к умеренно-холодному сектору. С ним связано развитие наибольшего числа типологических подразделений растительности, которые формируются в узкой амплитуде абсолютных высот и занимают небольшую площадь в нижних частях гор и днищах межгорных котловин.
Региональные различия и географические связи оробиома в системе биомного разнообразия, выраженные в его фитоценотическом разнообразии, находят отражение в биоклиматической матрице через занимаемые подразделениями растительности оптимумы по показателю степени континентальности климата. В горнотундровом поясе снижение континентальности климата в высокогорьях западной части области развития оробиома приводит к участию в растительном покрове альпинотипных и субальпинотипных лугов, климатопы которых связаны с континентальным сектором (Ic = 36–40). Увеличение увлажнения и снижение амплитуды годовых температур позволяют им произрастать на границе области своего распространения, связанной с горами Южной Сибири, где они встречаются повсеместно в условиях ослабленной континентальности климата при годовом количестве осадков более 1000 мм (Polikarpov et al., 1986). Условия меньшей континентальности климата способствуют развитию березовых редколесий в подгольцовом поясе, приуроченных к наиболее влагообеспеченным склонам хребтов.
С межгорными котловинами связаны наиболее континентальные условия на Становом нагорье (экстраконтинентальный сектор, Ic = 46–50). Слабодренированные поверхности днища Верхнеангарской котловины заняты лугово-болотной растительностью с преобладанием мезо-гигрофильных злаково-осоковых лугов. По террасам Верхней Ангары и нижним частям световых склонов окружающих ее хребтов распространены сосновые кустарниковые остепненные леса. Здесь они развиваются при большем увлажнении и меньших термических ресурсах по сравнению с сосновыми лесами юга Забайкалья, где они формируют пространственные сочетания со степями в составе лесостепного пояса (Namzalov, 1998). В Северовосточно-Забайкальском оробиоме с практически полным отсутствием степей сосновые леса образуют узкий по высотной амплитуде подпояс и характеризуют наиболее континентальные условия развития подтаежных лесов Урало-Южносибирской фратрии на контакте с лиственничными лесами (Drobushevskaya, Tzaregorodtzev, 2007). В восточной части Станового нагорья по днищам котловин встречаются лиственничные и ерниковые комплексы марей. Для оробиома они являются наиболее западными фрагментами области их распространения, основное экосистемное разнообразие которых сосредоточено восточнее на пологонаклонных склонах нагорий и равнинах юга Восточной Сибири и Дальнего Востока (Prozorov, 1961).
Таким образом, в пространственной организации растительного покрова в Северном Забайкалье можно проследить три ключевых градиента, связанных с распределением климатических условий. Фитоценотическое разнообразие Северовосточно-Забайкальского оробиома формируется в условиях повышенной континентальности климата, относительно небольшого годового количества осадков и низкой теплообеспеченности при выраженном высотном градиенте данных биоклиматических показателей, в соответствии с которым развиваются фоновые для оробиома растительные формации. Смена высотных поясов определяется комплексом биоклиматических показателей, характеризующих соотношение тепла и влаги. В пределах поясов дифференциация на подпояса связана, преимущественно, с термическими показателями. Региональная специфика типологического разнообразия, проявляющаяся в разных поясах и вносящая изменения в их разнообразие, маркируется, прежде всего, условиями увлажнения. Подобные биоклиматические закономерности выявлены в структуре растительного покрова гор Южной Сибири (Polikarpov et al., 1986; Nazimova et al., 2005).
Специфика фитоценотического разнообразия оробиома, его пространственной структуры в связи с климатическими условиями прослеживается при сравнении с условиями формирования растительного покрова прилегающих гор Южной Сибири и юга Забайкалья. Высокогорья представлены разнообразием сообществ гольцового типа высокогорий в широком интервале показателей тепло- и влагообеспеченности, высотный градиент которых обуславливает смены растительности гольцового, горнотундрового и подгольцового поясов. С увеличением количества осадков в высокогорьях (более 500 мм в год) при средних показателях теплообеспеченности, характерных для сообществ горнотундрового пояса (табл. 3), формируется разнообразие высокогорий альпийского типа, небольшие фрагменты сообществ которого (альпинотипные и субальпинотипные луга) представлены в западной части области развития оробиома. Их основное разнообразие (альпийские и субальпийские луга и редколесья) приходится на горы Южной Сибири, где они участвуют в сложении растительного покрова высокогорий альпийского типа при значительно большем увлажнении (среднее годовое количество осадков превышает 1000 мм) и более высокой температуре (средняя годовая температура выше –5°С). Фоновое участие в горнотаежном поясе имеют лиственничные (Larix gmelinii) леса. В Северовосточно-Забайкальском оробиоме их климатоп характеризуется относительно высокими значениями годового количества осадков и низкой теплообеспеченностью (табл. 4) по сравнению с требованиями к увлажнению горнотаежных лиственничных лесов из Larix sibirica, распространенными в горах Южной Сибири и на юго-западе Забайкалья (Nazimova et al., 2004). Сосновые леса в оробиоме соответствуют наиболее континентальным и теплообеспеченным условиями, что связывает их с лиственнично-сосновой подтайгой семигумидного климатического сектора, разнообразие которого формируется при годовом количестве осадков менее 400 мм (Drobushevskaya, Nazimova, 2006). Темнохвойные леса играют ключевую роль только в нижнем подпоясе горнотаежного пояса в низкогорьях Патомского нагорья на контакте с темнохвойными лесами среднесибирского комплекса, где они находят для себя благоприятные условия со сниженной континентальностью климата при относительно большом количестве осадков (более 450 мм в год).
ЗАКЛЮЧЕНИЕ
Пространственная дифференциация биоклиматических показателей в Северном Забайкалье находит наиболее тесные связи с высотным градиентом и прослеживается в соответствии с секторными различиями, определяемыми степенью континентальности климата. Наибольшая амплитуда показателей тепло- и влагообеспеченности связана с абсолютной высотой и развитием крупных морфоструктур (горные хребты, межгорные котловины). Дополнительный вклад в высотно-поясную структуру растительности вносит пространственная дифференциация климатических условий, выраженная в соответствии с изменением географической долготы, что маркируется степенью увлажнения и биоклиматическими показателями, связанными с осадками. Изменение данных показателей не достигает такой тесной связи по сравнению с высотно-поясной структурой, однако вносит вклад в региональную специфику растительного покрова, прежде всего, в разнообразие формирующихся подтипов и вариантов типов поясности растительности (Ogureeva, 1999). Выявленное в пределах характерных для Северовосточно-Забайкальского оробиома типов поясности растительности фитоценотическое разнообразие оценено с учетом комплекса биоклиматических показателей на региональном уровне его организации. Среди них средние многолетние значения тепло- и влагообеспеченности теплого периода года, максимальных и минимальных температур самого теплого и самого холодного месяцев соответственно, годовой температуры и осадков, а также рассчитанные на их основе биоклиматические индексы выступили в качестве информативных показателей.
Северовосточно-Забайкальский оробиом как региональное подразделение в системе экосистемного разнообразия гор характеризуется единством в отношении высотно-поясной структуры растительности и определяющих ее градиентов климатических условий. Для оробиома свойственна трехчленная структура поясности, формирующаяся в резкоконтинентальном климатическом секторе при выраженном температурном градиенте. Различия в высотных пределах поясов и составе их фитоценотического разнообразия определяют региональную специфику, проявляющуюся в растительном покрове Северного Забайкалья. Однако фоновые черты, которые заключаются в повсеместном развитии горных тундр, стланиковых сообществ в рамках гольцового типа высокогорий (выше 900–1000 м н.у.м.), господстве лиственничных лесов в горнотаежном поясе (600–900 м н.у.м.), участии сосновых лесов в нижней части высотного спектра (400–600 м н.у.м.), характеризуют оробиом в целом.
Типологические подразделения растительности высшего иерархического уровня (фратрии классов формаций) имеют определенные климатические ареалы, характеризующие современные условия развития их разнообразия в оробиоме на основе ключевых биоклиматических показателей, которые маркируют их в ординационном климатическом пространстве. Смена этих подразделений на высотном спектре может быть выявлена на основе комплекса показателей тепло- и влагообеспеченности.
В целом, фоновые типологические подразделения растительности Северовосточно-Забайкальского оробиома развиваются в условиях континентального климата (с оптимумами развития при значениях индекса континентальности от 36 до 50) при градиенте теплообеспеченности (средняя многолетняя температура июля – от +7 до +18°C). Переход от фоновых горнотаежных формаций на высокогорные надежно детерминируется индексом континентальности (значения около 43), показателем увлажненности (значения около –60) и средней многолетней температурой июля (значения около +14°C).
Смена высотных поясов растительности оробиома маркируется изменением комплексных биоклиматических показателей, основанных на соотношении тепла и влаги. Показатели теплообеспеченности, прежде всего, вегетационного периода, являются ключевыми в пространственной дифференциации высотных подпоясов (смена редколесных и стланиковых сообществ в подгольцовом поясе, лиственничных и лиственнично-сосновых и темнохвойных лесов в горнотаежном поясе). Региональные различия в структуре высотной поясности оробиома и типологическом разнообразии поясов, проявляющиеся в разных частях горной системы, маркируются значениями влагообеспеченности. С ними связано развитие более требовательных к увлажнению пихтово-еловых лесов низкогорий Патомского нагорья, развивающихся при среднем многолетнем количестве осадков более 450 мм в год, и сосновых лесов межгорных котловин Станового нагорья (экстраконтинентальные условия с большой годовой амплитудой температур и небольшим количеством осадков – до 400 мм в год).
Список литературы
[Aleksandrova, Preobrazhenskiy] Александрова Т.Д., Преображенский В.С. 1964. Ландшафты малых котловин горной тайги. М. 88 с.
[Atlas] Атлас Забайкалья. Бурятская АССР и Читинская область. 1967. Иркутск. 176 с.
[Belov] Белов А.В. 1973. Карта растительности юга Восточной Сибири. Принципы и методы картографирования. – Геоботаническое картографирование. С. 16–30.
Bocharnikov M.V., Ogureeva G.N., Luvsandorj J. 2018. Regional features of the altitudinal gradients in northern transbaikalia vegetation cover. – Geography, environment, sustainability. 11 (4): 67–84. https://doi.org/10.24057/2071-9388-2018-11-4-67-84
Box E.O. 1995. Factors determining distributions of tree species and plant functional types. – Vegetatio. 121: 101–116. https://doi.org/10.1007/978-94-011-0343-5_10
[Buks] Букс И.И. 1976. Методика составления и краткий анализ корреляционной эколого-фитоценологической карты Азиатской России. М 1 : 7500000. – Геоботаническое картографирование. С. 44–51.
Carballeira A., Devesa C., Retuerto R., Santillan E., Ucieda F. 1983. Bioclimatologia de Galicia. Fund Petro Barrie La Coruna, Spain. P. 1–392.
[Czerepanov] Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб. 992 с.
Conrad V. 1946. Usual formulas of continentality and their limits of validity. – Transactions of the American Geophysical Union. 27 (5): 663–664.
[Drobushevskaya, Nazimova] Дробушевская О.В., Назимова Д.И. 2006. Климатические варианты светлохвойной низкогорной подтайги Южной Сибири. – География и природные ресурсы. 2: 21–27.
[Drobushevskaya, Tzaregorodtzev] Дробушевская О.В., Царегородцев В.Г. 2007. Географо-климатические варианты светлохвойных травяных лесов Сибири. – Сибирский экологический журнал. 2: 211–219.
Emberger L. 1930. Sur une formule climatique et ses applications en botanique. CR Hebd Acad Sci. Paris. 191: 389–391.
Fick S.E., Hijmans R.J. 2017. WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. – International Journal of Climatology. 37: 4302–4315. https://doi.org/10.1002/joc.5086
[Garaschenko] Гаращенко А.В. 1993. Флора и растительность Верхнечарской котловины (Северное Забайкалье). Новосибирск. 280 с.
Gavilan R.G. 2005. The use of climatic parameters and indices in vegetation distribution. A case study in the Spanish Sistema Central. – Int. J. Biometeorol. 50: 111–120. https://doi.org/10.1007/s00484-005-0271-5
[Golubyatnikov, Denisenko] Голубятников Л.Л., Денисенко Е.А. 2004. Влияние климатических изменений на продуктивность растительного покрова Европейской России. – Использование и охрана природных ресурсов в России. 1: 78–85.
Gopar-Merino L.F., Velazquez A., Gimenez de Azcarate J. 2015. Bioclimatic mapping as a new method to assess effects of climatic change. – Ecosphere. 6 (1): 1–12. https://doi.org/10.1890/ES14-00138.1
[Gorichev, Davydychev] Горичев Ю.П., Давыдычев А.Н. 2010. Зональные лесные формации Южного Урала: географическое распространение и положение в пространстве климатических факторов. – Известия Самарского научного центра РАН. 12 (1–3): 867–869.
Greig-Smith P. 1983. Quantitative Plant Ecology / 3rd ed. Berkeley. 359 p.
[Grebenschikov] Гребенщиков О.С. 1974. Опыт климатической характеристики основных растительных формаций Кавказа. – Бот. журн. 59 (2): 161–173.
Hais M., Chytry M., Horsak M. 2016. Exposure-related forest-steppe: A diverse landscape type determined by topography and climate. – J. Arid Environ. 135: 75–84. https://doi.org/10.1016/j.jaridenv.2016.08.011
Hamet-Ahti L. 1981. The boreal zone and its biotic subdivision. – Fennia. 159 (1): 69–75.
Hijmans R.J., Cameron S.E., Parra J.L., Jones P.G., Jarvis A. 2005. Very high resolution interpolated climate surfaces for global land areas. – International Journal of Climatology. 25 (15): 1965–1978. https://doi.org/10.1002/joc.1276
[Kolomytz] Коломыц Э.Г. 1966. Снежный покров горнотаежных ландшафтов Северного Забайкалья. М.-Л. 184 с.
[Korrelyatsionnaya…] Корреляционная эколого-фитоценотическая карта Азиатской части России. М 1 : 7500000. И.И. Букс, В.Н. Байбородин, Л.С. Тимирбаева. 1977. Иркутск.
Krestov P.V. 2007. Climatic Control of Zonal Vegetation in Northeast Asia with Special Accent to the Continentality-Oceanicity Gradient. – In: Vegetation mapping Series: Proceedings of the Fifth Symposium of Vegetation Diversity in Taiwan. Taipei. 9: 175–178.
Lang G.E., Reiners W.A., Heier R.K. 1976. Potential alternation of precipitation chemistry by epiphytic lichens. – Oecologia. 25 (3): 229–241. https://doi.org/10.1007/BF00345100
Major J. 1963. A climatic index to vascular plant activity. – Ecology. 44: 485–498. https://doi.org/10.2307/1932527
[Map “Biomes of Russia”] Карта “Биомы России” в серии карт природы для высшей школы (М 1 : 7500000). Г.Н. Огуреева, Н.Б. Леонова, Л.Г. Емельянова и др. 2018. М.
[Molozhnikov] Моложников В.Н. 1986. Растительные сообщества Прибайкалья. Новосибирск. 272 с.
Mucina L. 2019. Biome: evolution of a crucial ecological and biogeographical concept. – New Phytologist. 222: 97–114. https://doi.org/10.1111/nph.15609
Nakamura Y., Krestov P.V. 2007. Biogeographical diversity of alpine vegetation in the oceanic regions of Northeast Asia. – In: Berichte der REINHOLD-TUXEN-GESELLSCHAFT. 19: 117–129.
Nakamura Y., Krestov P.V., Omelko A.M. 2007. Bioclimate and vegetation complexes in Northeast Asia: a first approximation to integrated study. – Phytocoenologia. 37 (3–4): 443–470. https://doi.org/10.1127/0340-269X/2007/0037-0443
[Namzalov] Намзалов Б.Б. 1998. О некоторых аспектах географии растительности Бурятии. – В кн.: Исследования флоры и растительности Забайкалья. Улан-Удэ. С. 72–76.
[Nazimova] Назимова Д.И. 1995. Климатическая ординация лесных экосистем как основа их классификации. – Лесоведение. 4: 63–73.
[Nazimova et al.] Назимова Д.И., Ермаков Н.Б., Андреева Н.М., Степанов Н.В. 2004. Концептуальная модель структурного биоразнообразия зональных классов лесных экосистем Северной Евразии. – Сибирский экологический журнал. 11 (5): 745–756.
[Nazimova et al.] Назимова Д.И., Пономарев Е.И., Степанов Н.В., Федотова Е.В. 2005. Черневые темнохвойные леса на юге Красноярского края и проблемы их обзорного картографирования. – Лесоведение. 1: 12–18.
[Neshataeva] Нешатаева В.Ю. 2011. Сообщества кедрового стланика (Pinus pumila (Pall.) Regel) полуострова Камчатка. – Растительность России. 19: 71–100.
[Ogureeva] Огуреева Г.Н. 1980. Ботаническая география Алтая. М. 192 с.
[Ogureeva] Огуреева Г.Н. 1999. Карта “Зоны и типы поясности растительности России и сопредельных территорий” (М 1 : 7500000). Серия карт природы для высшей школы. М.
[Ogureeva, Bocharnikov] Огуреева Г.Н., Бочарников М.В. 2017. Оробиомы как базовые единицы региональной оценки биоразнообразия горных территорий. – Экосистемы: экология и динамика. 1 (2): 52–81.
[Panferova, Chebakova] Парфенова Е.И., Чебакова Н.М. 2009. Биоклиматические модели коренных лесов гор Южной Сибири. – Лесоведение. 5: 34–42.
[Polikarpov et al.] Поликарпов Н.П., Чебакова Н.М., Назимова Д.И. 1986. Климат и горные леса Южной Сибири. Новосибирск. 225 с.
Popov S.Yu. 2018. Distribution pattern of seven Polytrichum species in the East European Plain and Eastern Fennoscandia. – Botanica Pacifica. A journal of plant science and conservation. 7 (1): 25–40. https://doi.org/10.17581/bp.2018.07108
[Prozorov] Прозоров Ю.С. 1961. Болота маревого ландшафта Средне-Амурской низменности. М. 123 с.
Retuerto R., Carballeira A. 1990. Phytoecological importance, mutual redundancy and phytological threshold values of certain climatic factors. – Vegetatio. 90: 47–62. https://doi.org/10.1007/BF00045588
Retuerto R., Carballeira A. 1992. Use of direct gradient analysis to study the climate-vegetation relationships in Galicia, Spain. – Vegetatio. 101: 183–194. https://doi.org/10.1007/BF00033201
Rivas-Martinez S. 1996. Clasificacion bioclimatica de la Tierra. – Folia Bot Matritensis. 16: 1–32.
Rivas-Martinez S., Armaiz C. 1984. Bioclimatologia y vegetacion en la Peninsula Iberica. – In: Bulletin de la Societe Botanique de France. Actualites Botaniques. 131 (2–4): 110–120.
Rivas-Martinez S., Canto P., Fernandez-Gonzalez F., Molina J.A., Pizarro J.M., Sanchez-Mata D. 1999. Synopsis of the Sierra de Guadarrama vegetation. – Itinera Geobotany. 13: 189–206.
Rivas-Martinez, Penas A., Diaz T.E. 2004. Bioclimatic map of Europe, thermoclimatic belts. Cartographic Service. University of Leon, Spain.
Rivas-Martinez S., Rivas Saenz S., Penas A. 2011. World-wide bioclimatic classification system. – Global Geobotany. 1 (1): 1–634. https://doi.org/10.5616/gg110001
Rivas-Martinez S., Rivas-Saenz S., Penas A. 2016. Synoptical table of worldwide bioclimatic classification. Phytosociological Research Center, Spain. www.globalbioclimatics.org.
[Sedel’nikov] Седельников В.П. 1988. Высокогорная растительность Алтае-Саянской горной области. Новосибирск. 222 с.
[Shumilova] Шумилова Л.В. 1962. Ботаническая география Сибири. Томск. 440 с.
[Sofronov] Софронов А.П. 2015. Геоботаническое картографирование растительного покрова котловин Северо-Восточного Прибайкалья. – Геоботаническое картографирование. С. 62–77.
[Sochava] Сочава В.Б. 1964. Классификация и картографирование высших подразделений растительности Земли. – В кн.: Современные проблемы географии. М. С. 167–173.
[Sochava] Сочава В.Б. 1979. Растительный покров на тематических картах. Новосибирск. 189 с.
[Sochava] Сочава В.Б. 1980. Географические аспекты сибирской тайги. Новосибирск. 256 с.
Stonevicius E., Stankunavicius G., Rimkus E. 2018. Continentality and Oceanity in the Mid and High Latitudes of the Northern Hemisphere and Their Links to Atmospheric Circulation. – Advances in Meteorology. Article ID 5746191, 12 pages. https://doi.org/10.1155/2018/5746191
[Telyatnikov] Телятников М.Ю. 2015. Синтаксономия альпийских лугов, лиственничных редколесий, ерниковых и лишайниковых тундр высокогорий Восточного Саяна. – Растительный мир Азиатской России. 1 (14): 49–65.
[Tolmachev] Толмачев А.И. 1948. Основные пути формирования растительности высокогорных ландшафтов северного полушария. – Бот. журн. 33 (2): 161–180.
Tuhkanen S. 1984. A circumboreal system of climatic-phytogeographical regions. – Acta Botanica Fennica. 127: 1–50.
Valencia-Barrera R., Comtois P., Fernandez-Gonzalez D. 2002. Bioclimatic indices as a tool in pollen forecasting. – Int. J. Biometeorol. 46 (4): 171–175. https://doi.org/10.1007/s00484-002-0138-y
[Vladimirov et al.] Владимиров И.Н., Софронов А.П., Сороковой А.А., Кобылкин Д.В., Фролов А.А. 2014. Структура растительного покрова западной части Верхнеангарской котловины. – География и природные ресурсы. 2: 44–53.
Walter H. 1985. Vegetation of the earth and ecological systems of the geo-biosphere. 3rd edn. N. Y. 527 p.
Walter H., Breckle S.-W. 1991. Okologishe Grundlagen in global sicht. Stuttgart: G. Fischer. 586 p.
Ward J.H. 1963. Hierarchical grouping to optimize an objective function. – Journal of the American Statistical Association. 58: 236–244. https://doi.org/10.1080/01621459.1963.10500845
Yim Y.J., Kira T. 1975. Distribution of forest vegetation and climate in the Korean Peninsula I. Distribution of some indices of thermal climate. – Japanese Journal of Ecology. 25 (2): 77–88. https://doi.org/10.18960/seitai.25.2_77
[Zibzeev et al.] Зибзеев Е.Г., Басаргин E.A., Недовесова Т.А. 2015. Разнообразие и эколого-ценотические особенности березовых криволесий с Betula tortuosa Ledeb. в Алтае-Саянской горной области. – Растительность России. 26: 38–54.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал