

Ботанический журнал, 2022, T. 107, № 7, стр. 672-679
СТРОЕНИЕ И РАЗВИТИЕ ГИНЕЦЕЯ LEDEBOURIA SOCIALIS (ASPARAGACEAE) КАК КЛЮЧ К ПОНИМАНИЮ ЭВОЛЮЦИИ СЕПТАЛЬНЫХ НЕКТАРНИКОВ В ПОРЯДКЕ ASPARAGALES
М. В. Ремизова *
1 Московский государственный университет имени М.В. Ломоносова,
биологический факультет, кафедра высших растений
119234 Москва,
Ленинские горы, 1, стр. 12, Россия
* E-mail: margarita.remizowa@gmail.com
Поступила в редакцию 26.01.2022
После доработки 15.02.2022
Принята к публикации 22.02.2022
- EDN: HFHLLV
- DOI: 10.31857/S0006813622050064
Аннотация
Для Ledebouria socialis характерна верхняя завязь с интерлокулярными септальными нектарниками, которые приурочены как к асцидиатной, так и пликатной зонам плодолистика. Зональность гинецея невозможно описать непротиворечиво, так как существующая терминология не позволяет отобразить все его особенности. Часть нектарников, расположенная в гинофоре и асцидиатной зоне интернализована с самого начала развития, для их формирования не требуется постгенитальное срастание. Для формирования дистальной части нектарника, приуроченной к пликатной зоне плодолистика, постгенитальное срастание необходимо для разграничения нектарных полостей со стороны центра гинецея. Интернализация нектарника по периферии гинецея происходит через разрастание тканей цветоложа. Строение нектарников, характерное для Ledebouria, можно обнаружить у многих Asparagales c верхней завязью.
Ledebouria socialis (Baker) Jessop – популярное комнатное растение, родиной которого является Южная Африка (Baker, 1870). Его выращивают в основном из-за красивых пятнистых листьев. Цветки Ledebouria мелкие и невзрачные, но при ближайшем рассмотрении обнаруживают некоторое изящество. В силу доступности материала Ledebouria представляет собой удобный объект для изучения строения и развития цветка у представителей семейства Asparagaceae.
Цветки Ledebouria имеют типичный для однодольных план строения. Гинецей с верхней трехгнездной завязью. В каждом гнезде расположены по 2 семязачатка, микропиле которых направлены к основанию завязи. Анатомия гинецея с интерпретацией его зональности по W. Leinfellner (1950) представлена в работе O. Dyka11 (2011). В основании завязи находится короткая стерильная синасцидиатная зона. Над синасцидиатной зоной в перегородках между гнездами завязи появляются щелевидные нектарники. Эта зона определяется как гемисинасцидиатная (Dyka, 2011). Гемисинасцидиатная зона фертильна, раздвоенные плаценты, несущие по два семязачатка, расположены в дистальном участке этой зоны. Нектарники продолжаются выше в гемисимпликатную зону, которая, по определению автора фертильна частично (на 1.9% от общей высоты завязи). Выше уровня открытия нектарников расположена асимпликатная зона, она формирует крышу завязи и столбик. В зрелом гинецее Ledebouria линии постгенитального срастания не прослеживаются. В установлении зон гинецея по W. Leinfellner (1950, O. Dyka (2011) во многом опиралась на данные по развитию цветка (Sattler, 1973). R. Sattler (1973), в первую очередь, интересовала последовательность заложения органов цветка. Особенностям строения гинецея уделено мало внимания, даже не указано наличие септальных нектарников (Sattler, 1973). Плодолистики, по его мнению, закладываются тремя отдельными серповидными примодиями, которые почти с самого начала соединены друг с другом по периферии гинецея. Плодолистики из серповидных становятся подковообразными, их края растут по направлению к центру гинецея. Одновременно с ростом краев плодолистиков апекс цветка слегка удлиняется, цветоложе становится выпуклым. Свободные части плодолистиков удлиняются, формируя столбик и завязь. Постгенитальное срастание плодолистиков друг с другом и зарастание брюшного шва завершают формирование гинецея. Семязачатки по два на гнездо закладываются на подвернутых внутрь краях плодолистика еще до начала постгенитальных срастаний.
Описание развития гинецея у R. Sattler (1973) и зональность гинецея, установленные на данных по развитию О. Дика (Dyka, 2011), несколько противоречат друг другу. Так, R. Sattler (1973) явно указывает, что семязачатки закладываются на подвернутых внутрь краях плодолистика. Следовательно, речь идет о пликатной зоне плодолистика и симпликатной или асимпликатной зоне гинецея. Также с точки зрения базовой архитектуры плодолистика не совсем понятно, как фертильными могут быть одновременно две зоны асцидиатная и пликатная, ведь в данном случае на плаценте нужно разместить всего два семязачатка.
Понимание устройства гинецея Ledebouria важно не столько само по себе, сколько для понимания эволюции септальных нектарников у Asparagales и вообще у однодольных. Ранее неоднократно высказывали мнение, что для формирования гинецея с септальными нектарниками необходимо постгенитальное срастание между плодолистиками в той области, где расположены нектарники (van Heel, 1988; Smets et al., 2000; Rudall, 2002a). Недавно была изложена точка зрения, что для формирования септальных нектарников в ходе развития гинецея наличие постгенитального срастания между плодолистиками не обязательно, и нектарники такого типа как раз могут быть распространены у представителей Asparagales (Odintsova22, 2013).
Для выяснения спорных вопросов нами изучено анатомическое строение и развитие гинецея Ledebouria socialis с помощью сканирующего электронного микроскопа.
МАТЕРИАЛЫ И МЕТОДИКА
Материал по развитию соцветий был взят из коллекции комнатных растений кафедры высших растений биологического факультета МГУ им. М.В. Ломоносова и из личной коллекции автора. Луковицы были зафиксированы в 70% спирте, а затем отпрепарированы под стереомикроскопом Olympus SZX7. Подготовленные соцветия на разных стадиях развития, цветки или их части были высушены в критической точке в аппарате Hitachi HCP-2, затем смонтированы с помощью углеродных дисков на столики для электронной микроскопии и напылены тяжелыми металлами с помощью установки Eiko IB-3. Материал был изучен под сканирующим электронным микроскопом CAMSCAN S2 в Общефакультетской лаборатории электронной микроскопии МГУ.
Для изучения анатомии гинецея из коллекции кафедры высших растений были взяты постоянные препараты серийных микротомных поперечных срезов цветка, подготовленные А.С. Беэром и используемые для проведения курса “Морфогенез и васкулатура цветка”. Фотографии срезов были получены под световым микроскопом Olympus BX53, оснащенного цифровой камерой Olympus SC50 под управлением программы Olympus CellSens Entry.
Полученные цифровые изображения были обработаны в графическом редакторе Adobe Photoshop Elements.
РЕЗУЛЬТАТЫ
Строение гинецея. Плодолистики чередуются с внутренними тычинками и срастаются по всей своей длине. Гинецей на всем протяжении трех-гнездный. Завязь верхняя, ребристая, возвышается на коротком массивном гинофоре (рис. 1). Ложбинки вдоль завязи расположены напротив тычинок. Более глубокие ложбинки формируются вдоль линий срастания плодолистиков, в них в бутоне входят нити внутренних тычинок. На спинках плодолистиков имеются менее глубокие ложбинки, в которых в бутоне помещаются нити наружных тычинок.
Рис. 1.
Строение гинецея Ledebouria socialis. а – гинофор, основание, b – гинофор с септальными нектарниками, c – базальная стерильная часть завязи, синасцидиатная зона, d – плаценты, гемисимпликатная зона, e–f – средняя и дистальная часть завязи, гемисимпликатная зона, g – уровень открытия септальных нектарников (отверстие нектарника показано черной стрелкой), асимпликатная зона, h – крыша завязи, асимпликатная зона, i – столбик, асимпликатная зона. Септальные нектарники обведены белой линией. Масштабные линейки – 200 мкм в (a–h) (все срезы в одном масштабе), и 50 мкм в (i).
Fig. 1. Structure of Ledebouria socialis gynoecium. а – gynophore base below septal nectaries. b – gynophore with septal nectaries. c – sterile ovary base, synascidiate zone. d – level of ovule insertion, hemisymplicate zone. e–f – middle and distal ovary part, hemisymplicate zone. g – level of nectary openings (shown by black arrowhead), asymplicate zone. h – ovary roof, asymplicate zone. i – style, asymplicate zone. Septal nectaries are outlined with white in (b–f). Scale bars: a–h – 200 µm (all on the same scale); i – 50 µm.
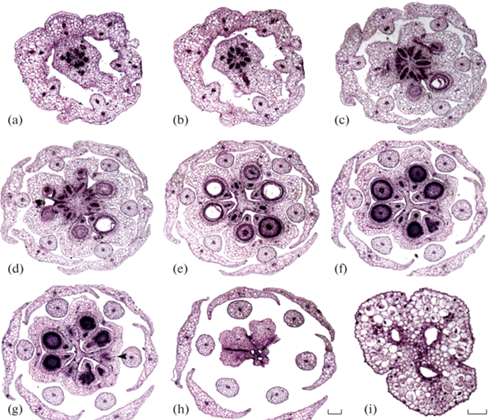
Без установления зональности по W. Leinfellner (1950) гинецей можно описать следующим образом. Септальные нектарники в виде трех отдельных полостей продолжаются из гинофора (рис. 1b) в септы (рис. 1c–1f) и открываются протяженной щелью на верхушке завязи (рис. 1g). Таким образом, гинофор, соответствует конгенитально сросшимся ножкам плодолистиков. Семязачатки прикрепляются на двулопастной плаценте в базальном участке завязи (рис. 1d) и расположены таким образом, что большая их часть помещается выше места прикрепления фуникулусов в несколько выступающих гнездах завязи.
Столбик узкий, с небольшим головчатым рыльцем, на всем протяжении с тремя отдельными каналами, которые ведут в гнезда завязи (рис. 1h, 1i). Отверстия каналов на верхушке гинецея постгенитально замкнуты, изолируя полости гинецея от внешней среды.
Линии постгенитального срастания в зрелом гинецее не заметны, поэтому для избежания ошибок в определении вкладов конгенитального и постгенитального срастания в формировании гинецея необходимо обратиться к его развитию.
Развитие гинецея. Данные по развитию цветка были частично опубликованы ранее (Remizowa et al., 2010), без описания зональности гинецея. Для иллюстрации положений данной статьи использованы новые изображения. Плодолистики закладываются тремя отдельными подковообразными примордиями (рис. 2а). Плодолистики начинают удлиняться, формируя пликатные зоны (рис. 2b–2d). Таким образом, в развитии гинецея первой появляется асимпликатная зона. Очень скоро края плодолистиков заворачиваются внутрь и начинается формирование плацент. Одновременно с заложением семязачатков по периферии гинецея между плодолистиками образуется внешняя стенка завязи (рис. 2b, 2c). Образование этой стенки дает начало заложению гемисимпликатной зоны. Дальнейший рост гинецея связан с увеличением длины гемисимпликатной и асимпликатной зон. Между растущими плодолистиками долгое время заметен голый участок цветоложа. В области завязи из-за формирования крупных семязачатков спинки плодолистиков начинают выпирать в сторону околоцветника, и между плодолистиками формируются глубокие ложбинки. По мере роста гинецея, свободные края плодолистиков сближаются в центре гинецея и происходит их постгенитальное срастание – плодолистики срастаются друг с другом и зарастают брюшные швы (рис. 2e–2h). В гемисимпликатной зоне срастание плодолистиков друг с другом и зарастание брюшных швов приводят к интернализации септальных нектарников – они становятся изолированными как со стороны внешней стенки завязи, так и со стороны центра гинецея. Постгенитальные срастания в гинецее начинаются на достаточно ранних стадиях, еще до полного формирования столбика, поэтому в зрелых цветках линии постгенитальных срастаний не видны. Столбик начинает удлиняться после формирования завязи (рис. 2i, 2k). Асцидиатные зоны плодолистиков и гинофор формируются путем зонального роста под основанием гемисимпликатной зоны (рис. 2j–2k). Для образования нектарников в этой части гинецея постгенитальные срастания не задействованы.
Рис. 2.
Развитие гинецея Ledebouria socialis. а – плодолистики на стадии подковообразных примордиев. заложение асимпликатной зоны. b – заложение семязачатков (*) и образование гемисимпликатной зоны. c, d – рост плодолистиков в длину, увеличение размеров асимпликатной зоны. e – внешний вид гинецея на момент начала постгенитального срастания между плодолистиками. f – гинецей с удаленным плодолистиком, начало постгенитальных срастаний в гемисимпликатной и асимпликатной зонах, область постгенитального срастания отмечена скобкой на левом плодолистике. g – внешняя стенка завязи в области септального нектарника, образованная в результате конгенитального срастания. h – гинецей перед удлинением столбика, вид сверху; плодолистики постгенитально срослись друг с другом, брюшные швы не замкнуты. i – гинецей перед удлинением столбика, вид сбоку. j – внешняя стенка завязи в области нектарника. k – вид нектарной щели сверху в сформированном гинецее. * – Семязачатки. Белыми стрелками показана верхняя граница гемисимпликатной зоны/нижняя граница нектарной щели. Черными стрелками показана верхняя граница нектарной щели. Масштабные линейки – 30 мкм в (a–d, g, h, j, k) и 100 мкм в (e, f, i).
Fig. 2. Development of Ledebouria socialis gynoecium. а – horseshoe-shaped carpel primordia. initiation of asymplacate zone. b – ovule initiation (*) and formation of hemisymplicate zone. c, d – elongation of young carpels, enlargement of asymplicate zone. e – gynoecium at beginning of postgenital carpel fusion. f – same stage as in (e) with one carpel removed to show areas of postgenital intercarpellary fusion in hemisymplicate and asymplicate zones, area of postgenital fusion is marked by an arc on the left carpel. g – outer ovary wall below the future opening of septal nectary, developed via congenital fusion. h – gynoecium before style elongation, view from above. margins of adjacent carpels are postgenitally fused, ventral slits are still open. i – gynoecium before style elongation, lateral view. j – outer ovary wall below nectary opening. k – slit of septal nectary opening in late bud. * – Ovules. White arrowheads show upper boundary of hemisymplicate zone/lower boundary of septal nectary opening. Black arrowheads show upper boundary of septal nectary opening. Scale bars: a–d, g, h, j, k – 30 µm; e, f, i – 100 µm.

ОБСУЖДЕНИЕ
Топология, или зональность гинецея согласно W. Leinfellner (1950), построена с учетом типа срастания между плодолистиками и с учетом того, что у типичного плодолистика имеются базальная асцидиатная и дистальная пликатная зоны. Кратко применение этой терминологии к гинецеям со сросшимися плодолистиками можно описать так. В синасцидиатной зоне плодолистики срастаются конгенитально своими боковыми и брюшными поверхностями с образованием синкарпного (многогнездного) участка. В случае, если срастание в асцидиатной зоне затрагивает только боковые поверхности плодолистика, оставляя пространство в центре гинецея, то зону следует описывать как гемисинасцилиатную. При конгенитальном срастании плодолистиков в пликатной зоне образуется симпликатная зона гинецея. Эта зона исходно одногнездная, но может стать вторично многогнездной через постгенитальное срастание плодолистиков в центре гинецея и зарастание брюшных швов. Гемисимпликатная зона образуется, если края плодолистиков в центре гинецея остаются свободными. При постгенитальном же срастании плодолистиков в пликатной зоне образуется асимпликатная зона гинецея, которая может быть как одногнездная, так и многогнездная в зависимости от зарастания брюшных швов.
Следует отметить, что концепция зональности по W. Leinfellner (1950) хорошо работает только для гинецеев с верхней завязью. Гинецеи с нижней завязью описывать достаточно сложно, так как их конструкцию однозначно интерпретировать по предложенной терминологии удается с трудом. Так, при нижней завязи у любого растения практически невозможно определить, где находится граница плодолистика по отношению к внешней стенке завязи, другими словами, участвуют ли ткани плодолистика в образовании этой стенки (Gustafsson, Albert, 1999; Leins, Erbar, 2010). Ориентироваться приходится на тип срастания плодолистиков в центре гинецея. Еще большие сложности возникают, если к нижней завязи добавляются септальные нектарники. Возьмем для примера представителей Asparagales из семейства Amaryllidaceae. При стандартном описании зональности, т.е. учитывая, что ткани плодолистиков участвуют в образовании стенки завязи, в основании гинецея находится синасцидиатная зона, затем следуют симпликатная и гемисимпликатная (в ней расположены нектарники) зоны, асимпликатная зона составляет крышу завязи и столбик (Fishchuk, 2021; Fishchuk, Odintsova, 2021a,b). Отметим, что в типичной гемисимпликатной зоне края плодолистиков должны быть свободны. У описываемых однодольных края плодолистиков в гемисимпликатной зоне срастаются постгенитально, отграничивая нектарники друг от друга в центре гинецея. Если же принять, что внешняя стенка нижней завязи образована без участия спинных частей плодолистиков, а сами плодолистики прикреплены очень косо на сильно вогнутом цветоложе, то нектарники расположены в асимпликатной зоне, для которой срастание краев плодолистиков в центре гинецея вполне ожидаемо.
При описании гинецея с септальными нектарниками при верхней завязи возникают не меньшие сложности. Сам W. Leinfellner (1950) никогда не применял разработанную им терминологию к гинецею такого типа.
Попробуем описать зональность гинецея Ledebouria. Последовательность событий при формировании гинецея в целом соответствует установленной R. Sattler (1973). Обращает на себя внимание характер заложения семязачатков – они закладываются по краям в пликатной зоне плодолистика. Во время их заложения по периферии гинецея подковообразные плодолистики соединены валиком, поэтому фертильна гемисимпликатная, а не гемисинасцидиатная (Dyka, 2011) зона гинецея. На нашем материале мы не обнаружили типичной синасцидиатной зоны, так как нектарники начинаются еще в гинофоре и идут через всю асцидиатную зону плодолистика. Согласно W. Leinfellner (1950), в синасцидиатной зоне плодолистики срастаются конгенитально как боковыми, так и брюшными поверхностями, а в гемисинасцидиатной зоне – только боковыми, оставляя полость в центре гинецея. Для признания области с септальными нектарниками гемисинасцидиатной зоной (хотя и нетипичной) необходимо, чтобы плодолистики в центре гинецея были постгенитально сросшимися. Однако срастание их конгенитальное. Интерпретации данной области как синасцидиатной зоны препятствуют свободные боковые участки плодолистиков в области нектарников. В нашем описании мы приняли решение охарактеризовать область срастания асцидиатных зон плодолистиков как синасцидиатную зону, основываясь на том, что срастание плодолистиков в центре гинецея конгенитальное. Согласно терминологии, в типичной гемисимпликатной зоне края плодолистиков должны быть свободны. В случае же Ledebouria они постгенитально срастаются.
В итоге получается следующая картина. Насколько можно судить по анализу срезов и из данных по развитию, у плодолистиков Ledebouria выражены короткая асцидиатная и длинная пликатная зоны. Брюшные швы в пликатной зоне замкнуты по всей ее длине. Зональность же гинецея достаточно сложна из-за наличия септальных нектарников и с трудом вписывается в терминологию W. Leinfellner (1950). Гинофор (рис. 1а, 1b) соответствует конгенитально сросшимся ножкам плодолистиков. Базальный участок завязи представлен нетипичной синасцидиатной зоной (рис. 1с). Выше расположена фертильная гемисимпликатная зона с постгенитально сросшимися краями плодолистиков в центре гинецея (рис. 1d–1f). Остальная часть гинецея (дистальная часть завязи от уровня открытия септальных нектарников и столбик) образованы асимпликатной зоной (рис. 1g–1i). Септальные нектарники проходят через все зоны гинецея. Для формирования участка нектарников, расположенного в синасцидиатной зоне, постгенитальное срастание не задействовано. Септальные нектарники в этом участке можно охарактеризовать как “раздельные” по классификации А.В. Одинцовой (Odintsova, 2013).
По наличию конгенитального срастания плодолистиков в центре гинецея в области расположения нектарников A. Odintsova (2013) выделила два типа септальных нектарников – “объединенный” и “раздельный”. При формировании нектарников объединенного типа будущие нектарники не отделены друг от друга в центре гинецея. Раздельный тип септальных нектарников представляет три нектарные полости, которые разделены с самого начала развития гинецея (т.е. ассоциированы с конгенитальным срастанием асцидиатных зон плодолистиков в центре гинецея). В своем онтогенетическом и эволюционном развитии раздельные септальные нектарники сначала открыты с внешней стороны. Образование внешней стенки у раздельных нектарников A. Odintsova (2013) понимает как результат углубления нектарных полостей в базальном направлении, тогда в основании гинецея появится зона с конгенитально замкнутыми септальными полостями. Особенно благоприятные условия для возникновения раздельного нектарника создаются в цветке с нижней завязью, внешняя стенка нектарника образуется путем конгенитального прирастания к завязи цветочной трубки. Распространение раздельных нектарников подлежит проверке и уточнению, но, вероятно, этот тип менее распространен. A. Odintsova (2013) считает, что выделенные ей типы септальных нектарников не могут сочетаться в одном гинецее и не переходят один в другой в ходе эволюции.
Ledebouria совершенно не вписывается в эту схему. Во-первых, в развитии гинецея вся условно синасцидиатная зона вместе с гинофором образуются с уже интернализованными нектарниками в результате зонального роста под гемисимпликатной зоной. Во-вторых, в гинецее этого растения сочетаются оба типа нектарников – и раздельный, и объединенный (начиная с гемисимпликатной зоны). Кроме Ledebouria имеются и другие достаточно многочисленные варианты отграниченных снаружи трех отдельных нектарных полостей, которые со стороны центра гинецея в базальной его области разделены “конгенитальными перегородками”, а в дистальной – “постгенитальными”, и протяженность этих участков достаточна, ни одним из них нельзя пренебречь – Allium (Hartl, Severin, 1981; van Heel, 1988; Zuraw et al., 2009), Aspholdelus (van Heel, 1988), Ornithogalum (van Heel, 1988), Scilla (Dyka, 2013). Характерно, что большая часть этих растений, действительно, принадлежит порядку Asparagales. Отметим, что у некоторых Allium, как и у Ledebouria, имеется гинофор с интернализованными нектарниками (Zuraw et al., 2009). По A. Odintsova (2013), образование наружной стенки нектарника в гинецеях с раздельным типом нектарников происходит путем перехода к нижней завязи, в этом случае к гинецею с открытыми нектарниками снаружи прирастает цветочная трубка. При этом автор как будто упускает из виду, что большая часть нектарников при нижней завязи – это как раз нектарники “объединенного” типа. Так, один из самых распространенных типов септальных нектарников при нижней завязи – трехлучевые, часто лабиринтовидные, т.е. без срастания плодолистиков брюшными поверхностями в асцидиатной зоне, например, у Bromeliaceae (Sajo et al., 2004). Если же при нижней завязи нектарные щели изолированы друг от друга, то нектарник приурочен к пликатной зоне плодолистика, например, у Iridaceae (Rudall et al., 2003).
В целом, для понимания устройства гинецея с септальными нектарниками, действительно, необходимо учитывать тип срастания плодолистиков. Но, вероятно, не следует выделять типы нектарников, основываясь только на установлении типа гинецея по W. Lenfellner (1950).
Asparagales – один из немногих порядков однодольных с очень вариабельной структурой гинецея. В эволюции порядка неоднократно были утеряны септальные нектарники, происходила смена положения завязи (Simpson, 1998; Rudall, 2002a,b). По положению относительно гнезд завязи септальные нектарники Asparagales бывают интерлокулярными и инфралокулярными. В последнем типе они расположены на ножках плодолистиков и/или в основании асцидиатной зоны “под гнездами” всегда верхней завязи и обычно открыты с наружной стороны гинецея (Simpson, 1998; Smets et al., 2000; Kocyan, Endress, 2001; Rudall, 2002a,b). У Borya плодолистики при инфралокулярных нектарниках срастаются только постгенитально в пликатной зоне. У большей же части растений с инфралокулярными нектарниками ножки плодолистиков и их асцидиатные зоны срастаются конгенитально. В развитии такие нектарники недостаточно изучены, но на конгенитальный характер срастания указывает наличие синвентральных пучков (Kocyan, Endress, 2001). Интерлокулярные нектарники у представителей Asparagales чаще всего развиваются при нижней завязи (Kocyan, Endress, 2001; Rudall, 2002a,b) и приурочены к пликатной зоне плодолистика. Наличие интерлокулярных нектарников при верхней завязи, как у Ledebouria, представляет собой совершенно особый тип. Выводить его, на наш взгляд, следует от гинецеев с инфралокулярными нектарниками через распространение секреторных поверхностей в пликатную зону и образование внешней стенки завязи, которая, вероятно, образуется как вырост цветоложа (о природе наружной стенки завязи в гинецеях с септальными нектарниками см. также van Heel, 1988). Такой же тип гинецея, как у Ledebouria, характерен для представителей нескольких линий Asparagales, причем сходство обнаруживается не только в строении взрослого гинецея, но и в его развитии. Особенностью этих однодольных растений является раннее начало постгенитальных срастаний в гинецее, поэтому в гинецее зрелых цветков линии срастания между плодолистиками и линии зарастания брюшных швов зачастую плохо различимы или не различимы вообще. Без обращения к развитию цветка эта особенность затрудняет правильную интерпретацию структуры гинецея, особенно при описании строения по W. Leinfellner.
Список литературы отсутствует.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал