

Ботанический журнал, 2022, T. 107, № 8, стр. 781-793
РЕПРОДУКТИВНАЯ БИОЛОГИЯ COLURIA GEOIDES (ROSACEAE)
Т. В. Елисафенко 1, *, В. Н. Годин 1, 2, **
1 Центральный сибирский ботанический сад СО РАН
630090 Новосибирск, ул. Золотодолинская, 101, Россия
2 Московский педагогический государственный университет
129164 Москва, ул. Кибальчича, 6, корп. 3, Россия
* E-mail: tveli@ngs.ru
** E-mail: vn.godin@mpgu.su
Поступила в редакцию 03.05.2022
После доработки 11.06.2022
Принята к публикации 14.06.2022
- EDN: HOEZNM
- DOI: 10.31857/S0006813622080051
Аннотация
Изучены строение семян и плодов, особенности завязывания семян и семенного размножения у травянистого короткокорневищного поликарпического растения Coluria geoides в естественных условиях Горного Алтая, Алтайского края и в интродукционном эксперименте (г. Новосибирск). C. geoides образует 1-, 2- или 3-цветковые синфлоресценции, представляющие собой вариант дихазиальных соцветий. Данный вид относится к ксеногамным по соотношению числа пыльцевых зерен и семязачатков (от 2356 до 3891). Для цветков характерна протогиния, препятствующая автофилии. Показано, что изученные размерные характеристики семян, плодов и их составных элементов обладают низкой или средней вариабельностью. Наоборот, количественные параметры семенной продуктивности (потенциальная и реальная семенная продуктивность, процент семенификации) демонстрируют высокий и очень высокий уровни вариабельности. Число плодолистиков в цветках у особей с 3-цветковыми соцветиями достоверно больше, чем в цветках у особей с 1‑ и 2-цветковыми синфлоресценциями. Также у особей с 3-цветковыми соцветиями реальная семенная продуктивность (в расчете на плод) и процент семенификации плодов, занимающих терминальное положение, выше, чем у плодов, расположенных на пазушных побегах.
Род Coluria R. Br. включает 5 видов, которые встречаются на территории Южной Сибири, Китая и Монголии (Kalkman, 2004). Coluria geoides (Pall.) Ledeb. (колюрия гравилатовидная) – гемиэндемик юга Сибири и северной Монголии. Р.В. Камелин (Kamelin, 1998) определяет ареал вида как восточноказахстанско-алтае-тувинско-северохангайский. Другие 4 вида рода являются эндемиками Китая (Chaoluan et al., 2003; Vydrina, 1988). Таксономическое положение рода дискуссионно, некоторые авторы включают его в состав рода Geum L. (Potter et al., 2007). Камелин (Kamelin, 1998) определяет C. geoides как высокотолерантный вид, по всей видимости лесного генезиса. В роде Coluria прослеживается уникальный пример родственных связей с центрально- и юго-западно-китайскими субтропическими видами.
C. geoides (рис. 1, А, B) – травянистое короткокорневищное поликарпическое растение с эпигеогенным плагиотропным корневищем и моноциклическими удлиненными генеративными побегами. Вид является ксеромезофитом, мезотрофом и факультативным петрофитом, при этом встречается до 2800 м над ур. м. на северных и северо-восточных степных склонах в составе луговых степей, горных и высокогорных степей, щебнистых и каменистых склонов (Polyntseva, Utemova, 1988), а также в разреженных лиственничных и сосновых лесах (Leonova et al., 2010). Это длительновегетирующее, летне-зимнезеленое растение с весенне-раннелетним ритмом цветения и эфемероидным типом развития генеративных побегов. C. geoides – ценное эфироносное растение, основным компонентом которого является эвгинол, включенный в фармакологию и использующийся в стоматологии и кулинарии. Проявляет иммуностимулирующее действие при стафилококковой инфекции (Kuz’mina, Ovcharenko, 1987; Vodolazova et al., 2011; Dutova et al., 2015).
Рис. 1.
Coluria geoides.
А – Республика Алтай (9 июня 2016 г.), В – экспозиция “Редкие и исчезающие виды растений Сибири” (г. Новосибирск, 3 мая 2012 г.), С – генеративный побег (Республика Алтай, 16 июня 2016 г.), D – схема синфлоресценции. a – 4-цветковый дихазий; b – 3-цветковый дихазий; c – 2-цветковый дихазий; d – одноцветковый дихазий. Фото Т.В. Елисафенко.
Fig. 1. Coluria geoides. A – Altai Republic (June 9, 2016), B – exposition “Rare and endangered plant species of Siberia” (Novosibirsk, May 3, 2012), С – generative shoot (Altai Republic, June 16, 2016), D – synflorescence scheme. a – 4-flowered dichasium; b – 3-flowered dichasium; c – 2-flowered dichasium; d – 1-flowered dichasium. Photo by T.V. Elisafenko.

C. geoides долгое время включали в региональные списки редких и исчезающих видов растений (Redkie…, 1980), а также в “Красную книгу Республики Алтай” (Orlov, 1996), Красную книгу Красноярского края (Sonnikova, 2005). В настоящее время данный вид входит только в Красную книгу Красноярского края (Sonnikova, 2012), поскольку на юге этого региона находится северная граница ареала, причем отмечается, что лимитирующими факторами являются изменение микроклимата в связи со строительством водохранилища и непосредственное уничтожение местообитаний. Т.В. Леонова с соавторами (Leonova et al., 2010) установила, что неблагоприятными условиями для роста и развития растений на территории Хакасии являются сильно увлажненные участки и бедные почвы, а также моховой покров, каменистые степи, антропогенная нагрузка (весенние палы степной растительности, выпас скота). Н.А. Полынцева и Л.Д. Утемова (Polyntseva, Utemova, 1988) сделали вывод о нецелесообразности сохранения данного вида в культуре и необходимости сохранять популяции C. geoides в естественных условиях.
Согласно данным литературы, поддержание популяций C. geoides осуществляется семенным и вегетативным способами (Elisafenko, 2008; Leonova, 2012). На основании проведенных ранее исследований в Горном Алтае установлено (Eli-safenko, 2008), что для C. geoides свойственно незначительное как вегетативное, так и семенное размножение, а также уязвимость к антропогенному воздействию. Реальная семенная продуктивность в расчете на генеративный побег составила от 11 до 33 семян, причем у 30% из них отсутствовал зародыш, и в итоге число полноценных семян составило 8 шт. (Elisafenko, 2008). В Хакасии Т.В. Леонова (Leonova, 2012) выявила аналогичную довольно низкую реальную семенную продуктивность в расчете на генеративный побег – 13.4–34.0 шт. и также отметила наличие пустосемянности.
Семенная продуктивность растений напрямую зависит от особенностей строения синфлоресценций. Виды рода Coluria, как и другие представители семейства Rosaceae, характеризуются монотелическими цимозными синфлоресценциями (Troll, 1969). В литературе нет однозначной трактовки синфлоресценций данного вида, которые, как правило, немного- (1–4) цветковые. Некоторые авторы относят синфлоресценции C. geoides к редуцированному многочленному монохазию (Cheryomushkina, Leonova, 2011).
Цель данного исследования – изучить семенную продуктивность и морфологию семян у C. geoides в зависимости от особенностей строения и пространственного положения в синфлоресценциях в различных эколого-фитоценотических условиях Горного Алтая и Алтайского края.
МАТЕРИАЛЫ И МЕТОДЫ
Изучение семенной продуктивности и морфологии семян Coluria geoides проводили в естественных условиях Алтайского края и Горного Алтая. Изучено 5 ценопопуляций (ЦП) C. geoides в различных эколого-фитоценотических условиях.
Семигумидный сектор Горного Алтая.
ЦП 1. Закустаренная каменистая луговая степь, Горный Алтай, Усть-Канский р-н, окр. с. Кайсын. Общее проективное покрытие (ОПП) – 60%. Доминанты: Carex pediformis C.A. Mey., Helictotrichon altaicum Tzvelev.
ЦП 2. Закустаренная каменистая луговая степь, Горный Алтай, Усть-Канский р-н, спуск Ябоганского перевала. ОПП – 60%. Доминанты: Spiraea trilobata L., Carex pediformis, C. humilis Leyss.
Переход к семиаридному сектору Горного Алтая.
ЦП 3. Закустаренная каменистая дерновинно-стоповидноосоковая степь, Горный Алтай, Онгудайский р-н, окр. п. Чуй-Оозы. ОПП – 50%. Доминант: Carex pediformis.
ЦП 4. Каменистая полынная дерновинно-злаковая степь, Горный Алтай, Онгудайский р-н, между поселками Купчегень и Иня. ОПП – 30%. Доминант: Koeleria cristata (L.) Pers.
Алтайский край, степная зона.
ЦП 5. Каменистая луговая степь, Алтайский край, Курьинский р-н, окр. оз. Колыванское. ОПП – 30%. Доминанты: Carex pediformis, C. humilis.
У C. geoides встречаются особи с разным числом цветков, входящих в синфлоресценции: с одним, двумя или тремя цветками. В каждой ЦП выявляли долю особей с разными вариантами разветвленности синфлоресценций. Для этого случайным образом собирали от 70 до 300 генеративных побегов в зависимости от общей численности вида в ЦП. Из этой выборки генеративных побегов в каждой ЦП у 30–40 синфлоресценций определяли размеры плодов и выявляли семенную продуктивность по методике И.В. Вайнагия (Vaynagiy, 1974). При этом подсчитывали потенциальную семенную продуктивность (число семязачатков в цветке), реальную семенную продуктивность (число образовавшихся полноценных семян в расчете на плод) и процент семенификации как отношение реальной семенной продуктивности к потенциальной (выраженной в процентах). Плоды, сформированные особями с разными вариантами разветвленности синфлоресценций, а также плоды в пределах 2–3 цветковых синфлоресценций C. geoides собирали и анализировали отдельно. Для каждого плода определяли следующие параметры: длина и диаметр плода, отношение длины плода к его диаметру.
В камеральных условиях семена, собранные с 30–40 синфлоресценций разной разветвленности, хранили и анализировали отдельно для выявления характера зависимости их размерных показателей от положения в синфлоресценции. Измерения размера семян проводили на оборудовании центра коллективного пользования ЦСБС СО РАН при помощи стереомикроскопа Carl Zeiss Stereo Discovery V12 при увеличении 10–30 раз с цветной цифровой камерой высокого разрешения AxioCam MRс-5 и программой AxioVision 4.8 для получения, обработки и анализа изображений в 30-кратной повторности. Семена перед измерениями замачивали на 12–18 ч. Для каждого семени определяли следующие параметры: длина, ширина и толщина семени, длина и ширина зародыша, длина семядолей, длина оси зародыша, отношение длины семени к его ширине, отношение длины зародыша к длине семени (в %), отношение длины семядоли к длине оси зародыша (в %).
Для изучения соотношения числа пыльцевых зерен и семязачатков использовали 10 цветков, зафиксированных в 70%-ном этиловом спирте. Подсчет пыльцевых зерен производили по общепринятой методике (Cruden, 1977). Для каждого цветка оценивали число пыльцевых зерен в одной из тек одного произвольно выбранного невскрывшегося пыльника, а также подсчитывали мерность андроцея и гинецея выбранного цветка. Для подсчета пыльцевых зерен в препаратах использовали оптический микроскоп Axio Lab. A1 (Carl Zeiss) с цветной цифровой камерой высокого разрешения Axiocam MRc 5 с программой AxioVision 4.8 при увеличении объектива 40× с окуляр-микрометрометром с сеточкой. Так как пыльник состоит из двух тек, количество пыльцы в пыльнике получали, удваивая ее численность в одной теке. Для оценки числа пыльцевых зерен, продуцируемых цветком, полученное содержание пыльцы в пыльнике умножали на число тычинок в данном цветке. Поскольку для трибы Colurieae характерно наличие единственного фертильного семязачатка в каждом гнезде завязи (Potter et al., 2007), отношение числа пыльцевых зерен к числу семязачатков оценивали для каждого цветка делением числа пыльцевых зерен в цветке на число плодолистиков.
Все полученные данные обрабатывали с помощью непараметрических методов анализа (Sokal, Rohlf, 2012). Перед проведением статистического анализа данные тестировали на нормальность распределения. Степень асимметричности распределения показателей изученных признаков вычисляли через отношение моментов третьего порядка к кубу среднего квадратичного отклонения. Оценку достоверности показателя асимметрии производили через отношение показателя асимметрии к его ошибке. Значения показателей асимметрии параметров семян и их составных элементов в разных ценопопуляциях варьировали от 0.08 до 0.24, ошибка – от 0.41 до 0.49. Следовательно, распределение этих признаков в разных ценопопуляциях достаточно близко к симметричному. Распределение значений таких показателей как потенциальная и реальная семенная продуктивность, процент семенификации в разных ценопопуляциях показывал асимметричность. Поэтому для адекватного сравнения всех изученных признаков в дальнейшем использовали медиану и медианный коэффициент вариации (Zhivotovskiy, 1991).
Для оценки степени влияния разных факторов (тип сообщества, мерность синфлоресценции, локализация цветка в соцветии) на изученные показатели семян и семенной продуктивности использовали ранговый дисперсионный анализ для групп равного или неравного объема по методу Краскелла и Уоллиса (Sokal, Rohlf, 2012).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
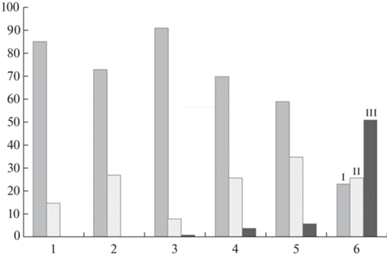
Структура синфлоресценций. Coluria geoides образует 1-, 2- или 3-цветковые синфлоресценции (в условиях интродукционного эксперимента иногда формируются 4-цветковые соцветия) (рис. 1, C, D). Мерность синфлоресценции зависит от степени благоприятности условий произрастания: в интродукционном эксперименте процент особей с 3-цветковыми дихазиями был самый высокий и составлял не менее 50% (рис. 2). Интерпретация таких малоцветковых синфлоресценций не всегда проста. Мы склонны считать, что синфлоресценция у данного вида представляет собой вариант редуцированного дихазия, когда наблюдается недоразвитие одного или обоих пазушных цветков и остается только терминальный цветок. Однако в литературе иногда синфлоресценцию C. geoides описывают как редуцированный многочленный монохазий (Cheryomushkina, Leonova, 2011). Рассмотрим причины, почему мы относим синфлоресценции у C. geoides к дихазиальным, а не монохазиальным вариантам соцветий.
Рис. 2.
Распределение типов синфлоресценций Coluria geoides в разных сообществах и в условиях интродукции.
Условные обозначения: по оси X – популяции, по оси Y – процент типа синфлоресценции в популяции (%). I – одноцветковое соцветие; II – 2-цветковое соцветие; III – 3- цветковое соцветие.
1 – ЦП1, 2 – ЦП2, 3 – ЦП3, 4 – ЦП4, 5 – ЦП5, 6 – интродукционная популяция.
Fig. 2. Distribution of synflorescence types of Coluria geoides in different communities and in conditions of the botanical garden.
Legend: X-axis – populations, Y-axis – percentage of synflorescence types in the population. I – 1-flowered dichasium; II – 2‑flowered dichasium; III – 3-flowered dichasium.
1–5 – numbers of coenopopulations; 6 – population in the botanical garden.
Во-первых, во всех возможных вариантах монохазиев у двудольных растений (улитки и извилины) оси каждого порядка имеют парные профиллы (прицветнички), расположенные всегда трансверзально (Kondorskaya, 1990). В дихазиальных соцветиях парные профиллы расположены всегда медианно (Kondorskaya, 1989), что характерно и для синфлоресценций C. geoides. Необходимо отметить, что специфика внешнего облика дихазиев у представителей сем. Rosaceae заключается в том, что для них характерен не супротивный филлотаксис, а очередный.
Во-вторых, монохазиальные и дихазиальные соцветия различаются характером раскрывания цветков. Как известно, последовательность раскрывания цветков в соцветиях определяется порядком побега, на котором цветки формируются (Kuznetsova, 1992; Kuznetsova, Timonin, 2017). Среди цветковых растений известны единичные исключения из этого правила, описанные у некоторых представителей сем. Apiaceae (Godin, Perkova, 2017; Kuznetsova, Timonin, 2017). Во всех монохазиальных соцветиях первыми зацветают цветки на побегах самых низких порядков, расположенные в основании соцветия. В целом такая последовательность цветения цветков монохазиальных соцветий напоминает “акропетальную” (настоящая акропетальность характерна только для рацемозных соцветий). В дихазиальных соцветиях всегда первым зацветает его терминальный цветок (в случае очередного филлотаксиса расположенный обычно выше пазушных), а после – пазушные цветки на побеге следующего порядка и т.д. У C. geoides характер раскрывания цветков, несомненно, свидетельствует о дихазиальности ее синфлоресценций: первым раскрывается и отцветает терминальный цветок соцветия, а пазушные цветки на побегах следующего порядка цветут синхронно друг с другом и всегда после окончания функционирования терминального.
В-третьих, многие морфологи в области синфлоресценций в случае формирования малоцветковых соцветий у конкретного вида и неоднозначности интерпретации их строения рекомендуют проводить сравнение с близкородственными таксонами (Kuznetsova, 1992). По мнению W. Troll (1969), для всех представителей сем. Rosaceae характерны монотелические синфлоресценции (с терминальным цветком). Многие исследователи этого семейства отмечают, что часто у его представителей наблюдается редукция числа цветков в соцветиях до 1 (Troll, 1969; Tichonova. 1974; Kalkman, 2004; Petuchova, 2000). Род Coluria относится к трибе Colurieae, близкой к трибе Potentilleae. Согласно многим исследованиям (Focke, 1891; Wolf, 1908; Kuznetsova, 1992), основной вариант синфлоресценций у обоих триб – дихазии, часто с элементами редукции числа цветков до единственного. На основании всех вышеперечисленных фактов, все-таки мы считаем, что синфлоресценции C. geoides представляют собой вариант дихазия.
Соотношение пыльцевых зерен и семязачатков. Соотношение числа пыльцевых зерен и семязачатков – один из важных показателей, характеризующих успешность опыления у растений. По мнению R.W. Cruden (1977), этот показатель отражает вероятность попадания на каждое рыльце количества пыльцы, необходимого для наиболее полного завязывания семян. Другими словами, это минимальное количество пыльцы, при повышении которого не увеличивается семенная продуктивность. Чем выше эффективность транспорта пыльцы от тычинок к рыльцам, тем ниже этот показатель. Таким образом, соотношение числа пыльцевых зерен и семязачатков скоррелировано с типом системы размножения вида. Ниже всего данный показатель у растений, для которых характерна автогамия, особенно клейстогамия (Cruden, 1977, 2000).
Наши исследования показали, что число пыльцевых зерен в микроспорангиях тычинок у C. geoides варьирует от 1100 до 2360. Число тычинок в изученных нами цветках колеблется от 35 до 65, а плодолистиков – от 8 до 23. Следовательно, в цветках соотношение числа пыльцевых зерен и семязачатков варьирует от 2356 до 3891, что согласно представлениям R.W. Cruden (1977) позволяет отнести C. geoides к ксеногамным видам. Например, у близкого вида Geum urbanum L. это соотношение составляет 999, поэтому он относится к таксонам с факультативной ксеногамией (Weber, 1995).
Особенности опыления и завязывания семян. Энтомофильный облик цветков C. geoides (яркая окраска венчика, довольно крупные цветки, открытость и доступность нектарного кольца, многомерность андроцея с доступной пыльцой и др.) позволяет предполагать, что круг насекомых-опылителей и посетителей довольно широк. Наши немногочисленные наблюдения в природе и в условиях интродукционного эксперимента показали, что цветки C. geoides посещаются в основном представителями двух отрядов насекомых – Hymenoptera и Diptera. Для цветков этого вида характерна протогиния – более ранее созревание рылец в сравнении с пыльниками. После раскрывания цветка тычинки согнуты к готовым к опылению плодолистикам, пыльники их не вскрыты. Насекомые, посещающие в это время цветки C. geoides в поисках нектара, спрятанного глубоко в цветке между андроцеем и гинецеем, осуществляют перекрестное опыление за счет пыльцы с других цветков. После окончания функционирования рылец, тычинки удлиняются, выпрямляются и располагаются параллельно чашелистикам, образуя в цветках своеобразную прочную “щетку”, снабженную большим числом вскрытых пыльников с многочисленной пыльцой. В эту фазу цветения (пыльниковую) цветка насекомые посещают их ради пыльцы, поскольку им совершенно невозможно добраться до нектарного диска. Такой характер временного функционирования репродуктивных органов значительно уменьшает вероятность автогамии в цветках C. geoides с их полимерными андроцеем и гинецеем.
Наши опыты по изоляции еще нераскрывшихся цветков C. geoides в условиях интродукционного эксперимента показали, что под марлевыми изоляторами завязывание семян не наблюдалось. Следовательно, случайный перенос пыльцы в пределах цветка на рыльца у данного вида практически невозможен из-за наличия временного разрыва в функционировании его репродуктивных органов (протогинии). С другой стороны, данный опыт также показывает отсутствие вероятности образования семян у C. geoides за счет апомиксиса. Хотя в близком роде Potentilla разные формы апомиксиса широко распространены (Müntzing, Müntzing, 1941), у представителей других близкородственных таксонов (Geum, Dryas и др.) апомиктичное завязывание семян происходит очень редко (Gajewski, 1959). Поэтому можно предположить, что для ксеногамного вида C. geoides также не характерно образование семян при помощи апомиксиса.
Морфология и размерные характеристики семян и семенной продуктивности. Семена C. geoides заключены в орешки, образующие плоды многоорешки. Орешки C. geoides продолговато-яйцевидные, 2.3–3.3 мм длиной, 1.2–1.5 мм шириной и 1.2–1.5 мм толщиной (рис. 3), коричневого цвета, покрытые стекловидными сосочками. По мнению F. Boll (1933), эти сосочки представляют собой приспособление для распространения семян этого вида муравьями. Это связано с тем, что среди клеток экзокарпия располагаются крупные масляные клетки, которые содержат жирные масла, служащие аттрактантом для муравьев. Зародыш хорошо развит, крупный, хорошо дифференцирован и занимает почти всю внутреннюю часть семени.
Рис. 3.
Орешек (А) и зародыш (В) Coluria geoides.
Fig. 3. Nutlet (A) and embryo (B) of Coluria geoides.

У C. geoides гипантий и чашечка сохраняются при плодах, в результате чего многоорешек полностью в них погружен. Из-за этого плод у этого вида длинно колокольчатой формы, 1.0–1.4 см дл. и диаметром 0.5–0.7 см. Число выполненных семян в одном плоде варьирует в широких пределах (от 0 до 20), что также приводит к большой изменчивости в завязываемости семян: процент семенификации колебался от 0 до 100%, составляя, около 70%. Масса 100 выполненных семян довольно слабо варьирует: от 0.09 до 0.12 г. Реальная семенная продуктивность в расчете на побег варьирует у 1-цветковых особей от 6 до 18 семян, у 2-цветковых особей – от 21 до 39 семян и у 3‑цветковых особей от 36 до 64 семян.
Анализ размерных показателей семян, их структурных элементов и характеристик семенной продуктивности показал следующее (рис. 4). По степени варьирования (коэффициенты вариации) значений изученные признаки разделяются на две группы: размерные и счетные. Все размерные показатели семян, плодов, зародышей характеризуются слабым или средним варьированием своих значений: медианные коэффициенты вариации колеблются от 3.6 до 24.0% (рис. 5). Количественные показатели семенной продуктивности (потенциальная и реальная семенная продуктивности, процент семенификации) отличаются высоким и очень высоким варьированием: медианные коэффициенты вариации больше 29%. В целом это общая закономерность варьирования мерных и счетных признаков (Mamaev, 1973). Размерные показатели семян и их составных элементов относятся к генеративным органам, изменчивость которых жестко контролируется естественным отбором, что способствует их малой или невысокой вариабельности. Напротив, мерность гинецея в цветке, число сформированных полноценных семян в плоде и соотношение этих показателей относятся к стохастическим характеристикам и определяются вероятностными причинами. У многих представителей сем. Rosaceae, в том числе и у C. geoides, число плодолистиков в цветках не фиксированное (Kalkman, 1988). Число выполненных семян в плодах и процент семенификации полностью определяются успешностью опыления, которое зависит от многих факторов, не подпадающих под прямое действие естественного отбора.
Рис. 4.
Размерные характеристики и показатели семян и семенной продуктивности у Coluria geoides.
Условные обозначения: по оси X – признаки, по оси Y – значения медиан признаков.
I – максимальное значение, II – минимальное значение, III – квартиль, IV – медиана.
1 – длина (мм), 2 – ширина (мм), 3 – толщина семени (мм), 4 – длина (мм), 5 – ширина зародыша (мм), 6 – длина семядоли (мм), 7 – длина оси зародыша (мм), 8 – соотношение длины и ширины семени, 9 – длина плода (см), 10 – диаметр плода (см), 11 – соотношение длины и диаметра плода, 12 – масса семян (г), 13 – потенциальная семенная продуктивность (шт.), 14 – реальная семенная продуктивность (шт.), 15 – процент семенификации (%), 16 – отношение длины зародыша к длине семени, 17 – отношение длины семядоли к длине зародыша.
Fig. 4. Dimensional characteristics and indicators of seeds and seed set in Coluria geoides
Legend: X-axis – traits, Y-axis – values of trait medians (mm or %).
I – maximum value, II – minimum value, III – quartile, IV – median.
1 – length (mm), 2 – width (mm), 3 – seed thickness (mm), 4 – length (mm), 5 – embryo width (mm), 6 – cotyledon length (mm), 7 – embryo axis length (mm), 8 – seed length/width ratio, 9 – fruit length (cm), 10 – fruit diameter (cm), 11 – fruit length/diameter ratio, 12 – mass of seeds (g), 13 – number of ovules per flower (pcs.), 14 – number of seeds per fruit (pcs.), 15 – ratio of seed number per fruit to ovule number per flower (%), 16 – ratio of embryo length to seed length, 17 – ratio of cotyledon length to embryo length.

Рис. 5.
Медианный коэффициент вариации показателей семян и семенной продуктивности у Coluria geoides.
Условные обозначения: по оси X – признаки, по оси Y – значения медианного коэффициента вариации признаков (%).
I – максимальное, II – минимальное значение, III – квартиль, IV – медиана.
1 – длина семени, 2 – ширина семени, 3 – толщина семени, 4 – длина зародыша, 5 – ширина зародыша, 6 – длина семядоли, 7 – длина оси зародыша, 8 – отношение длины зародыша к длине семени, 9 – отношение длины семядоли к длине зародыша, 10 – соотношение длины и ширины семени, 11 – длина плода, 12 – диаметр плода, 13 – соотношение длины и диаметра плода, 14 – масса семян, 15 – потенциальная семенная продуктивность, 16 – реальная семенная продуктивность, 17 – процент семенификации.
Fig. 5. Median coefficient of variation of seed and seed set indicators in Coluria geoides.
Legend: X-axis – traits, Y-axis – median coefficients of trait variation (%).
I – maximum value, II – minimum value, III – quartile, IV – median.
1 – seed length, 2 – seed width, 3 – seed thickness, 4 – embryo length, 5 – embryo width, 6 – cotyledon length, 7 – length of embryo axis, 8 – ratio embryo length to seed length, 9 – ratio of cotyledon length to embryo length, 10 – seed length/width ratio, 11 – fruit length, 12 – fruit diameter, 13 – fruit length/diameter ratio, 14 – weight of seeds, 15 – number of ovules per flower, 16 – number of seeds per fruit, 17 – ratio of seed number per fruit to ovule number per flower.

Поскольку частота встречаемости особей в изученных ценопопуляциях с разным числом цветков в синфлоресценции крайне неравномерна, не представляется возможным провести дисперсионный анализ сразу по всем возможным факторам (тип сообщества, особенности строения соцветий, положение цветка в синфлоресценции), которые могут влиять на размерные показатели семян, плодов и семенную продуктивность у C. geoides. В связи с этим нам пришлось оценивать влияние каждого фактора отдельно.
Тип сообщества. Вид C. geoides изучен нами в двух типах сообществ на территории Алтайского края и Горного Алтая: в настоящих и луговых степях. В этих двух типах сообществ были проанализированы морфометрические показатели семян, плодов и количественные параметры семенной продуктивности для всех особей с разным числом цветков в соцветиях. Применение рангового дисперсионного анализа для групп равного объема показало, что тип сообщества достоверно влияет лишь на один из изученных показателей: длина плода (χ2 = 10.2, P <0.044). По остальным проанализированным параметрам семян и семенной продуктивности влияние типа сообщества крайне мало и статистически недостоверно. Следовательно, в луговых степях у C. geoides формируются более длинные плоды, чем в степных сообществах. Скорее всего, формирование более длинных плодов в луговых сообществах обусловлено сравнительно благоприятными условиями для произрастания данного вида, что выражается в образовании более крупных особей в сравнении со степными фитоценозами.
Число цветков в соцветиях. C. geoides формирует особи с 1-, 2- и 3-цветковыми синфлоресценциями в естественных условиях (рис. 2). Поскольку нами не было выявлено влияния типа сообщества на почти все анализируемые признаки семян и семенной продуктивности, выборки из всех ценопопуляций были использованы для получения обобщенной характеристики параметров особей, различающихся мерностью синфлоресценций (число цветков в соцветиях). Применение рангового дисперсионного анализа для групп неравного объема выявило, что мерность соцветия заметно влияет только на один анализируемый признак – потенциальную семенную продуктивность (χ2 = 11.4, P = 0.032). Таким образом, число плодолистиков в цветках у особей с 3-цветковыми соцветиями достоверно больше (рис. 6, A), чем в цветках у особей с 1- и 2-цветковыми синфлоресценциями (последние достоверно не различаются по данному признаку). Возможно, что наблюдаемое небольшое увеличение числа плодолистиков в цветках у 3-цветковых особей обусловлено их большей мощностью и возможностью вкладывать больше пластических веществ на формирование генеративных органов и как итог большую потенциальную семенную продуктивность.
Рис. 6.
Семенная продуктивность Coluria geoides.
А – число плодолистиков в цветках (потенциальная семенная продуктивность) в соцветиях с разной мерностью. В – процент семенификации в плодах с разным положением в 3-цветковых соцветиях.
Условные обозначения: 1 – число плодолистиков в цветках 1- и 2-цветковых соцветий, 2 – число плодолистиков в цветках 3-цветковых соцветий. 3 – процент семенификации в плодах, развившихся из терминальных цветков, 4 – процент семенификации в плодах, расположенных на боковых побегах. Остальные условные обозначения, как на рис. 4 и 5.
Fig. 6. The seed set of Coluria geoides.
A – number of carpels per flower in inflorescences with different number of flowers, В – ratio of seed number per fruit to ovule number per flower in fruits with different positions in 3-flowered inflorescences.
Symbols: 1 – carpel number in flowers of 1- and 2-flowered inflorescences, 2 – carpel number in flowers of 3-flowered inflorescences. 3 – ratio of seed number per fruit to ovule number per flower in fruits formed on the main shoot, 4 – ratio of seed number per fruit to ovule number per flower in fruits of lateral shoots. For the other symbols see Figs. 4 and 5.
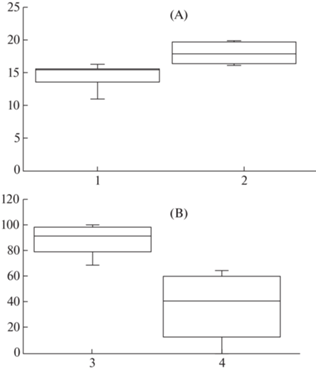
Положение плода в соцветии. В дихазиальных соцветиях C. geoides цветки и, соответственно затем плоды, могут иметь два положения: терминальное и пазушное. Мы проанализировали влияние положения плода у 2- и 3-цветковых соцветий на показатели семян и семенной продуктивности. Использование рангового дисперсионного анализа для групп неравного объема показало, что у особей с 3-цветковыми соцветиями реальная семенная продуктивность и процент семенификации плодов, занимающих терминальное положение, выше, чем у пазушных (χ2 >10.9, P < 0.041) (рис. 6, B). У особей с 2-цветковыми соцветиями такого влияния не выявлено.
Цветки с разным положением в синфлоресценциях цветут в разное время и соответственно успех завязывания ими плодов напрямую зависит от обеспеченности пыльцой. Терминальные цветки в 3-цветковых синфлоресценциях у C. geoides цветут в период, когда в популяциях наблюдается массовое цветение других особей, способных стать источником пыльцы для опыления и завязывания семян. Пазушные цветки в этих соцветиях раскрываются и функционируют позже, когда большая часть особей в популяции отцвела (с 1- и/или 2-цветковыми соцветиями). Поэтому вероятность успешного опыления и завязывания ими плодов значительно снижается. Плоды, занимающие терминальное положение в 3-цветковых синфлоресценциях, всегда завязывают семена, а для плодов, расположенных на пазушных побегах дихазия, очень часто характерна пустосемянность, когда не завязывается ни одно семя.
ЗАКЛЮЧЕНИЕ
1. Coluria geoides образует малоцветковые синфлоресценции, представляющие собой вариант редуцированного дихазия, когда наблюдается недоразвитие одного или обоих пазушных цветков и остается только терминальный цветок.
2. В цветках C. geoides соотношение числа пыльцевых зерен и семязачатков варьирует от 2356 до 3891, что позволяет отнести его к ксеногамным видам.
3. В цветках C. geoides из-за разрыва во временном функционировании репродуктивных органов (протогиния) практически отсутствует вероятность автогамии в его цветках несмотря на полимерность андроцея и гинецея.
4. Подавляющее большинство изученных признаков семян, плодов и их элементов характеризуется низкой или средней изменчивостью, а количественные характеристики семенной продуктивности (число плодолистиков, семян и процент семенификации) демонстрируют высокий и очень высокий уровни вариабельности.
5. Выявлено влияние таких факторов, как мерность синфлоресценции и положение плода в соцветии, на некоторые показатели структурных элементов семенной продуктивности. Число плодолистиков в цветках у особей с 3-цветковыми соцветиями достоверно больше, чем в цветках у особей с 1- и 2-цветковыми синфлоресценциями. У особей с 3-цветковыми соцветиями реальная семенная продуктивность (в расчете на плод) и процент семенификации плодов, занимающих терминальное положение, выше, чем у плодов, расположенных на пазушных побегах.
Список литературы
Bolle F. 1933. Eine Übersicht über die Gattung Geum L. und die ihr nahestehenden Gattungen. – Repert. Spec. Nov. Fedde, Beih. 72: 1–119.
[Cheryomushkina, Leonova] Черемушкина В.А., Леонова Т.В. 2011. Онтогенез колюрии гравилатовидной (Coluria geoides (Pall.). – В кн.: Онтогенетический атлас растений. Т. 6. Йошкар-Ола. С. 164–170.
Cruden R.W. 1977. Pollen-ovule ratios: a conservative indicator of breeding systems in flowering plants – Evolution. 31 (1): 32–46. https://doi.org/10.1111/j.1558-5646.1977.tb00979.x
Cruden R.W. 2000. Pollen grains: why so many. – Plant Syst. Evol. 222 (1): 143−165. https://doi.org/10.1007/BF00984100
[Dutova] Дутова С.В., Неделькина Н.П., Карпова М.Р., Чумаков В.Ю., Мяделец М.А. 2015. Иммуностимулирующее действие экстракта Coluria geoides (Rosaceae) при генерализованной стафилококковой инфекции. – Российский иммунологический журнал. 9 (2): 15–17.
[Elisafenko] Елисафенко Т.В. 2008. Онтогенез и структура ценопопуляций Coluria geoides (Pall.) Ledeb. (Rosaceae) в Центральном Алтае. – Бот. журн. 93 (8): 1240–1248.
Focke W.O. 1891. Rosaceae. – In: Die Naturlichen Pflanzenfamilien. Vol. 3. P. 1–61.
[Godin, Perkova] Годин В.Н., Перкова Т.В. 2017. Биология цветения и половой полиморфизм у видов семейства Apiaceae (Московская область). – Бот. журн. 102 (1): 35–47. https://doi.org/10.1134/S0006813617010033
Gajewski W. 1959. Evolution in the Genus Geum. – Evolution. 13 (3): 378–388.
https://doi.org/10.2307/2406114
Kalkman C. 1988. The phylogeny of the Rosaceae. – Bot. J. Linn. Soc. 98 (1): 37–59. https://doi.org/10.1111/j.1095-8339.1988.tb01693.x
[Kamelin] Камелин Р.В. 1998. Материалы по истории флоры Азии (Алтайская горная страна). Барнаул. 240 с.
[Kondorskaya] Кондорская В.Р. 1989. О применении термина “дихазий”. – Биологические науки. 2: 66–71.
[Kondorskaya] Кондорская В.Р. 1990. Монохазиальные соцветия. – Биологические науки. 2: 81–92.
[Kuznetsova] Кузнецова Т.В. 1992. Морфология соцветий: современное состояние. – В кн.: Итоги науки и техники. Ботаника. М. С. 51–174.
[Kuznetsova, Timonin] Кузнецова Т.В., Тимонин А.К. 2017. Соцветие: морфология, эволюция, таксономическое значение (применение комплементарных подходов). М. 183 с.
[Kuz’mina, Ovcharenko] Кузьмина Л.В., Овчаренко С.Н. 1987. Род Coluria R.Br. – Колюрия. – В кн.: Растительные ресурсы СССР: Цветковые растения, их химический состав, использование; Семейства Hydrangeaceae–Haloragaceae. Л. С. 31–32.
[Leonova] Леонова Т.В. 2012. Семенная продуктивность Coluria geoides (Pall.) Ledeb. – В кн.: Экосистемы Центральной Азии: исследования, сохранение, рациональное использование. Кызыл. С. 70–73.
[Leonova et al.] Леонова Т.В., Черемушкина В.А., Водолазова С.В. 2010. Онтогенетическая структура популяций Coluria geoides (Rosaceae) в разных эколого-ценотических условиях в Хакасии. – Раст. ресурсы. 46 (2): 24–32.
[Mamaev] Мамаев С.А. 1973. Внутривидовая изменчивость и проблема интродукции древесных растений. – В кн.: Успехи интродукции растений. М. С. 128–148.
Müntzing A., Müntzing G. 1941. Some new results concerning apomixis, sexuality and polymorphism in Potentilla. – Botaniska Notiser. 94 (3): 237–278.
[Orlov] Орлов В.П. 1996. Колюрия гравилатовидная. – В кн.: Красная книга Республики Алтай. Редкие и находящиеся под угрозой исчезновения виды растений. Новосибирск. С. 75–76.
[Petuchova] Петухова Л.В. 2000. Гравилат городской. – В кн.: Биологическая флора Московской области. М. Вып. 14. С. 112–127.
[Polyntseva, Utemova] Полынцева Н.А., Утемова Л.Д. 1988. Колюрия гравилатовидная – Coluria geoides (Pall.) Ledeb. – В кн.: Биоэкологические особенности растений Сибири, нуждающихся в охране. Новосибирск. С. 68–74.
Potter D., Eriksson T., Evans R.C., Oh S., Smedmark J.E.E., Morgan D.R., Kerr M., Robertson K.R., Arsenault M., Dickinson T.A., Campbell C.S. 2007. Phylogeny and classification of Rosaceae. – Plant Syst. Evol. 266 (1/2): 5–43. https://doi.org/10.1007/s00606-007-0539-9
[Redkie…] Редкие и исчезающие растения Сибири. 1980. Новосибирск. 223 с.
Sokal R.R., Rohlf F.J. 2012. Biometry: the principles and practice of statistics in biological research. New York. 937 p.
[Sonnikova] Сонникова. А.Е. 2005. Колюрия гравилатовидная. – В кн.: Красная книга Красноярского края: растения и грибы. Красноярск. С. 177.
[Sonnikova] Сонникова. А.Е. 2012. Колюрия гравилатовидная. – В кн.: Красная книга Красноярского края. Т. 2: редкие и находящиеся под угрозой исчезновения виды дикорастущих растений и грибов. Красноярск. С. 294.
Troll W. 1969. Die Infloreszenzen. Bd. 2. T. 1. Jena. 630 S.
[Tichonova] Тихонова В.Л. 1974. Лапчатка прямостоящая. – В кн.: Биологическая флора Московской области. М. Вып. 1. С. 67–77.
[Vaynagiy] Вайнагий И.В. 1974. О методике изучения семенной продуктивности растений. – Бот. журн. 59 (6): 826–831.
[Vodolazova et al.] Водолазова С.В., Мяделец М.А., Карпова М.Р., Саранчина Ю.В. 2011. Антимикробная активность эфирных масел и водных извлечений из лекарственных растений Хакасии. – Сиб. мед. журн. 26 (2/2): 54–58.
[Vydrina] Выдрина С.Н. 1988. Coluria R. Br. – Колюрия. – В кн.: Флора Сибири. Rosaceae. Новосибирск. С. 89.
Weber H.E. 1995. Rosoideae. – In: Illustrierte Flora von Mitteleuropa. Blackwell Wissenschafts-Verlag, Berlin. 4 (2A). S. 269–659.
Wolf Th. 1908. Monographie der Gattung Potentilla. – Biblioth. Bot. 16 (71): 1–714.
Kalkman C. 2004. Rosaceae. – In: Flowering plants. Dicotyledons: Celastrales, Oxalidales, Rosales, Cornales, Ericales. Berlin. P. 343–386.
[Zhivotovskiy] Животовский Л.А. 1991. Популяционная биометрия. М. 271 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал