

Ботанический журнал, 2023, T. 108, № 1, стр. 51-65
МОРФОЛОГИЯ ПЫЛЬЦЫ НЕКОТОРЫХ РАСТЕНИЙ ГРЯЗЕВОГО ВУЛКАНА ГЫЗМЕЙДАН (АЗЕРБАЙДЖАНСКАЯ РЕСПУБЛИКА)
Ш. Г. Исаева 1, *, В. В. Григорьева 2, **
1 Министерство науки и образования АР Институт ботаники
Az1004 Баку, Бадамдарское шоссе, 40, Азербайджан
2 Ботанический институт им. В.Л. Комарова РАН
197022 Санкт-Петербург, ул. Профессора Попова, 2, Россия
* E-mail: isayeva.shabnam@outlook.com
** E-mail: mikhailov_val@mail.ru
Поступила в редакцию 13.09.2021
После доработки 24.11.2022
Принята к публикации 16.12.2022
- EDN: LNPLHX
- DOI: 10.31857/S0006813623010052
Аннотация
С помощью светового, сканирующего и трансмиссионного электронных микроскопов изучена морфология пыльцы 20 видов из 14 семейств двудольных растений, произрастающих на грязевом вулкане Гызмейдан (Шамахы) в Азербайджанской Республике. Существенных отклонений основных морфологических признаков пыльцы (форма, очертания, размер пыльцевых зерен, тип и число апертур, скульптура экзины, толщина и ультраструктура спородермы) не выявлено. Большинство изученных видов имеют низкий процент деформированных, недоразвитых, морфологически стерильных и атипичных пыльцевых зерен.
Азербайджанская Республика является мировым лидером по числу известных грязевых вулканов. Большинство извержений в Азербайджане и особенно в Шамахы-Гобустанском районе связано с сейсмическими событиями. При каждом крупном извержении вулканом из недр выносятся около 100–200 тыс. м3 грязевулканической брекчии, в которой установлено около 90 минералов и 30 микроэлементов. Непременными, весьма характерными микроэлементами брекчии являются: бор, ртуть, марганец, барий, стронций; щелочные металлы: литий, рубидий, цезий. Извержение вулкана может привести к рассеиванию и разбросу не только грязевулканического материала, но и других весьма нежелательных потенциальных загрязнителей воздуха и почвы. С другой стороны, деятельность грязевых вулканов сопровождается образованием ландшафтов с особым типом засоления почвы. Последние иногда пропитываются нефтью. Эти ландшафты относятся к геохимически редким и представляют интерес для изучения влияния на живые организмы избыточного содержания ряда микроэлементов (бора, молибдена, стронция), приводящего к эндемическим заболеваниям животных и т.д. Как отмечает А.Г. Ахмедов (Akhmedov, 1985), каждый грязевулканический конус представляет собой выраженную борно-молибденовую геохимическую аномалию, что проявляется не только в брекчии, но и в почвах, золе растений и животных. При этом влияние вулканической деятельности на геохимические характеристики окружающей среды особенно ощутимо проявляется в зоне полупустынь, где площади, занимаемые вулканическими ландшафтами, сравнительно велики и аридный климат способствует накоплению выносимых вулканами микроэлементов (Aliev, 2006).
Грязевой вулкан Гызмейдан располагается в 4–5 км к северо-западу от одноименного селения. Слегка выпуклое грязевулканическое поле, достигающее в поперечнике 1 км, представляет собой бугор с относительной высотой 20 м, рассеченный оврагами и промоинами. Вулкан расположен на отложениях верхнего мела и приурочен к северному крылу одноименной антиклинали. Площадь грязевулканической брекчии достигает 345 га при средней мощности 25 м (Abbasov et al., 2015). За последние 25 лет грязевой вулкан Гызмейдан, извергался в 1987 и 2011 гг. Грязевулканическое поле вулкана Гызмейдан характеризуется действующими микрофорами – крупными сопками высотой 15 м, небольшими грифонами и лужеобразными сальзами, выделяющими газ, воду и грязь. В Шамахы-Гобустанском р-не почти все грязевулканические структуры перспективно нефтегазоносны. Газы вулканов Шамахы-Гобустанского р-на характеризуются большими значениями СО2 – до 10%. Количество азота изменяется в диапазоне от 0.06 до 11.7%, инертные газы иногда достигают в среднем 0.012%, в небольшом количестве присутствует Н2 (0.006%). Локальная флора грязевого вулкана Гызмейдан насчитывает 46 видов из 21 семейства сосудистых растений. Наибольшим числом видов характеризуются семейства Asteraceae (9 видов – 19.6%) и Rosaceae (7 видов – 15%). Параллельно с комплексным изучением флоры и растительности грязевого вулкана Гызмейдан и в связи с изучением влияния грязевых вулканов на живые организмы начато изучение палиноморфологии основных видов данной территории.
В классических трудах по палиноморфологии считается, что признаки пыльцы консервативны, мало изменяются и практически не зависят от мест обитания конкретного вида, что позволяет определять таксономическую принадлежность дисперсной пыльцы и использовать палинологический метод (метод спорово-пыльцевого анализа) для решения как фундаментальных задач (уточнение вопросов систематики и филогении растений, динамики палеофлор и палеоклимата и т.д.), так и для прикладных аспектов медицины, сельского хозяйства, криминалистики и т.д. (Erdtman, 1952; Kupriyanova, Aleshina, 1972).
В связи с ростом техногенного загрязнения окружающей среды возникло новое направление палинологических исследований – палиноиндикация (Dzuba, 2006, Dzuba et al., 2006 и др.). В этих работах приводятся данные, свидетельствующие о том, что морфологические особенности пыльцевых зерен зависят от воздействия таких факторов, как пожары, радиация, повышенные концентрации ионов тяжелых металлов, пестицидов и т.д. Сторонники этого направления считают, что в неблагоприятных условиях растения продуцируют большое количество нежизнеспособной, неспособной к прорастанию и оплодотворению, уродливой (тератоморфной) пыльцы. Процент естественного полиморфизма, в зависимости от таксона, обычно не превышает 5–10%, тогда как при техногенных загрязнениях он значительно выше (45%), а может достигать и 100% (Dzuba, 2006). Тяжелые металлы могут привести к различным изменениям морфологических и физиологических особенностей пыльцевых зерен; изменяют их плоидность, меняют форму пыльцевого зерна, число апертур, симметрия меняется от симметричной на асимметричную, изменения могут затрагивать скульптуру наружной оболочки зерна (экзины). Есть предположение, что утолщение оболочки пыльцевых зерен может быть вызвано избыточным ультрафиолетовым излучением и обеспечивает таким образом лучшую защиту пыльцы от повреждения (Yeloff et al., 2008). Кроме того, изменяются функциональные физиологические процессы, обеспечивающие прорастание пыльцевой трубки (Shivanna et al., 1991; Albooghobaich, Zarinkamar, 2011). В условиях техногенного загрязнения отмечают снижение фертильности и жизнеспособности пыльцевых зерен, а также резкое уменьшение или резкое увеличение размеров пыльцевых зерен (Nilan et al., 1981). При воздействии электромагнитных излучений от высоковольтных линий ингибируются белки мембран и нарушаются процессы формирования спородермы (оболочки пыльцевого зерна). Пыльцевые зерна с очень тонкой оболочкой часто деформируются (Amjad, Shafighi, 2010). Показано, что засоление почвы увеличивает процент стерильных пыльцевых зерен, снижает жизнеспособность пыльцы и нарушает рост пыльцевых трубок (Dhingra, Varghese, 1993; Gull, Achmad, 2006).
Пыльца является частью жизненного цикла растений и изменение основных (базовых) характеристик пыльцевых зерен может существенно сказаться на репродуктивной биологии всего растения (Batygina, Vasilyeva, 1999).
В настоящее время, несмотря на актуальность, сведения о влиянии техногенных загрязнений и аномальных природных факторов на пыльцу растений остаются фрагментарными и часто противоречивыми. Пыльца растений, произрастающих в условиях, обусловленных деятельностью грязевых вулканов, до сих пор не изучалась.
Цель данного палинологического исследования – оценка влияния неблагоприятных условий, созданных деятельностью вулкана, на морфологию пыльцы.
МАТЕРИАЛ И МЕТОДЫ
Материалом для исследования послужили зрелые пыльцевые зерна 20 видов из 14 семейств двудольных растений, произрастающих на грязевом вулкане Гызмейдан. Растительный материал был собран Ш.Г. Исаевой во время 2-х полевых сезонов в рамках изучения локальной флоры данного вулкана. Для изучения пыльцы отобраны широко представленные в данном фитоценозе виды, массово цветущие в момент сбора материала, с типичными неизмененными макроморфологическими признаками.
Дополнительно для 10 видов проведен сравнительный анализ морфологии пыльцы растений, произрастающих на вулкане и в типичных местах обитания. Пыльцу для сравнения брали с гербарных образцов, хранящихся в Гербарии Ботанического института им. В.Л. Комарова РАН (LE): Archanthemis fruticulosa (M. Bieb.) Lo Presti et Oberpr. Абхазия. Рицинский заповедник. 5 VIII 1973. Максимов; Artemisia alpina Pall. ex Willd. (A. caucasica Wild.). Transcaucasia. Azerbajdzhan. 11 VI 1928. M. Sachokia, A. Grossheim; Convolvulus lineatus L. Закавказье. № 052. 28 VI 1903. И. Бородин; Euphorbia seguieriana Neck. Армения. Вайоцдзорский р-н. № 218. 20 VI 2019. М. Оганесян, И. Табаков, В. Иванова, Д. Гельтман; Filipendula vulgaris Moench. Дагестанская АССР. Буйнакский р-н. № 2134. 7 VI 1961. Н.Н. Цвелев; Hypericum linarioides Bosse. Грузинская ССР. Дманисский р-н. 25 VI 1971. Ю. Меницкий, Т. Леонова; Lappula barbata Gürke. Mortarasuni. 1837; Linum corymbulosum Rchb. Закавказье. Арешский у. 18 V 1908. Ю. Воронов; Polygala anatolica. Южно-Карабахская флористическая экспедиция 1948 г. Азербайджан, Карягинский р., между с. Туг и г. Карягино, каменистый кустарниковый склон. Собр. М.И. Кирпичников и Л.А. Смольянинова, 2 VI 1948 г. Опред. С. Тамамшян; Potentilla pedata Willd. ex Hornem. Armenia №187. A. Kechaykin.
В работе использовали методы световой (СМ), сканирующей (СЭМ) и трансмиссионной (ТЭМ) электронной микроскопии. Для светооптического исследования применяли классический ацетолизный метод Эрдтмана (Erdtman, 1952). Световые микрофотографии получены при увеличении 1000 на микроскопе Micmed-6 (LOMO, St. Petersburg, Russia) с помощью цифровой фотокамеры Canon EOS 20D.
Особенности строения поверхности пыльцевых зерен исследовали на сканирующем электронном микроскопе JEOL JSM-6390 в центре коллективного пользования Ботанического института им. В.Л. Комарова. Для исследования предварительно обработанную ацетолизной смесью пыльцу закрепляли на специальном столике с помощью двусторонней липкой ленты, затем напыляли сплав золота и палладия в вакуумной установке.
Описания проводили по общепринятой схеме. Учитывали форму и очертания пыльцевых зерен, размеры полярной оси, экваториального диаметра, толщину экзины и характер скульптуры. Измерения проводили с помощью окулярмикрометра. В каждом образце измеряли не менее 25 пыльцевых зерен. После каждого описания пыльцы помещена краткая характеристика типичных мест обитания изученного вида, составленная на основании данных, приведенных во “Флоре СССР” (Flora URSS, 1937, 1939, 1945, 1946, 1948, 1949, 1953, 1954, 1961, 1962, 1963) и “Флоре Кавказа” (Grossgeim, 1950, 1952, 1962).
Подсчет деформированных пыльцевых зерен проведен на выборках не менее 500 зерен.
Ультраструктуру оболочки пыльцевых зерен изучали на сколах с помощью сканирующего микроскопа и ультратонких срезах с помощью трансмиссионного микроскопа Hitachi-600 в центре коллективного пользования Ботанического института им. В.Л. Комарова. Для исследования с помощью ТЭМ пыльники, содержащие пыльцевые зерна, фиксировали в 2.5%-ном растворе глутарового альдегида на фосфатном буфере pH 6.8–7.2 при комнатной температуре в течение 24 часов, затем в 1%-ном растворе OsO4 на том же буфере в течение 2 часов. Материал обезвоживали ацетоном с нарастающей концентрацией и заключали в смесь эпона с аралдитом. Ультратонкие срезы изготавливали на ультратоме LCB. Срезы на сеточках контрастировали уранилацетатом (20 минут) и цитратом свинца (10 минут).
Список использованных сокращений: п. з. – пыльцевые зерна; п. о. – полярная ось пыльцевого зерна; э. д. – экваториальный диаметр пыльцевого зерна.
РЕЗУЛЬТАТЫ
Общая морфология пыльцевых зерен изученных видов по данным СМ и СЭМ
Archanthemis fruticulosa (M. Bieb.) Lo Presti et Oberpr. [Asteraceae Dumort.] (рис. 3, 1–6)
П. з. 3-бороздно-оровые, почти сфероидальные, средних размеров (табл. 1), в очертании с экватора почти округлые, с полюса почти округлые или слабо 3-лопастные. Борозды длинные, неширокие, суженные на полюсах с острыми концами. Оры овальные или округлые, большей частью плохо различимые. Скульптура шиповатая. Данные СЭМ: шипы широко-конические, крупные, более 1 мкм высотой с шероховатой поверхностью.
Таблица 1.
Количественные характеристики пыльцы растений, произрастающих на грязевом вулкане и за его пределами Table 1. Quantitative characteristics of pollen of the plants growing on the mud volcano and beyond
| Исследованный образец Studied sample |
Количественные характеристики Quantitative characteristics |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | |
| Грязевой вулкан Mud volcano |
Другое место обитания Other habitat |
|||||||||
| Asteraceae Archanthemis fruticulosa |
32.7−43.3 | 32.5−44.0 | 0.9−1 | 4.5−5.5 | <1 | 32.4−43.5 | 32.4−43.6 | 0.9−1 | 4.5−5.4 | <1 |
| Artemisia alpina | 28.5−31.4 | 30.0−34.5 | 0.9−1 | 4.0−5.0 | <1 | 26.5−29.8 | 27.2−29.8 | 0.9−1 | 4.3−4.9 | 3 |
| Convolvulaceae Convolvulus lineatus |
51.3−57.5 | 51.0−60.2 | 0.9−1 | 5.1−5.5 | 5 | 57.6−64.0 | 53.8−63.1 | 0.9−1 | 5.2−5.7 | 1 |
| Euphorbiaceae Euphorbia seguieriana |
40.0−50.0 | 41.0−50.0 | 0.8−1.1 | 4.0−4.5 | 3 | 38.9−49.7 | 40.7−49.8 | 0.9−1.1 | 4.0−4.5 | 5 |
| Rosaceae Filipendula vulgaris |
21.0−24.3 | 20.0−24.3 | 1 | 1.5−1.8 | 10 | 19.0−23.7 | 17.4−21.7 | 1−1.15 | 1.5−1.7 | 32 |
| Potentilla pedata | 22.7−26.8 | 23.5−24.8 | 0.97−1.1 | 1.8−2.0 | 56 | 23.0−25.1 | 21.6−23.0 | 1−1.2 | 1.5−1.9 | 88 |
| Boraginaceae Lappula barbata |
13.4−16.5 | 6.7−8.0 | 1.7−2.4 | 1.0 | 1 | 14.8−17.4 | 7.1−9.0 | 1.7−2.4 | 1 | 1 |
| Linaceae Linum corymbulosum |
56.8−66.0 | 59.5−69.8 | 0.8−1 | 5−5.4 | 3 | 54.8−62.0 | 54.9−59.0 | 0.9−1.0 | 4.7−5.4 | 9 |
| Clusiaceae Hypericum linarioides |
19.0−20.0 | 16.4−18.5 | 1.1−1.2 | 1.3−1.6 | 1 | 25.8−27.1 | 20.3−23.6 | 1.1−1.3 | 1.3−1.6 | 5 |
| Polygalaceae Polygala anatolica |
44.2−50.0 | 40.2−50.0 | 0.97−1.2 | 2.8−3.4 | <1 | 40.0−45.5 | 40.0−45.6 | 1 | 2.5−3.0 | 1 |
Примечание. 1 − Длина полярной оси (мкм); 2 − Длина экваториального диаметра (мкм); 3 − P/E − соотношение длины полярной оси к длине экваториального диамера; 4 − Толщина экзины (мкм) (СМ); 5 − процент деформированный пыльцевых зерен.
Note. 1 − polar axis length (P, μm); 2 − equatorial axis length (E, μm); 3 – P/E; 4 − exine thickness (μm) (LM); 5 − percentage of deformed pollen grains.
Типичными местами обитания являются сухие скалы в горном поясе.
Artemisia alpina Pall. ex Willd. [Asteraceae] (рис. 3, 8–15, 17)
П. з. 3-бороздно-оровые, почти сфероидальные, средних размеров (табл. 1), в очертании с экватора почти округлые, с полюса 3-лопастные. Борозды длинные, неширокие, суженные у полюсов, с острыми концами, оры часто нечеткие. Скульптура при светооптическом методе изучения просматривается плохо. Данные СЭМ: скульптура гранулярно-мелкошиповатая, шипы широко-конические, менее 1 мкм высотой, между шипами многочисленные округлые мелкие гранулы.
Типичными местами обитания являются степные останцовые холмы и низкогорья.
Jurinea arachnoidea Bunge [Asteraceae] (рис. 4, 1, 2)
П. з. 3-бороздно-оровые, почти сфероидальные, средних размеров или немного крупнее, п. о. 47.6–55.9 мкм, э. д. 44.0–52.0 мкм, в очертании с экватора почти округлые, с полюса почти округлые или слабо 3-лопастные. Борозды длинные, неширокие, суженные у полюсов, с острыми концами, оры не всегда ясно очерченные. Толщина экзины 7.2–7.4 мкм. Скульптура шиповатая. Данные СЭМ: шипы широко-конические, крупные, более 1 мкм высотой. Между шипами и по основаниям шипов расположены округлые довольно крупные перфорации. В препарате 1% деформированных пыльцевых зерен.
Типичными местами обитания являются ковыльные степи и остепненные каменистые склоны.
Xeranthemum cylindraceum Sm. [Asteraceae] (рис. 4, 3, 4)
П. з. 3-бороздно-оровые, почти сфероидальные, средних размеров, п. о. 38.8–43.5 мкм, э.д. 37.8–40.9 мкм, в очертании с экватора почти округлые, с полюса округлые или слабо 3-лопастные. Борозды длинные, неширокие, суженные у полюсов, с острыми концами, оры часто неотчетливые. Толщина экзины 7–7.4 мкм. Скульптура перфорировано-шиповатая, шипы широко-конические, крупные, более 1 мкм высотой. В препарате 1% деформированных п. з.
Типичными местами обитания являются каменистые, песчаные склоны холмов, остепненные луга, обочины дорог.
Lappula barbata Gürke [Boraginaceae Juss.] (рис. 2, 10–13)
Рис. 1.
Морфология пыльцы видов с грязевого вулкана Гызмейдан (Шамахы). 1−5 − Thalictrum simplex, 6−15 − Rapistrum rugosum. 1−5 − общий вид пыльцевого зерна. 6, 7, 10 − общий вид пыльцевого зерна с полюса. 8, 9, 11, 12 − общий вид пыльцевого зерна с экватора. 4, 13 − скульптура поверхности. 5, 15 − скол оболочки. 14 − ультратонкий срез оболочки.Масштабные линейки, мкм: 1, 2, 5, 6−9, 14, 15 – 1; 3, 10−12 − 5.
Fig. 1. Pollen morphology of the species from the Gyzmeidan mud volcano. 1−5 − Thalictrum simplex; 6−15 − Rapistrum rugosum. 1–3 − view of pollen grain. 6, 7, 10 − polar view of pollen grain. 8, 9, 11, 12 − equatorial view of pollen grain. 4, 13 − detail of ornamentation. 5, 15 − broken pollen wall (exine). 14 − ultrathin section of sporoderm. Scale bar, μm: 1, 2, 5, 6−9, 14, 15 – 1; 3, 10−12 − 5.

Рис. 2.
Морфология пыльцы видов с грязевого вулкана Гызмейдан (Шамахы). 1−9 – Linum corymbulosum. 10−14 – Lappula barbata. 1, 3, 6, 7, 10, 12 − общий вид пыльцевого зерна с экватора. 2, 4, 5, 11 − общий вид пыльцевого зерна с полюса. 8, 13 − скульптура поверхности. 9 − скол оболочки. 14 − ультратонкий срез оболочки. Масштабные линейки, мкм: 1−4, 8−14 – 1; 5−7 − 5.
Fig. 2. Pollen morphology of the species from the Gyzmeidan mud volcano. 1−9 – Linum corymbulosum. 10−14 – Lappula barbata. 1, 3, 6, 7, 10, 12 − equatorial view of pollen grain. 2, 4, 5, 11 − polar view of pollen grain. 8, 13 − detail of ornamentation. 9 − broken pollen wall (exine). 14 − ultrathin section of sporoderm. Scale bar, μm: 1−4, 8−14 – 1; 5−7 − 5.

П. з. 6-бороздно-оровые, гантелевидные, мелкие (табл. 1), в очертании с экватора сжато-эллиптические, с полюса округло-6-угольные. Борозды щелевидные, длинные. Оры плохо различимые, экваториально-вытянутые. При светооптическом исследовании скульптура не просматривается. Данные СЭМ: скульптура неясно морщинистая, с редкими орбикулами на поверхности.
Типичными местами обитания являются каменистые склоны, скалы. Редко встречаются в дубовых лесах.
Rapistrum rugosum (L.) All. [Brassicaceae Burnet] (рис. 1, 6–13)
П. з. 3-бороздные, почти сфероидальные или эллипсоидальные, средних размеров, п. о. 29.0–31.5 мкм, э. д. 27.5–29.7 мкм, в очертании с экватора почти округлые или эллиптические, с полюса 3-лопастные. Борозды длинные, суженные у полюсов, с острыми концами, мембрана борозд часто разрушается при ацетолизной обработке. Толщина экзины 2.5–2.7 мкм. Скульптура крупно-сетчатая, хорошо просматривается при светооптическом исследовании. Данные СЭМ: ячеи сетки глубокие, округлые или округло-угловатые в очертании, более или менее одинакового размера. Стенки ячей ровные, гладкие. В препарате 2% деформированных п. з.
Типичными местами обитания являются полынные степи, солончаки, сухие склоны, сорные места, пастбища, обочины дорог.
Helianthemum salicifolium (L.) Mill. [Cistaceae Juss.] (рис. 4, 5, 6)
Рис. 3.
Морфология пыльцы видов с грязевого вулкана Гызмейдан (Шамахы). 1−7 − Archanthemis fruticulosa; 8−18 − Artemisia alpina. 1, 2, 5, 8, 9, 15 − общий вид пыльцевого зерна с экватора. 3, 4, 14 − общий вид пыльцевого зерна с полюса. 12, 13 − атипичные пыльцевые зерна. 6, 17 − скульптура поверхности. 7, 16 − ультратонкие срезы оболочки. 18 − скол оболочки. Масштабные линейки, мкм: 1−3, 6−13, 16, 18 – 1; 4, 5, 14, 15 − 5.
Fig. 3. Pollen morphology of the species from the Gyzmeidan mud volcano. 1−7 − Archantheis fruticulosa; 8−18 − Artemisia alpina. 1, 2, 5, 8, 9, 15 − equatorial view of pollen grain. 3, 4, 14 − polar view of pollen grain. 12−13 − atypical pollen grains. 6, 17 − detail of ornamentation. 7, 16 − ultrathin section of sporoderm. 18 − broken pollen wall (exine). Scale bar, μm: 1−3, 6−13, 16, 18 – 1; 4, 5, 14, 15 − 5.
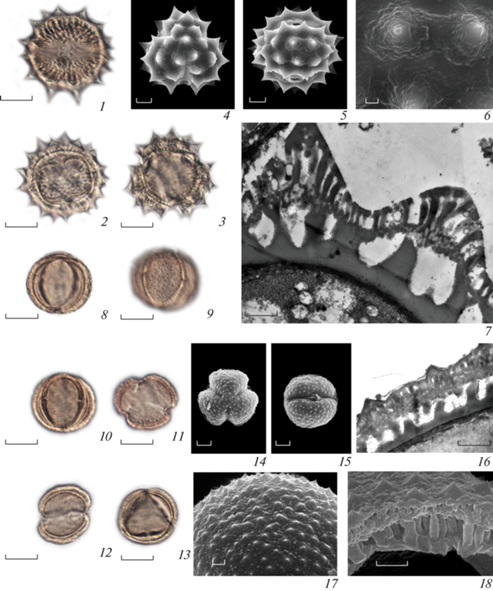
Рис. 4.
Морфология пыльцы видов с грязевого вулкана Гызмейдан (Шамахы). 1, 2 – Jurinea arachnoidea. 3, 4 – Xeranthemum cylindraceum. 5, 6 − Helianthemum salicifolium. 7, 8 − Convolvulus lineatus. 9, 10 − Euphorbia seguieriana. 11, 12 − Hedysarum sericeum. 13−14 − Hypericum linarioides. 15−16 − Scutellaria orientalis. 17−18 − Teucrium polium. 19−20 − Potentilla pedata. 21−22 − Filipendula vulgaris. 23−25 − Polygala anatolica.1, 5, 11, 13, 15, 17, 19, 21, 24 − общий вид пыльцевого зерна с экватора. 3, 7, 9, 23 − общий вид пыльцевого зерна с полюса. 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 25 − скульптура поверхности.Масштабные линейки, мкм: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 25 − 1; 13, 15, 17, 19, 21 − 5; 1, 3, 5, 7, 9, 11, 23, 24 − 10.
Fig. 4. Pollen morphology of the species from the Gyzmeidan mud volcano. 1, 2 – Jurinea arachnoidea. 3, 4 – Xeranthemum cylindraceum. 5, 6 − Helianthemum salicifolium. 7−8 − Convolvulus lineatus. 9, 10 − Euphorbia seguieriana. 11−12 − Hedysarum sericeum. 13, 14 − Hypericum linarioides. 15−16 − Scutellaria orientalis. 17−18 − Teucrium polium. 19, 20 − Potentilla pedata. 21, 22 − Filipendula vulgaris. 23−25 − Polygala anatolica.1, 5, 11, 13, 15, 17, 19, 21, 24 − equatorial view of pollen grain. 3, 7, 9, 23 − polar view of pollen grain. 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 25 − detail of ornamentation.Scale bar, μm: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 25 – 1; 13, 15, 17, 19, 21 − 5; 1, 3, 5, 7, 9, 11, 23, 24 − 10.
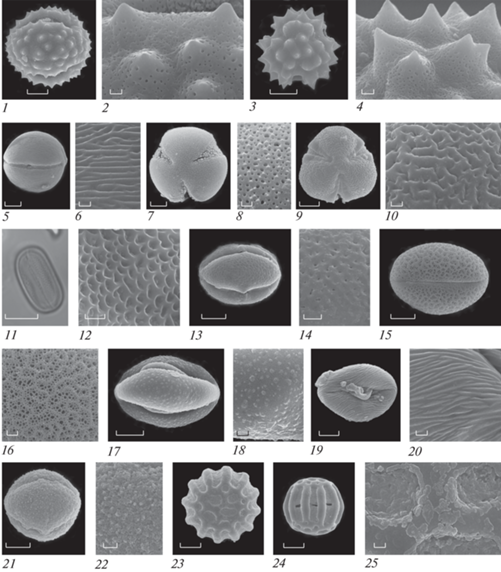
П. з. 3-бороздно-оровые, эллипсоидальные, средних размеров или немного крупнее, п. о. 44.7–52.5 мкм, э. д. 39.2–50.0 мкм, в очертании с экватора эллиптические, с полюса округло-треугольные. Борозды длинные, неширокие, оры округлые или экваториально вытянутые. Толщина экзины 2 мкм. Скульптура при светооптическом исследовании неясная. Данные СЭМ: скульптура струйчатая. Струи тонкие короткие, широко расставленные, переплетающиеся. Между струями мелкие округлые ячеи. В целом поверхность п.з. выглядит сглаженной.
Типичными местами обитания являются сухие каменистые места, горные склоны, степи.
Hypericum linarioides Bosse [Clusiaceae Lindl.] (рис. 4, 13, 14)
П. з. 3-бороздно-оровые, эллипсоидальные, среднего размера или немного меньше (табл. 1), в очертании с экватора эллиптические, с полюса 3-лопастные. Борозды неширокие, длинные. Оры экваториально вытянутые, неясно заметные. Скульптура при светооптическом исследовании неясная. Данные СЭМ: скульптура ямчато-перфорированная.
Типичными местами обитания являются открытые каменистые склоны гор, горные пастбища.
Convolvulus lineatus L. [Convolvulaceae Juss.] (рис. 4, 7, 8)
П. з. 3-бороздные, почти сфероидальные, реже эллипсоидальные, крупные (табл. 1), в очертании с экватора почти округлые или эллиптические, с полюса 3-лопастные. Борозды широкие, суженные к полюсам, с ровными краями, мембрана борозд с большим количеством неравномерно расположенных крупных гранул. Скульптура при использовании светооптического микроскопа неясная. Данные СЭМ: скульптура мелкосетчатая (почти перфорированная) с редкими очень мелкими шипиками, мембрана борозды с крупными гранулами.
Типичными местами обитания являются каменистые склоны, меловые обнажения, полупустыни, степи, сухие луга, обочины дорог, пустыри.
Euphorbia seguieriana Neck. [Euphorbiaceae Juss.] (рис. 4, 9, 10)
П. з. 3-бороздно-оровые, почти сфероидальные, средних размеров (табл. 1), в очертании с экватора почти округлые, с полюса 3-лопастные. Борозды длинные, неширокие, суженные у полюсов с острыми концами. Ора крупная, прямоугольная, экваториально вытянутая. Данные СЭМ: скульптура крупно-складчатая, по краям борозд слегка шероховатая, почти гладкая.
Типичными местами обитания являются песчаные, каменистые склоны, известняковые и меловые обнажения, ковыльно-разнотравные и кустарниковые степи. Иногда растут на выгонах и по речным галечникам.
Astragalus bungeanus Boiss. [Fabaceae Lindl.]
П. з. 3-бороздно-оровые, эллипсоидальные, в очертании с экватора округло-4-угольные, c полюса округло-треугольные или слабо 3-лопастные, средних размеров, п. о. 31.2–38.6 мкм, э. д. 16.8–20.0 мкм. Борозды длинные, узкие, оры нечеткие. Толщина экзины 1.5–1.9 мкм. Скульптура сетчатая. Данные СЭМ: ячеи неглубокие, округлые или округло-угловатые в очертании, ширина стенок меньше диаметра просветов. В препарате менее 1% деформированных п. з.
Типичными местами обитания являются сухие каменистые, песчано-каменистые, щебнистые и травяные склоны, тимьянниковые и разнотравно-злаковые степи.
Hedysarum sericeum M. Bieb. [Fabaceae] (рис. 4, 11, 12)
П. з. 3-бороздные, эллипсоидальные, в очертании с экватора округло-прямоугольные, средних размеров или немного меньше, п. о. 24.5–25.5 мкм, э. д. 15.8–20.0 мкм. Борозды длинные, узкие. Толщина экзины 1.5–1.9 мкм. Скульптура мелко сетчатая, плохо различима при использовании светооптического микроскопа. Данные СЭМ: ячеи округлые или округло-угловатые, с мелкими перфорациями или гранулами на дне, стенки ячей узкие, гладкие. В препарате около 5% деформированных п. з.
Типичными местами обитания являются выходы мела и известняка.
Medicago monspeliaca (L.) Trautv. [Fabaceae]
П. з. 3-бороздно-оровые, эллипсоидальные, в очертании с экватора округло-4-угольные, с полюса округлые или округло-треугольные, среднего размера, п. о. 35.0–36.6 мкм, э. д. 23.5–26.6 мкм. Борозды узкие, оры округлые или широкоэллиптические, слабо заметные. Скульптура при использовании светооптического микроскопа неясная. Толщина экзины 1.4–1.7 мкм. Данные СЭМ: скульптура мелкосетчатая.
Типичными местами обитания являются каменистые и щебнистые склоны, степи, обочины дорог.
Scutellaria orientalis L. [Lamiaceae Lindl.] (рис. 4, 15, 16)
П. з. 3-бороздные, почти сфероидальные или эллипсоидальные, средних размеров или немного меньше, п.о. 22.5–27.6 мкм, э.д. 21.5–24.0 мкм. В очертании с экватора почти округлые или эллиптические, с полюса 3-лопастные. Борозды длинные, неширокие. Толщина экзины 1.5–1.8 мкм. Скульптура неясно сетчатая. Данные СЭМ: скульптура дважды сетчатая. Стенки ячей первого порядка очень тонкие, ячеи крупные, округло-угловатые в очертании, неглубокие. На дне ячей первого порядка многочисленные округлые, разноразмерные ячеи второго порядка. В препарате 1% деформированных п. з.
Типичными местами обитания являются сухие травянистые места и пастбища, открытые каменистые и известняковые склоны, глинистые обрывы, щебень.
Teucrium polium L. [Lamiaceae] (рис. 4, 17, 18)
П. з. 3-бороздные, почти сфероидальные или эллипсоидальные, средних или немного меньше среднего размеров, п.о. 22.0–26.0 мкм, э.д. 22.0–25.6 мкм, в очертании с экватора почти округлые или эллиптические, с полюса 3-лопастные или округло-треугольные. Борозды длинные, неширокие, суженные у полюсов. Толщина экзины 1.5–1.9 мкм. Скульптура при светооптическом исследовании неясно сетчатая. Данные СЭМ: скульптура бородавчато-перфорированная. Бородавки довольно крупные, разноразмерные, между ними многочисленные очень мелкие перфорации. В препарате менее 1% деформированных п. з.
Типичными местами обитания являются сухие глинистые и каменистые склоны, осыпи, известняковые и меловые обнажения.
Linum corymbulosum Rchb. [Linaceae Perleb.] (рис. 2, 1–8)
П. з. 3-бороздные, почти сфероидальные, крупные (табл. 1), в очертании с экватора почти округлые, с полюса 3-лопастные. Борозды длинные, широкие, суженные у полюсов, с острыми концами. Мембрана борозд часто разрушается при ацетолизной обработке. Скульптура столбиковая. Столбики одинаковой высоты, но разного диаметра (двух типов: крупные и мелкие) плотно прилегают друг к другу. Данные СЭМ: поверхность столбиков морщинистая. Мелкие столбики с одним микроскопическим шипиком в центре, крупные с 3–4 шипиками по краям.
Типичными местами обитания являются песчаные и щебнистые поймы, равнинные участки, невысокие горы, сухие луга.
Polygala anatolica Boiss. et Heldr. [Polygalaceae Hoffmanns. et Link] (рис. 4, 23–25)
П. з. 11–13-бороздные, эллипсоидальные, с орой в виде экваториального пояска, крупные или слегка меньшего размера (табл. 1). Борозды длинные, с закругленными концами, широкие. Ширина борозд равна ширине мезокольпиума. Скульптура на полюсах сетчатая с крупными округлыми ячеями, на мезокольпиумах гладкая. Данные СЭМ: поверхность мезокольпиумов и стенок ячей почти гладкая, дно ячей и мембрана борозд шероховатые.
Типичными местами обитания являются луга, опушки, травянистые склоны.
Thalictrum simplex L. [Ranunculaceae Juss.] (рис. 1, 1–4)
П. з. многопоровые, сфероидальные, почти среднего размера, диаметр 23.0–25.1 мкм, толщина экзины 1.4–1.5 мкм, в очертании округлые. Поры округлые, без ободка, погруженные, 3.5–4 мкм в диаметре, на расстоянии 5–7 мкм друг от друга, с зернистой мембраной. Скульптура при свето-оптическом исследовании неясная. Данные СЭМ: скульптура мелко шипиковатая с редкими микроскопическими перфорациями между шипиками. Мембрана пор с крупными плотно прилегающими друг к другу бугорками, покрытыми мелкими шипиками. В препарате менее 1% деформированных п. з.
Типичными местами обитания являются разреженные леса, опушки, суходольные и пойменные луга.
Filipendula vulgaris Moench. [Rosaceae Juss.] (рис. 4, 21–22)
П. з. 3-бороздно-оровые, почти сфероидальные или широкоэллипсоидальные, средних размеров или немного меньше (табл. 1), в очертании с экватора почти округлые или широкоэллиптические, с полюса 3-лопастные. Борозды длинные, узкие, часто с неровными краями, над орой края борозд часто смыкаются. Оры плохо просматриваются. При светооптическом исследовании скульптура неясная. Данные СЭМ: скульптура шероховатая.
Типичными местами обитания являются луга, лесные опушки, поляны, степные склоны.
Potentilla pedata Willd. ex Hornem. [Rosaceae Juss.] (рис. 4, 19, 20)
П. з. 3-бороздно-оровые, почти сфероидальные или эллипсоидальные, средних размеров или немного меньше (табл. 1), в очертании с экватора почти округлые или эллиптические, с полюса округло-3-лопастные. Борозды длинные, узкие, края сближаются в области оры. Оры довольно крупные, экваториально вытянутые, овальные. Скульптура неясно струйчатая. Данные СЭМ: струи на поверхности зерна длинные, тонкие, широко расставленные.
Типичными местами обитания являются каменистые пустоши, засушливые луга, подлески, фруктовые сады.
Сравнительный анализ основных количественных характеристик пыльцевых зерен у растений, произрастающих на грязевом вулкане, и у растений из типичных мест обитания представлен в табл. 1.
Ультраструктура спородермы по данным СЭМ и ТЭМ
Для решения вопроса о влиянии последствий извержения вулкана на ультраструктуру спородермы был проведен сравнительный анализ особенностей строения оболочек пыльцевых зерен у 6 видов, произрастающих как на грязевом вулкане, так и в типичных местах обитания.
Archanthemis fruticulosa (рис. 3, 7)
По данным ТЭМ экзина тектатно-колумеллятного типа представлена сложно устроенной эктэкзиной и ламеллятной эндэкзиной. Наружный слой эктэкзины образован одним рядом широко расставленных тонких (около 0.1–0.2 мкм толщиной) столбиков, коротких (0.5–0.7 мкм высотой) между шипами и довольно длинных (1–2 мкм высотой) в шипах, которые опираются на относительно тонкий гранулярный инфратектум (не более 0.5 мкм). Столбики внутреннего ряда крупные, заметно длиннее и толще столбиков наружного ряда (0.5–0.7 мкм толщиной и до 2.5 мм высотой), часто разветвленные в верхней части, опираются на сплошной гомогенный, довольно толстый (0.5–0.7 мкм) подстилающий слой. Ламеллятная эндэкзина на срезах выглядит более электронно плотной по сравнению с эктэкзиной, по толщине равна подстилающему слою, на межапертурных участках ламеллы плотно прилегают друг к другу и эндэкзина выглядит гомогенной, в области апертур видны отдельные ламеллы.
Artemisia alpina (рис. 3, 16, 18)
По данным ТЭМ экзина тектатно-колумеллятного типа представлена сложно устроенной эктэкзиной и ламеллятной эндэкзиной. Столбики наружного слоя эктэкзины тонкие (не более 0.1 мкм толщиной и не более 0.5 мкм высотой), располагаются плотно друг к другу. Инфратектум толстый (около 1 мкм толщиной), часто этот слой выглядит гранулярным или сетчатым. Столбики внутреннего слоя высокие (около 1.5 мкм высотой), нетолстые (около 0.5 мкм толщиной), иногда раздвоенные в верхней части. На сколах оболочки, исследованных при помощи СЭМ, наружный слой столбиков выглядит губчатым, и отдельные столбики в большинстве случаев не просматриваются.
Lappula barbata (рис. 2, 14)
По данным ТЭМ экзина тектатно-коллумелятного типа, тонкая, менее 1 мкм толщиной. Все слои эктэкзины (тектум, колумеллы и подстилающий слой) имеют почти одинаковую толщину. Эндэкзина (контрастно темная на срезах) очень тонкая на межапертурных участках, около апертур сильно утолщается и по толщине становится равной эктэкзине.
Linum corymbulosum (рис. 2, 9)
По данным СЭМ экзина атектатного, колумеллятного типа представлена столбиками эктэкзины и эндэкзиной. Эктэкзина и эндэкзина почти равны по толщине. Эндэкзина гранулярного типа, гранулы плотно прилегают друг к другу.
Rapistrum rugosum (рис. 1, 14, 15)
По данным ТЭМ и СЭМ экзина тектатно-коллумелятного типа. Столбики высокие, ровные, слегка расширяются в верхней части. Головки столбиков сливаются и образуют прерывистый тектум, характерный для пыльцы с сетчатой скульптурой. Подстилающий слой толстый, по толщине равен или слегка превышает толщину тектума. Эндэкзина очень тонкая, представлена одной электронно-плотной ламеллой.
Thalictrum simplex (рис. 1, 5)
Экзина тектатно-коллумелятного типа. На сколах (СЭМ) экзина представлена толстым тектумом, низкими, короткими столбиками и подстилающим слоем. На сколах оболочки эндэкзина не просматривается.
ОБСУЖДЕНИЕ
Пыльца изученных видов относится к 5 палинотипам:
1. 3-бороздный (виды из семейств Brassicaceae, Convolvulaceae, Fabaceae (Hedysarum sericeum), Lamiaceae, Linaceae) (рис. 1, 6–15; рис. 2, 1–9; рис. 4, 7, 8, 11, 12, 15–18).
2. 3-бороздно-оровый (Asteraceae, Cistaceae, Clusiaceae, Euphorbiaceae, Fabaceae, Rosaceae) (рис. 3; рис. 4, 5, 6, 9, 10, 13, 14, 19–22).
3. 6-гетеробороздный (Boraginaceae) (рис. 2, 10–14).
4. Многобороздно-оровый (Polygalaceae) (рис. 4, 23–25)
5. Многопоровый (Ranunculaceae) (рис 1, 1–5).
Сопоставление полученных данных с материалами палинологической базы данных PALDAT и сведениями из литературных источников (Sánchez Agudo et al., 1998; Grigoryeva et al., 2015; Kluza-Wieloch et al., 2015; Humphrey, 2016; Tekin, Yilmaz, 2016; Marzouk et al., 2017; Kamel et al., 2018 и др.) показало, что пыльца всех изученных видов, произрастающих на грязевом вулкане Гызмейдан (Шамахы) обладает чертами соответствующих таксонов (семейств и родов).
Сравнительное палиноморфологическое исследование 10 видов, произрастающих на грязевом вулкане, с соответствующими видами из типичных мест обитания не выявило никаких различий в морфологии пыльцевых зерен. Это касается как качественных палинологических характеристик (тип пыльцевого зерна, форма, очертания, строение, апертур, скульптура и ультраструктура спородермы), так и количественных признаков (размеры полярной оси, экваториального диаметра, толщина экзины) (табл. 1). Выявленные незначительные различия касаются только размеров пыльцевых зерен. Однако, во всех случаях разница в размерах пыльцы исследованных видов из разных мест обитания (вулкан и другие местообитания) соответствует размаху внутривидовой изменчивости, не превышает 10 мкм и всегда остается в рамках размерной группы по классификации G. Erdtman (1952).
Признаки пыльцевых зерен считаются консервативными (Erdtman, 1952). Однако нередко, в препаратах (обычно из одного цветка, реже соцветия) наряду с типичными (значительно преобладающими) встречаются пыльцевые зерна с отличным от типичного числом и (или) расположением апертур. Обычно это единичные зерна и их количество исчисляется долями процента (Pozhidaev, 1998, 2000), хотя для некоторых таксонов характерна и более частая встречаемость атипичных по этому признаку пыльцевых зерен (Pozhidaev, 1993, 1995). В большинстве случаев такие атипичные пыльцевые зерна являются фертильными и жизнеспособными (Pire, Dematteis, 2007). Как уже отмечалось ранее, имеются данные, согласно которым техногенные загрязнения окружающей среды, в том числе повышенный уровень радиации, приводят к появлению и увеличению числа аномальных (в том числе по признаку числа и расположения апертур) пыльцевых зерен у ряда древесных и травянистых растений (Dzyuba, 2006; Dzyuba et al., 2006). Из 20 изученных видов флоры вулкана Гызмейдан (Шамахы) только у 2 представителей семейства Asteraceae (Archanthemis fruticulosa и Artemisia alpina) были обнаружены единичные атипичные зерна. У них выявлено 2 варианта таких зерен: с одной кольцевой апертурой и с 6 апертурами. Следует отметить, что такие единичные атипичные зерна встречаются как у растений, произрастающих на грязевом вулкане, так и за его пределами в типичных для видов местах обитания. Наличие атипичной пыльцы у сложноцветных – обычное явление и уже было описано ранее (Grigoryeva et al., 2015). Так, например, у некоторых полыней (Artemisia), произрастающих в благоприятных, привычных для них местах обитания количество атипичных зерен может достигать 8% (Grigoryeva et al., 2018).
Важным морфологическим показателем функционального состояния пыльцы является процент деформированных пыльцевых зерен в образце (цветке, тычинке или пыльнике). Уменьшение или полное разрушение клеточного содержимого приводит к появлению разнообразных вмятин на поверхности, нарушению формы зерна, т. е. к ее деформации. Деформированными также считают недоразвитые, очень мелкие (по сравнению с обычными) пыльцевые зерна (Shishova et al., 2019). Деформация пыльцевых зерен может быть обусловлена разными факторами, среди которых называют гибридизацию, полиплоидию, клейстогамию, апомиксис. Иногда к сильной деформации могут привести резкие нарушения условий существования вида (Kupriyanova, Aleschina, 1972). При техногенном загрязнении число деформированных пыльцевых зерен значительно возрастает и может достигать 45–100%. (Dzuba, 2006). Параллельное изучение морфологии и фертильности пыльцевых зерен в разных таксонах цветковых растений свидетельствует о том, что стерильные пыльцевые зерна, как правило, сильно деформированы или имеют очень маленькие по сравнению с обычными для вида размеры. Для большинства исследованных видов, произрастающих на грязевом вулкане Гызмейдан (Шамахы) характерен низкий процент (не более 1%) деформированной пыльцы. У Rapistrum rugosum выявлено 2% деформированных пыльцевых зерен, у Euphorbia seguieriana и Linum corymbulosum – 3%, а у Hedysarum sericeum – 5%. Сравнительный анализ пыльцы у растений с вулкана и у растений из других мест обитания показал, что процент деформированных пыльцевых зерен не зависит от места произрастания растения (табл. 1). Незначительные расхождения были отмечены у Euphorbia seguieriana, Linum corymbulosum, Hypericum linarioides. Следует отметить, что у некоторых растений, произрастающих за пределами грязевого вулкана, число деформированной пыльцы немного больше по сравнению с теми же видами из флоры вулкана (табл. 1). Самый высокий процент деформированных пыльцевых зерен был выявлен у представителей семейства Rosaceae (Filipendula vulgaris и Potentilla pedata). Так у растений Filipendula vulgaris, произрастающих на грязевом вулкане, выявлено 10% деформированной пыльцы, а у контрольных растений 32%. У Potentilla pedata 56 и 88% соответственно. Полученные данные не являются показателями влияния окружающей среды на морфологию пыльцы этих растений, поскольку для многих родов семейства Rosaceae, и в частности для рода Potentilla, всегда характерно большое количество деформированной пыльцы, что традиционно связывают с высокой степенью гибридизации представителей этого таксона.
Таким образом, в ходе проведенного палиноморфологического изучения некоторых видов флоры грязевого вулкана Гызмейдан (Шамахы) негативного влияния условий произрастания на морфологию их пыльцы не выявлено. Никаких нарушений развития основных палиноморфологических характеристик не обнаружено. С высокой степенью уверенности можно предположить, что исследованные растения, продуцируя морфологически полноценную пыльцу, имеют высокий потенциал для опыления, оплодотворения и дальнейшего семенного размножения.
Список литературы
Abbasov O.R., Mammadova A.N., Akhundov R.V., Baloglanov E.E., Huseynov A.R., Yagubov A.R. 2015. Atlas of the World Mud Volcanoes. Baku. P. 44.
[Akhmedov] Ахмедов А.Г. 1985. Грязевые вулканы и окружающая среда. 49 с.
Albooghobaich N., Zarinkamar F. 2011. Effect of lead toxicity on pollen grains in Matricaria chamomilla. – In: IPCBEE. Vol. P. 292–295.
[Aliev] Алиев Ад.А. 2006. Грязевой вулканизм южно-каспийского нефтегазоносного бассейна. – Геология и полезные ископаемые Мирового океана. 3: 35–51.
Amjad L., Shafighi M. 2010. Effect of electromagnetic fields on structure and pollen grain development in Chenopodium album L. – International Journal of Bioengineering and Life Sciences. 4 (10): 763–765.
[Batygina, Vasilyeva] Батыгина Т.Б., Васильева В.Е. 1999. Размножение растений. СПб. 230 с.
Dhingra H.R., Varghese T.M. 1993. Flowering and male reproductive functions of chick pea (Cicer arietinum L.) genotypes as affected by salinity. – Biol. Plantarum. 35 (3): 447–452.
[Dzyuba] Дзюба О.Ф. 2006. Палиноиндикация качества окружающей среды. СПб. 197 с.
Dzyuba O.F., Shrekova O.V., Tokarev P.I. 2006. On the natural polymorphism of pollen grains of Acer tataricum L. – Paleontol. J. 40 (5): 590–594.
Erdtman G. 1952. Pollen morphology and taxonomy. Angiosperms. Stockholm. 539 p.
[Flora URSS] Флора СССР. 1937, 1939, 1945, 1946, 1948, 1949, 1953, 1954, 1961, 1962, 1963. Т. 7, 8, 11–15, 19, 20, 26–28. М., Л.
[Grigoryeva et al.] Григорьева В.В., Коробков А.А., Брицкий Д.А., Михайлова Л.В. 2015. Морфология пыльцы представителей трибы Anthemideae (Asteraceae). – Бот. журн. 100 (10): 1040–1047. https://doi.org/10.1134/S000681361510004X
[Grigoryeva et al.] Григорьева В.В., Брицкий Д.А., Коробков А.А. 2018. Морфология пыльцевых зерен видов рода Artemisia (Asteraceae) Дальнего Востока России. – Бот. журн. 103 (10): 1255–1264. https://doi.org/10.7868/S0006813618100046
[Grossgeim] Гроссгейм А.А. Флора Кавказа. Издание 2-е. 1950, 1952, 1962. Т. IV, V, VI. Л.
Gull H., Achmad R. 2006. Effect of salinity on pollen viability of different canola (Brassica napus L.) cultivars as reflected by the formation of fruits and seeds. – Pak. J. Bot. 38 (2): 237–247.
Humphrey R.P. 2016. Pollen heteromorphism is pervasive in Thalictrum (Ranunculaceae). – Plant Syst. Evol. 302 (8): 1171–1177. https://doi.org/10.1007/s00606-016-1312-8
Kamel M.A., El Hadidy A.M.H., Hamed S.T., Hus-sein N.R.A. 2018. A Palynological Review for Some Species of Family Boraginaceae Juss. from the Egyptian Flora. – Annual Research and Review in Biology. 30 (3): 1–16. https://doi.org/10.9734/ARRB/2018/46408
Kluza-Wieloch M., Maciejewska-Rutkowska I., Wójtowicz W., Wiland-Szymańska J. 2015. Pollen morphology of rare species of Linum L. (Linaceae) from Poland. – Biodiv. Res. Conserv. 40: 1–12. https://doi.org/10.1515/biorc-2015-0028
[Kupriyanova, Aleshina] Куприянова Л.А., Алешина Л.А. 1972. Пыльца и споры растений флоры европейской части СССР. Т. 1. Л. 172 с.
Marzouk R.I., El-Darier S.M., Askar A.B.M. 2017. Pollen morphology of Teucrium L. (Lamiaceae, Ajugoideae) in Libya. – Bangladesh J. Plant Taxon. 24 (2): 219–226.
Nilan R.A., Rosichan J.L., Arenaz P., Hodgdon A.L., Kleinhofs A. 1981. Pollen genetic markers for detection of mutagens in the environment. – Environ. Health Perspect. 37: 19–25. https://doi.org/10.1289/ehp.813719
PalDat – Palynological Database, an online publication on recent pollen. https://www.paldat.org/
Pire S.M., Dematteis M. 2007. Pollen aperture heteromorphism in Centaurium pulchellum (Gentianaceae). – Grana. 46 (1): 1–12. https://doi.org/10.1080/00173130601101245
Pozhidaev A.E. 1993. Polymorphism of pollen in the genus Acer (Aceraceae). Isomorphism of deviant forms of Angiosperm pollen. – Grana. 32: 79–85.
Pozhidaev A.E. 1995. Pollen morphology of the genus Aesculus (Hippocastanaceae). Patterns in the variety of morphological characteristics. – Grana. 34 (2): 10–20.
Pozhidaev A.E. 1998. Hypothetical way of pollen aperture patterning. 1. Formation of 3-colpate patterns and endoaperture geometry. – Rev. Palaeobot. Palynol. 104 (1): 67–83.
Pozhidaev A.E. 2000. Hypothetical way of pollen aperture patterning. 2. Formation of polycolpate patterns and pseudoaperture geometry. – Rev. Palaeobot. Palynol. 109 (3–4): 235–254. https://doi.org//10.1016/S0034-6667(99)00057-3
Sánchez Agudo J.A., Rico E., Sánchez Sánchez J. 1998. Palynological study of Potentilla subg. Potentilla (Rosaceae) in the Western Mediterranean. – Grana. 37 (5): 276–284.10.1080/00173139809362679
Shishova M., Puzanskiy R., Gavrilova O., Kurbanniazov S., Demchenko K., Yemelyanov V., Pendinen G., Shavarda A., Gavrilenko T. 2019. Metabolic Alterations in Male-Sterile Potato as Compared to Male-Fertile. – Metabolites. 9 (2): 24. https://doi.org/10.3390/metabo9020024
Shivanna K.R., Linskens H.F., Cresti M. 1991. Pollen viability and pollen vigor. – Theoret. Appl. Genetics. 81 (1): 38–42. https://doi.org/10.1007/BF00226109
Tekin M., Yilmaz G. 2016. Palynological Studies of the Genus Convolvulus L. (Convolvulaceae) from Turkey. – Botanical Sciences. 94 (3): 543–549.
Yeloff D., Blokker P., Boelen P. Rozema J. 2008. Is Pollen Morphology of Salix polaris Affected by Enhanced UV-B Irradiation? Results from a Field Experiment in High Arctic Tundra. – Arctic, Antarctic, and Alpine Research. 40 (4): 770–774. https://doi.org/0.1657/1523-0430(07-045)[YELOFF]2.0.CO;2
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал