

Ботанический журнал, 2023, T. 108, № 1, стр. 37-50
СТРОЕНИЕ ПЫЛЬЦЕВОГО ЗЕРНА И ОПЫЛЕНИЕ У ПРЕДСТАВИТЕЛЕЙ РОДА RIBES (GROSSULARIACEAE)
Л. Ф. Яндовка 1, *, И. В. Барабанов 1
1 Российский государственный педагогический университет им. А.И. Герцена
191186 Санкт-Петербург, Наб. реки Мойки, 48, Россия
* E-mail: yandovkaTGU@mail.ru
Поступила в редакцию 29.10.2022
После доработки 11.12.2022
Принята к публикации 16.12.2022
- EDN: LOEBPP
- DOI: 10.31857/S000681362301009X
Аннотация
Изучено строение пыльцевого зерна и опыление у ряда представителей рода Ribes, которые произрастают на территории ботанических садов Ботанического института им. В.Л. Комарова РАН и Всероссийского института генетических ресурсов растений им. Н.И. Вавилова (г. Санкт-Петербург). У исследуемых растений пыльца порово-оровая. Встречаются нарушения ее морфологии, причем более всего у Ribes mandshuricum, R. ussuriense, R. alpinum, R. dikusha, R. pauciflorum. Выявлен высокий процент морфологически сформированной (фертильной) пыльцы (80.3 ± 1.9–96.4 ± 1.0%) и невысокая ее жизнеспособность при проращивании. Лучше прочих прорастает пыльца R. sanguineum, R. spicatum, R. nigrum, R. ussuriense, R. aureum (45.0 ± 2.8–29.0 ± 2.5%). Установлено, что у некоторых видов пыльца способна прорастать внутри пыльника. Выявлены опылители цветков Ribes – представители Apis, Bombus и Chrysopa oculatа. Прорастание пыльцевого зерна на рыльце пестика активное. Прохождение пыльцевой трубки до семязачатка длится 3–5 суток. В завязи развиваются 4 геми-ортотропных семязачатка в 6–10 “этажей”. Пыльца попадает на рыльце в двухклеточном состоянии, деление генеративной клетки с образованием спермиев происходит в пыльцевой трубке. Менее всего нарушений, касающихся структуры пыльцы и процессов опыления в условиях Санкт-Петербурга, у R. sanguineum, R. spicatum, R. nigrum, R. aureum.
В роде Ribes L. насчитывается 100–150 видов, обитающих в северной умеренной зоне обоих полушарий, горах Центральной Америки и Северной Африки (Firsov, Yandovka, 2018). Это листопадные, реже вечнозеленые кустарники, имеющие гладкие или колючие побеги с очередными лопастными листьями. Цветки собраны в кистях, реже пучках или одиночные. Представители рода Ribes – обоеполые, реже раздельнополые растения.
Познание закономерностей организации и функционирования генеративных органов растений, определяющих специфику процессов воспроизведения, размножения, возобновления и репродуктивную стратегию, как отдельного растения, так и популяции, является актуальным для любого семейства цветковых растений. Представители сем. Grossulariaceae (Крыжовниковые) в этом отношении не являются исключением. Важным компонентом исследования процессов репродукции является изучение механизмов и эффективности опыления. Опыление поддерживает высокий уровень гетерозиготности в популяции, определяя целостность вида. Основным структурным элементом опыления является пыльцевое зерно. Изучение его структуры и качества важно при исследовании репродукционных процессов у растений.
Систематика рода Ribes и положение отдельных видов рода является предметом дискуссий. В решении этого вопроса анализ пыльцы также может иметь значение. Изучение морфологии, фертильности и жизнеспособности пыльцевых зерен является необходимым показателем и при определении филогенетических связей в роде Ribes. Сравнительный порово-оровый анализ пыльцы, скульптуры поверхности у изученных видов позволяет определить черты сходства и различия у разных видов, и, следовательно, определить степень генетического родства сравниваемых форм.
Изучение пыльцы и процессов опыления у представителей Ribes важно и с практической точки зрения. Выявление видов, образующих пыльцу высокого качества, имеет значение в генетико-селекционной работе.
Наиболее полно пыльца смородины изучена Huang Pu-Hwa, Ye Wan-hui (Huang P.H., Ye W.-h., 1989), Ю.В. Бурменко и др. (Burmenko et al., 2009), О.А. Гавриловой и О.А. Тихоновой (Gavrilova, Tikhonova, 2013, 2016). Сведения о морфологии пыльцы отдельных представителей сем. Grossulariaceae имеются в работах Л.А Куприяновой и Л.А. Алешиной (Kupriyanova, Aleshina, 1972), A.A.M.L. Verbeek-Reuvers (Verbeek-Reuvers, 1977), Hesse et al. (Hesse et al., 2009). У видов рода Ribes жизнеспособность пыльцы определяется исследователями как высокая – подавляющее большинство пыльцевых зерен нормально развиты и характеризуются высокой оплодотворяющей способностью (Gavrilova, Tikhonova, 2016).
Процессы опыления и прорастания пыльцы в столбике пестика по крыжовниковым в литературе слабо освещены. Имеются лишь отдельные сведения об эмбриологических аспектах этих процессов у смородины. В.Ю. Мандрик и М.А. Андрусяк (Mandrik, Andrusyak, 1969a, b) изучили фазы деления ядра генеративной клетки при проращивании пыльцы на искусственной питательной среде. А.Я. Радионенко (Radionenko, 1970) рассмотрены процессы образования пыльцевой трубки возле верхушки столбика пестика и процессы вхождения пыльцевой трубки в зародышевый мешок у Ribes nigrum. А.А. Мелехина и Л.И. Якобсон (Melekhina, Yakobson, 1971) проследили ход процессов опыления и оплодотворения при скрещиваниях Ribes. В литературе отсутствует детальное описание прорастания пыльцы in vivo после ее попадания на рыльце пестика.
Перечисленные проблемы стали основанием для исследования, целью которого было определение особенностей пыльцевых зерен, опыления и прорастания пыльцы на рыльце пестика и искусственной питательной среде у видов Ribes для выявления видов, характеризующихся пыльцой максимальной жизнеспособности, пригодной для селекционной работы в условиях г. Санкт-Петербурга.
МАТЕРИАЛЫ И МЕТОДЫ
Объектами исследования были растения видов Ribes L. из коллекции открытого грунта Ботанического сада Петра Великого Ботанического института им. В.Л. Комарова РАН (Ribes sachalinense (F. Schmidt) Nakai, R. sanguineum Purch., R. rubrum L., R. mandschuricum (Maxim.) Kom., R. spicatum E. Robson, R. saxatile Pall., R. aureum Pursh., R. alpinum L., R. nigrum L.). Их характеристика приводится в статье Г.А. Фирсова и Л.Ф. Яндовка (Firsov, Yandovka, 2018). Использовались также виды, произрастающие в питомнике Всероссийского института генетических ресурсов растений им. Н.И. Вавилова: Ribes dikuscha Fisch. ex Turcz., R. hudsonianum Rich., R. nigrum ssp. europeum Zanez, R. nigrum ssp. sibiricum (Wolf.), R. pauciflorum Turcz. ex Pojark., R. usuriense Jancz ex M. Vilm.
Все исследуемые растения – многолетние листопадные кустарники, стелющиеся или крупные прямостоячие. Цветки обоеполые (у Ribes alpinum раздельнополые цветки), собраны в кистевидные соцветия.
Для определения фертильности и жизнеспособности пыльцы использовали свежую пыльцу (срок хранения не более 10 суток). Фертильность пыльцы оценивали окрашиванием ацетокармином, жизнеспособность – проращиванием на искусственной питательной среде (Romanova et al., 1988). Для изучения морфологии пыльцевых зерен их обрабатывали с использованием ацетолизного метода (Erdtman, 1952). Затем просматривали пыльцу на сканирующем электронном микроскопе Zeiss EVO-40 в Центре коллективного пользования “Атомно-силовой и электронной микроскопии” РГПУ им. А.И. Герцена. Прохождение пыльцевой трубки в столбике пестика изучали, приготовив анатомические срезы, которые просматривали также на сканирующем электронном микроскопе.
РЕЗУЛЬТАТЫ
У исследуемых растений Ribes нормально развитые пыльцевые зерна сферической, реже эллипсовидной формы, с вогнутыми латеральными плоскостями. Пыльца является порово-оровой. Она характеризуется сложными апертурами. У такой пыльцы в поре видна экваториально вытянутая ора. Ора образована в результате утончения или полного отсутствия в поре слоев экзины. Обычно ора имеет линзовидные очертания (Punt et al., 2007). По диаметру ора больше ширины экзоапертуры (поры), поэтому как бы прикрывает пору (рис. 1g), при этом очертания эндопоры и эктопоры не соответствуют друг другу. У изученных растений порово-оровая пыльца в очертании округлая (рис. 1a) или округло-угловатая (рис. 1b). У смородин между двумя обозначенными типами пыльцевых зерен имеются переходные формы, выражающиеся, как правило, в аномальном расположении оры и ее размерах. Плоскости пыльцевого зерна имеют околооровые зоны (рис. 1c), различающиеся у разных видов по форме и размерам. Оры в околооровой зоне располагаются или в ее центре, или смещены к периферии зоны (рис. 1f, g). Околооровые зоны у ряда видов хорошо обособленные, но в некоторых случаях они способны сливаться между собой (рис. 1d). Структура околооровых зон у большинства исследуемых видов мелкозернистая, однако, у R. usuriense она гладкая, а у R. sanguineum, R. rubrum и R. saxatile крупнозернистая. У изученных видов оры выпуклые (рис. 1a–h), прикрывают пору, располагаются по 1 или 2 (реже, рис. 1f) на одну зону. Поверхность оры может быть ровной (рис. 1b, e, h) или мелкобугорчатой (рис. 1c, d, i).
Рис. 1.
Пыльца у представителей Ribes. a – Ribes alpinum; b – R. hudsonianum; c – R. saxatile; d – R. aureum; e – R. hudsonianum; f, g – R. rubrum; h – R. sanguineum; i – R. saxatile; j – R. ussuriense; k – R. mandshuricum; l – R. nigrum. c nc z – слияние околооровых зон; s p g – сморщивание пыльцевого зерна; b p – граница поры под орой; c – ора; nc z – околооровая зона; l p – крупная пыльца; m p – пыльца средних размеров; a c – отсутствие оры. Fig. 1. Pollen of the representatives of Ribes.a – Ribes alpinum; b – R. hudsonianum; c – R. saxatile; d – R. aureum; e – R. hudsonianum; f, g – R. rubrum; h – R. sanguineum; i – R. saxatile; j – R. ussuriense; k – R. mandshuricum; l – R. nigrum. c nc z – confluence of the zones surrounding colpi; s p g – shrinkage of pollen grains; b p – the pore boundary under the colpus; c – colpus; nc z – zone around colpus; l p – large pollen; m p – medium-sized pollen; a c – absence of colpus.
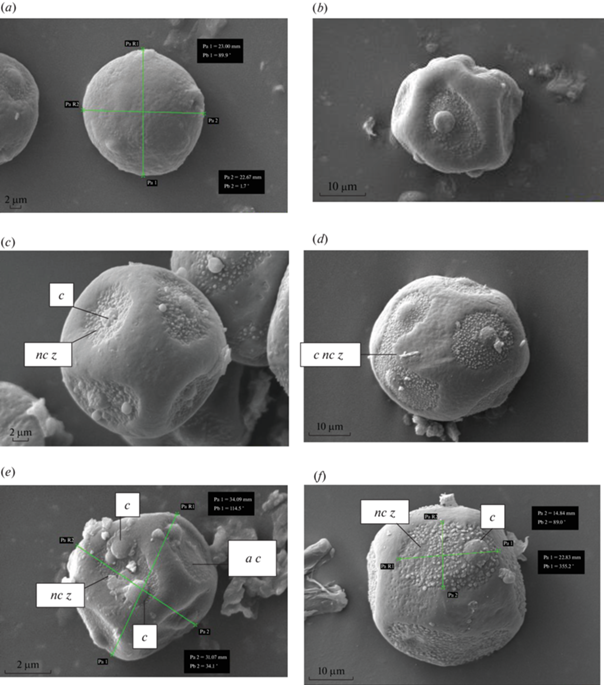
Рис. 1.
Окончание.
Скульптура поверхности пыльцевого зерна различная – от гладкой до гранулярной, мелкосетчатой либо струйчатой.
Краткая характеристика пыльцевых зерен у видов Ribes.
Ribes alpinum. Пыльцевое зерно шарообразное, диаметром около 22 мкм. Зоны вокруг оры обособленные, округлые, со слабо зернистой поверхностью. Таких зон 5–6 на одно зерно, в центре содержат по одной выпуклой оре с невыраженной структурой. Скульптура поверхности пыльцы слабо морщинистая.
Ribes aureum. Пыльцевое зерно шаровидное, диаметр достигает 36 мкм. Околооровых зон 6–8 на одно пыльцевое зерно, встречаются обособленные зоны округлой формы (диаметр от 13 до 17 мкм) и сливающиеся зоны, приобретающие подковообразную форму. Скульптура поверхности пыльцевого зерна напоминает таковую у R. alpinum – слабо морщинистая, поверхность зон вокруг оры с ярко выраженной зернистостью. Оры выпуклые, расположены в центре зон, характеризуются зернистой поверхностью.
Ribes dikusha. Пыльцевое зерно шаровидное, диаметром до 29 мкм, несет от 5 до 7 околооровых зон округлой формы, не имеющих тенденции к слиянию. Поверхность пыльцевого зерна гладкая, скульптура поверхности околооровых зон слабо зернистая. Околооровые зоны диаметром около 15 мкм, в их центре находится сильно выпуклая крупная ора (5–6 мкм в диаметре).
Ribes hudsonianum. Пыльцевое зерно неправильной формы, но приближающейся к шаровидной форме, с ярко выраженными вогнутыми плоскостями. Диаметр пыльцевого зерна около 27 мкм. Зоны вокруг оры эллипсоидной формы (около 15 мкм в длину и 11 мкм в ширину), 7–9 на пыльцевое зерно. Структура поверхности пыльцевого зерна гладкая, поверхность зоны вокруг оры зернистая. Оры выпуклые, с невыраженной структурой, располагаются в центре околооровых зон, однако некоторые зоны лишены ор и пор.
Ribes mandshuricum. Пыльцевое зерно шарообразное, до 30 мкм в диаметре, поверхность морщинистая либо слабо морщинистая. Пыльцевые зерна несут 4–5 крупных околооровых зон с выраженной зернистой скульптурой, имеющих тенденцию к слиянию. Оры слабо выпуклые, их структура не выражена, расположены ближе к периферии зоны. Часть зон лишена оры.
Ribes nigrum. Пыльцевые зерна неправильной формы, оры (5–6 на пыльцевое зерно) располагаются на вогнутых гранях, некрупные (12–14 мкм в длину и 8–10 мкм в ширину), околооровые зоны имеют тенденцию к слиянию. Диаметр зерна до 30 мкм. Скульптура поверхности пыльцевого зерна слабоморщинистая, оровых зон – выраженно зернистая. Оры расположены центрально, имеют правильную округлую форму, выпуклые.
Ribes pauciflorum. Пыльцевое зерно имеет форму, приближающуюся к шаровидной, грани вогнутые, несут некрупные (около 10 мкм в диаметре) округлые околооровые зоны в количестве 5 или 6, имеющие тенденцию к слиянию и образованию зон подковообразной формы. Поверхность пыльцевого зерна выемчатая, зоны вокруг оры имеют слабо зернистую структуру, расположены центрально. Диаметр пыльцевого зерна около 29 мкм.
Ribes rubrum. Пыльцевое зерно крупное (диаметр 36 мкм), шарообразное, с гладкой поверхностью. Несет 6 крупных (до 22 мкм в длину и 14 мкм в ширину) эллипсоидных околооровых зон с выраженной крупнозернистой структурой. Оры расположены ближе к периферии зоны, выпуклые, не имеют выраженной структуры.
Ribes sanguineum. Пыльцевое зерно широкояйцевидной формы, его длина 33 мкм, ширина 29 мкм. Околооровых зон 3–4 на зерно. Скульптура поверхности пыльцевого зерна струйчатая.
Ribes saxatile. Пыльцевое зерно некрупное (диаметр до 22 мкм), шаровидное, несет до 8 маленьких (~8 мкм в диаметре), округлых околооровых зон, не сливающихся между собой, крупнозернистой структуры. Оры выпуклые, расположены в зоне центрально, их структура не отличается от общей структуры зоны. Поверхность пыльцевого зерна не имеет выраженных скульптурных особенностей, гладкая.
Ribes ussuriense. Форма пыльцевого зерна неправильная, приближается к кубической форме, с выраженными вогнутыми гранями. Диаметр пыльцевого зерна 30–31 мкм. Оры (5–7 на одно зерно) выпуклые, расположены центрально, либо смещены к периферии. Скульптура поверхности пыльцевого зерна, околооровых зон и ор гладкая (рис. 1a–l).
Эти характеристики согласуются со сведениями О.А. Гавриловой и О.А. Тихоновой (Gavrilova, Tikhonova, 2013) о морфологии пыльцы видов сем. Grossulariaceae.
Анализируя полученные данные, среди изученных видов можно выделить две группы, различающиеся по морфологии пыльцевого зерна:
I. Ribes hudsonianum, R. nigrum, R. pauciflorum, R. ussuriense – пыльцевое зерно неправильной формы, с явно выраженными вогнутыми латеральными плоскостями.
II. Ribes aureum, R. mandshuricum, R. dikusha, R. saxatile, R. spicatum, R. rubrum, R. sanguineum – пыльцевое зерно шарообразной формы, без вогнутых плоскостей, зоны вокруг оры часто сливаются.
Предполагается, что виды, принадлежащие к одной группе, характеризуются большей степенью родства по сравнению с видами из разных групп.
Пыльца смородины характеризуется широким полиморфизмом не только между разными видами, но и внутри одного вида. Иную скульптуру поверхности пыльцы у некоторых пыльцевых зерен от типичной для вида скульптуры следует рассматривать как аномальность. Нарушения морфологической структуры пыльцевых зерен разнообразные. Деформации пыльцы, встречающиеся чаще всего, проявляются в значительном (рис. 1h) или небольшом сжатии клеточного содержимого (рис. 1l). Значительно сжатая пыльца не окрашивается ацетокармином, что свидетельствует о ее морфологической несформированности; такая пыльца не прорастает на искусственной питательной среде. Как нарушение морфологии пыльцы встречаются также: наличие одной оры в апертурной области (околооровой зоне) вместо двух ор (рис. 1e), отсутствие симметрии в расположении пор и ор, неравномерное распределение по пыльцевому зерну ор, различие размеров околооровых зон в пределах одного пыльцевого зерна (рис. 1j). У всех видов встречается слишком крупная пыльца (рис. 1k). Чрезмерно крупная пыльца (более 50 мкм) у видов Ribes почти вся деформированная, что выражается в ее вытягивании вдоль полярной оси, сжатии, в редких случаях чрезмерной бугристости. Такая пыльца обычно не окрашивается ацетокармином (не является фертильной) и не прорастает на искусственной питательной среде. Как видно из таблицы 1, более всего деформированной пыльцы у R. mandshuricum (12.7%), R. ussuriense (10.4%), R. alpinum (10.0%), R. dikusha (6.9%) и R. pauciflorum (6.7%). У всех видов встречается также слишком мелкая пыльца (менее 20 мкм), которая обычно, как и крупная пыльца, деформированная и не способная к оплодотворению. При проращивании на искусственной питательной среде она либо не прорастает, либо дает пыльцевые трубки, быстро останавливающиеся в росте (короткие). Больше всего мелкой пыльцы у R. rubrum (15.8%) и R. alpinum (13.6%).
Таблица 1.
Фертильность пыльцевых зерен видов Ribes, определяемая окрашиванием ацетокармином Table 1. Fertility of pollen grains of Ribes species as determined by acetocarmine staining
| Вид Species |
Пыльцевые зерна/Pollen grains | |||||||
|---|---|---|---|---|---|---|---|---|
| всего total |
с измененной формой of modified form |
окрашенные stained |
крупные large |
средние medium |
мелкие small |
|||
| шт./pcs. | шт./pcs. | % | шт./pcs. | % ± m | % | |||
| Ribes sanguineum | 314 | 1 | 0.3 | 280 | 89.2 ± 1.7 | 1.9 | 92.7 | 5.4 |
| R. aureum | 606 | 3 | 0.5 | 529 | 87.3 ± 1.3 | 2.0 | 94.6 | 3.4 |
| R. spicatum | 549 | 14 | 2.6 | 480 | 87.4 ± 1.4 | 3.3 | 91.3 | 5.4 |
| R. mandschuricum | 427 | 54 | 12.7 | 363 | 85.1 ± 1.7 | 2.6 | 80.7 | 3.0 |
| R. alpinum | 396 | 36 | 10.0 | 318 | 80.3 ± 1.9 | 4.5 | 81.8 | 13.6 |
| R. nigrum | 286 | 6 | 2.1 | 268 | 93.7 ± 1.4 | 6.3 | 86.7 | 7.0 |
| R. hudsonianum | 300 | 11 | 3.6 | 289 | 96.4 ± 1.0 | 7.8 | 89.2 | 3.0 |
| R. dikusha | 220 | 15 | 6.9 | 203 | 92.2 ± 1.8 | 5.2 | 90.5 | 4.3 |
| R. ussuriense | 280 | 23 | 10.4 | 251 | 89.6 ± 1.8 | 5.4 | 92.6 | 2.0 |
| R. pauciflorum | 300 | 20 | 6.7 | 265 | 88.2 ± 1.9 | 3.2 | 87.4 | 9.4 |
| R. saxatile | 324 | 16 | 5.0 | 284 | 87.7 ± 1.7 | 1.0 | 91.8 | 7.2 |
| R. rubrum | 260 | 3 | 1.2 | 234 | 90.0 ± 0.6 | 4.3 | 79.9 | 15.8 |
Наиболее пригодной для оплодотворения считается пыльца средних размеров (21–49 мкм; рис. 1k). Больше всего пыльцы средних размеров у R. saxatile (91.8%), R. sanguineum (92.7%), R. ussuriense (92.6%) и R. aureum (94.6%) (табл. 1).
Нормально сформированная (фертильная) пыльца окрашивается ацетокармином. Как видно из таблицы 1, изученные растения имеют высокий процент окрашивания пыльцы: 80.3 ± 1.9% (R. alpinum), 85.1 ± 1.7% (R. mandschuricum), 87.3 ± ± 1.3%–87.7 ± 1.7% (R. aureum, R. spicatum, R. saxatile), 88.2 ± 1.9% (R. pauciflorum), 89.2 ± 1.7 (R. sanguineum), 89.6 ± 1.8% (R. ussuriense), 90.0 ± ± 0.6% (R. rubrum), 92.2 ± 0.8% (R. dikusha), 93.7 ± ± 1.4% (R. nigrum), 96.4% (R. hudsonianum).
Несмотря на то что пыльца смородины в большинстве своем нормально сформирована, прорастание ее на искусственной питательной среде демонстрирует довольно низкие результаты. Наиболее ценными для оплодотворения считаются пыльцевые зерна, образующие пыльцевые трубки средней длины (5–6 диаметров пыльцевого зерна); короткие пыльцевые трубки не способны обеспечить доставку спермиев к зародышевому мешку, а длинные трубки, как показали наблюдения, часто прорастают с аномалиями – изгибаются, закручиваются в столбике пестика и также часто не способны дорасти до микропиле. Процент проросших на искусственной питательной среде пыльцевых зерен у изученных видов показан в таблице 2. Лучше всего прорастает пыльца R. sanguineum (45.0 ± 2.8%), R. spicatum (41.0 ± 2.4%), R. nigrum (36.1 ± 2.6%), R. aureum (29.0 ± 2.5%). Наибольшее количество пыльцевых зерен, проросших пыльцевыми трубками средней длины, наблюдается у видов R. sanguineum (60.6%), R. spicatum (51.2%), R. nigrum (47.8%).
Таблица 2.
Жизнеспособность пыльцевых зерен Ribes, определяемая проращиванием на искусственной питательной среде Table 2. Viability of Ribes pollen grains as determined by germination on an artificial nutrient medium
| Вид Species |
Всего просмотрено пыльцевых зерен, шт. Number of examined pollen grains |
Пыльцевых зерен, из них проросших пыльцевыми трубками Share of germinated pollen grains; of them germinated with pollen tubes |
|||
|---|---|---|---|---|---|
| всего, % ± m total, % ± m |
длинными, % long, % |
средней длины, % medium, % |
короткими, % short, % |
||
| Ribes aureum | 320 | 29.0 ± 2.5 | 3.2 | 27.1 | 69.5 |
| R. alpinum | 230 | 16.0 ± 2.4 | 0 | 32.4 | 67.5 |
| R. hudsonianum | 256 | 21.9 ± 2.6 | 10.7 | 32.1 | 57.1 |
| R. mandshuricum | 295 | 11.8 ± 1.8 | 0 | 0 | 100.0 |
| R. dikuscha | 200 | 19.0 ± 2.7 | 0 | 5.0 | 94.7 |
| R. nigrum | 324 | 36.1 ± 2.6 | 33.3 | 47.8 | 18.8 |
| R. sanguineum | 300 | 45.0 ± 2.8 | 17.9 | 60.6 | 36.7 |
| R. saxatile | 182 | 14.8 ± 2.6 | 0 | 0 | 100.0 |
| R. spicatum | 200 | 41.0 ± 2.4 | 27.0 | 51.2 | 31.0 |
| R. usuriense | 200 | 17.0 ± 2.6 | 0 | 26.5 | 73.5 |
У части видов Ribes пыльца способна прорастать, находясь еще внутри пыльника. Проросшая внутри пыльника пыльца в основном образует неправильно ориентированные пыльцевые трубки. Часто они изгибаются, быстро останавливаются в росте и лопаются, образуя на конце булавовидные утолщения (рис. 2a–c). В таблице 3 приведены виды, у которых в пыльнике наблюдали прорастание пыльцы. Обычно прорастание неполное, однако у некоторых видов (R. mandschuricum, R. sachalinense, R. spicatum) пыльцевые зерна прорастали как пыльцевыми трубками средней длины, способными к оплодотворению, так и короткими пыльцевыми трубками (табл. 3).
Рис. 2.
Прорастание пыльцы внутри пыльника у Ribes spicatum. a–c – прорастание пыльцы внутри пыльника; d – части соцветия (b i – основание соцветия, e i – окончание соцветия, c z i – центральная зона соцветия); s p t – остановка роста пыльцевой трубки; c p t – изогнутые пыльцевые трубки.
Fig. 2. Pollen germination inside the anther in Ribes spicatum. a–c – pollen germination inside the anther; d – parts of the inflorescence; (b i – the base of the inflorescence, e i – the end of the inflorescence, c z i – the central zone of the inflorescence); s p t – stop of pollen tube growth; c p t – curved pollen tubes.
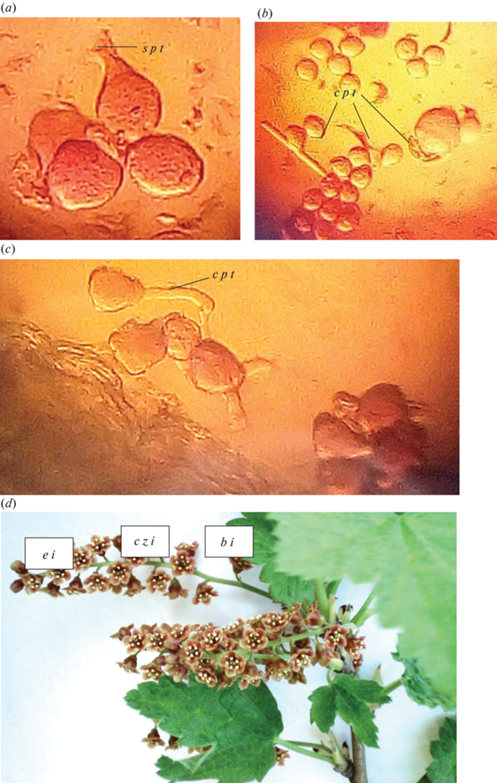
Таблица 3.
Прорастание пыльцевых зерен внутри пыльника у некоторых видов Ribes (окрашивание ацетокармином) Table 3. Germination of pollen grains of some Ribes species inside the anther (acetocarmine staining)
| Вид Species |
Зона соцветия Inflorescence zone |
Всего просмот-
рено пыльцевых
зерен, шт. Number of exami- ned pollen grains |
Пыльцевых зерен, из них проросших пыльцевыми трубками Share of germinated pollen grains; of them germinated with pollen tubes |
|||
|---|---|---|---|---|---|---|
| всего, % ± m total, % ± m |
длинными, % long, % |
средней длины, % medium, % |
короткими, % short, % |
|||
| Ribes spicatum | ЦЗ/c z i | 41 | 34/82.9 | 32.4 | 50.0 | 17.6 |
| ОкС/e i | 79 | 3/3.8 | 0 | 100.0 | 0 | |
| R. sachalinense | ЦЗ/c z i | 75 | 21/28.0 | 0 | 71.4 | 28.6 |
| ОкС/e i | 55 | 2/3.6 | 0 | 100.0 | 0 | |
| R. sanguineum | ОС/ b i | 91 | 7/7.7 | 0 | 100.0 | 0 |
| ЦЗ/c z i | 33 | 24/72.7 | 0 | 100.0 | 0 | |
| ОкС/e i | 118 | 59/50.0 | 0 | 100.0 | 0 | |
| R. mandschuricum | ОС/ b i | 71 | 1/1.4 | 0 | 100.0 | 0 |
| ЦЗ/c z i | 104 | 22/21.2 | 0 | 100.0 | 0 | |
| ОкС/e i | 179 | 1/0.6 | 0 | 0 | 100.0 | |
| R. aureum | ОС/ b i | 95 | 15/15.8 | 46.7 | 53.3 | 0 |
| ЦЗ/c z i | 98 | 15/15.3 | 0 | 100.0 | 0 | |
| ОкС/e i | 90 | 10/11.1 | 0 | 100.0 | 0 | |
Установлено, что процент прорастания пыльцы внутри пыльника неодинаковый в цветках, взятых из разных участков соцветия. В связи с этим условно соцветия (кисти) были разделены на три зоны: основание соцветия, центральная часть и окончание соцветия (рис. 2d). Из каждой зоны отдельно собирали пыльники и просматривали их под микроскопом. Полученные результаты представлены в таблице 3, из которой видно, что почти у всех видов (исключение – R. sanguineum) наилучшее прорастание внутри пыльника (образование пыльцевых трубок средней длины) имеется у пыльцы из цветков центральной зоны и в ряде случаев – основания соцветия.
Высокая доступность генеративных структур цветка смородины и расположение их в хорошо заметных соцветиях делает цветки Ribes доступными для многих видов насекомых-опылителей, поэтому в опылении четкой специализации не замечено. Выявлено, что среди опылителей цветков Ribes в условиях Санкт-Петербурга представители родов Apis, Bombus и златоглазки Chrysopa oculatа. Представители отрядов Coleoptera, Diptera и Hemiptera периодически переползают по растению. Насекомые активно посещают цветки в светлое время суток, при этом наблюдаются 2 пика активности: в утренние (между 10 и 11) и вечерние (между 17 и 18) часы.
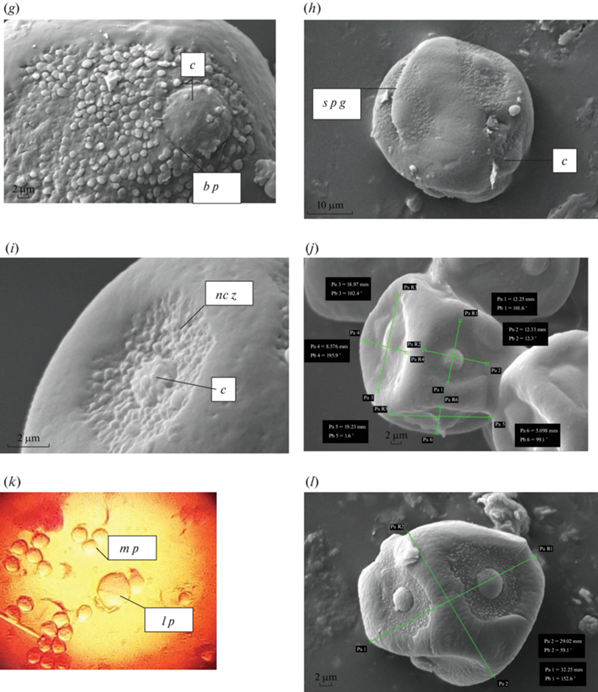
Поверхность рыльца пестика у смородины сильно выемчатая, поэтому пыльца, попадающая на него, прочно задерживается (рис. 3c, d). После попадания пыльцевого зерна на рыльце пестика даже при невысоких положительных температурах воздуха почти все находящиеся на рыльце пыльцевые зерна начинают прорастать (рис. 3d). При этом пыльцевое зерно может провалиться сразу в канал столбика пестика, в этом случае оно также способно к прорастанию (рис. 3m, n). Согласно А.С. Афанасьевой (Afanas’eva, 1964), для прохождения пыльцевой трубки в столбике пестика до завязи растениям Ribes требуется 96–120 часов после распускания цветка. А.Ф. Попова (Popova, 1965) отмечает, что слияние гамет в семязачатке происходит через 2 суток. Проведенное нами исследование позволяет лишь отчасти согласиться с авторами. Северо-Западный регион, где проводили исследование, отличается возможностью резкого изменения температуры среды, даже в течение суток. При относительно высокой температуре воздуха прохождение пыльцевой трубки до семязачатка происходит в среднем за 3–5 суток, что согласуется с данными А.А. Радионенко (Radionenko, 1970). При понижении температуры рост пыльцевой трубки в тканях столбика может задержаться до 10–12 дней, или вообще остановиться, или остановиться на некоторое время (до 3 суток), после чего пыльцевая трубка дегенерирует.
Рис. 3.
Прорастание пыльцы в столбике пестика у видов Ribes. a – Ribes sachalinense (срез завязи пестика); b – R. alpinum; c, d, e – R. sachalinense (c, d – рыльце пестика); f, g – R. aureum; h, i, j – R. sachalinense; k – R. nigrum; l – R. sachalinense; m, n – R. sanguineum (пыльцевые зерна в канале столбика пестика). o p g – нормальное пыльцевое зерно; f p g – сплющенное пыльцевое зерно; p t – пыльцевая трубка; c s p – канал столбика пестика; o – семязачаток; g t – железистые трихомы на поверхности пестика; s p t – сморщенные пыльцевые трубки, изменившие направление роста в канале столбика пестика; c d g – изменение направления роста пыльцевой трубки; t p t – скручивания пыльцевых трубок между собой; s p – столбик пестика; p g – пыльцевое зерно. Fig. 3. Pollen germination within the pistil style in the Ribes species.a – Ribes sachalinense (ovary section); b – R. alpinum; c, d, e – R. sachalinense (c, d – stigma); f, g – R. aureum; h, i, j – R. sachalinense; k – R. nigrum; l – R. sachalinense; m, n – R. sanguineum (pollen grains in the canal of the pistil style).o p g – normal pollen grain; f p g – flattened pollen grain; p t – pollen tube; c s p – channel of the pistil style; o – ovule; g t – glandular trichomes on the pistil surface; s p t – shrinked pollen tubes which changed the direction of growth in the canal of the pistil style; c d g – change in the direction of growth of the pollen tube; t p t – intertwining pollen tubes; s p – pistil style; p g – pollen grain.

Рис. 3.
Окончание.

В семязачатках уже в первый день цветения наблюдаются зрелые зародышевые мешки, готовые к оплодотворению. Согласно В.Ю. Мандрик и М.А. Андрусяк (Mandrik, Andrusyak, 1969b), моноспорический зародышевый мешок у Ribes восьмиядерный и в зрелом состоянии состоит из яйцевого аппарата из двух синергид и яйцеклетки, центральной клетки, трех антипод. В каждом гнезде завязи развиваются по 4 геми-ортотропных семязачатка гинецея, согласно представлениям И.И. Шамрова (Shamrov, 2017), расположенных в 6–10 “этажей” (рис. 3a).
По литературным данным (Popova, 1965; Mandrik, Andrusyak, 1969a; Radionenko, 1970; Kobasnidze, 1983) и нашим наблюдениям, пыльца попадает на рыльце в двухклеточном состоянии. Деление генеративной клетки на два спермия происходит в пыльцевой трубке во время роста пыльцевой трубки. Вегетативное ядро движется вместе с ними. Э.Я. Кобаснидзе (Kobasnidze, 1983) как у смородины, так и у крыжовника, наблюдала вхождение в зародышевый мешок сразу до трех пыльцевых трубок. Несмотря на присутствие добавочных пыльцевых трубок и спермиев в зародышевом мешке, диспермия автором не наблюдалась.
Попав в полость завязи, пыльцевая трубка продолжает активный рост, пока не дорастет до микропиле семязачатка. Поверхность пыльцевой трубки у представителей Ribes мелкобугорчатая, диаметр пыльцевой трубки 12–17 мкм, ее окончание имеет вздутия (рис. 3d–h). По мере прохождения спермиев в пыльцевой трубке к области микропиле, она из цилиндрической формы приобретает сплющенную форму (рис. 3b). Следует отметить, что мелкие пыльцевые зерна, попав на рыльце пестика, часто даже не начинают роста, быстро сморщиваются и погибают. Часть пыльцы может начать рост через 2–3 дня после попадания на рыльце. Пыльцевые зерна, которые долго не прорастают, сморщиваются (рис. 3c). Многие нормально сформированные пыльцевые зерна, начав прорастать в столбике пестика, быстро останавливаются, образуя лишь короткие пыльцевые трубки и затем дегенерируют. Другая часть пыльцы во время роста в канале столбика пестика проходит дальше, затем пыльцевые трубки могут остановиться на разной длине столбика, или пыльцевые трубки продолжают активный рост, но при этом изгибаются, складываются “в гармошку” (рис. 3b, e), скручиваются между собой (рис. 3j). Впоследствии такие пыльцевые трубки обычно дегенерируют, но иногда дорастают до микропиле семязачатка. У всех видов выявлены случаи, когда пыльца, попав не на рыльце, а на стенку столбика пестика, прорастала в его канале сбоку (рис. 3k, l).
ЗАКЛЮЧЕНИЕ
У исследуемых растений Ribes нормально развитые пыльцевые зерна сферической или эллипсовидной формы, с вогнутыми латеральными плоскостями, порово-оровые. Плоскости пыльцевого зерна имеют околооровые зоны. Оры в околооровой зоне располагаются или в ее центре, или смещены к периферии зоны. Околооровые зоны хорошо обособленные (чаще) или сливаются между собой.
Пыльца смородины характеризуется широким полиморфизмом размеров и формы, как между разными видами, так и в пределах одного вида. У всех изученных видов встречаются нарушения морфологической структуры пыльцевых зерен; такая пыльца не прорастает на искусственной питательной среде. Более всего деформированной пыльцы у Ribes mandshuricum, R. ussuriense, R. alpinum, R. dikusha и R. pauciflorum (12.7–6.7%). Наиболее пригодной для оплодотворения является пыльца средних размеров (21–49 мкм). Больше всего пыльцы средних размеров у R. sanguineum, R. ussuriense, R. saxatile и R. aureum (91.8–94.6%).
Изученные растения имеют высокий процент морфологически сформированной (фертильной) пыльцы, окрашивающейся ацетокармином (80.3–96.4%). При этом обычно пыльца имеет довольно низкую жизнеспособность во время ее проращивания на искусственной питательной среде. Лучше всего прорастает пыльца R. sangui-neum, R. spicatum, R. nigrum, R. ussuriense, R. aureum (45–29%). При этом пыльцевые зерна могут прорастать с нарушениями.
У части видов Ribes пыльца способна прорастать, находясь еще внутри пыльника. Процент прорастания пыльцы внутри пыльника неодинаковый в цветках, взятых из разных участков соцветия; наилучшее прорастание имеется у пыльцы из цветков центральной зоны соцветия (15.3–82.9%).
Опылителями цветков Ribes в условиях Санкт-Петербурга являются представители Apis, Bombus и Chrysopa oculatа. Наблюдаются 2 пика активности опылителей: в утренние часы (10.00–11.00) и вечерние часы (17.00–18.00).
Прорастание пыльцы видов Ribes на рыльце пестика (in situ) происходит активно, но может сопровождаться нарушениями. При относительно высокой температуре воздуха прохождение пыльцевой трубки до семязачатка происходит за 3–5 суток. В семязачатках уже в первый день цветения наблюдаются зрелые зародышевые мешки, готовые к оплодотворению. В гнездах завязи развиваются по 4 геми-ортотропных семязачатка, расположенных в 6–10 “этажей”. Пыльца попадает на рыльце в двухклеточном состоянии. Деление генеративной клетки на два спермия происходит в пыльцевой трубке во время роста пыльцевой трубки.
Проведенные исследования позволяют рекомендовать R. sanguineum, R. spicatum, R. nigrum, R. aureum для использования в экспериментах по селекции Ribes в условиях Санкт-Петербурга.
Список литературы
[Afanas’eva] Афанасьева А.С. 1964. Роль дополнительного опыления в процессе оплодотворения. – Тр. института генетики АН СССР. 31: 34–42.
[Burmenko et al.] Бурменко Ю.В., Сорокопудов В.Н., Колесников Д.А., Маслова Н.Н. и др. 2009. Дополнение к морфологии пыльцы сем. Grossulariaceae Dumort. – Вестник КрасГАУ. 1: 25–31.
Erdtman G. 1952. Pollen morphology and plant taxonomy: Angiosperms. Stokholm. 539 p.
[Firsov, Yandovka] Фирсов Г.А., Яндовка Л.Ф. 2018. Смородины (Ribes L., Grossulariaceae) Ботанического сада Петра Великого. – Труды по прикладной ботанике, генетике и селекции. 179 (2): 31–43. https://doi.org/10.30901/2227-8834-2018-2-31-43
[Gavrilova, Tikhonova] Гаврилова О.А., Тихонова О.А. 2013. Разнообразие форм пыльцевых зерен и их распределение у некоторых видов и гибридов крыжовниковых. – Труды Кар НЦ. Сер. Экспериментальная биология. 3: 82–92.
[Gavrilova, Tikhonova] Гаврилова О.А., Тихонова О.А. 2016. Качество пыльцы видов и отдаленных гибридов в семействе Grossulariaceae DC. – Успехи современного естествознания. 12: 68–73.
URL:https://natural-sciences.ru/ru/article/view?id=36263 (дата обращения: 09.10.2022).
Hesse M., Halbritter H., Zetter R. et al. 2009. Pollen terminology. An illustrated handbook. New York. 261 p.
Huang P.H., Ye W.-h. 1989. Pollen morphology of Ribes L. and its taxonomic significance. – Acta Phytotaxonomica Sinica. 27 (5): 378–385.
[Kobasnidze] Кобаснидзе Э.Я. 1983. Эмбриология представителей сем. Grossulariaceae в условиях высокогорья Грузии: Дис. … канд. биол. наук. Тбилиси. 201 с.
[Kupriyanova, Aleshina] Куприянова Л.А., Алешина Л.А. 1972. Пыльца и споры растений флоры европейской части СССР. Л. 171с.
[Mandrik, Andrusyak] Мандрик В.Ю., Андрусяк М.А. 1969a. Изучение пыльцы, пыльцевых трубок и спермиогенеза у крыжовника Grossularia reclinata Mill. и смородины Ribes alpinum L. – Бот. журн. 54 (7): 1112–1117.
[Mandrik, Andrusyak] Мандрик В.Ю., Андрусяк М.А. 1969b. Макроспорогенез и развитие женского гаметофита у крыжовника (Grossularia reclinata Mill.) и альпийской смородины (Ribes alpinum L). – Бюл. МОИП. Отд. биол. 74 (5): 101–108.
[Melekhina, Yakobson] Мелехина А.А., Якобсон Л.И. 1971. Ход процессов опыления и оплодотворения при скрещивании Ribes nigrum L. × R. aureum Pursh. в связи с их несовместимостью. – Изв. Академии наук Латвийской ССР. 6 (287): 23–28.
[Popova] Попова А.Ф. 1965. Заложение и развитие генеративных органов у черной смородины Ribes nigrum L. – Укр. бот. журн. 22 (6): 83–89.
Punt W., Hoen P.P., Blackmore S. et al. 2007. Glossary of pollen and spore terminology. – Rev. Palaeobot. Palynol. 143 (1–2): 1–81.
[Radionenko] Радионенко А.Я. 1970. Процесс оплодотворения у Ribes nigrum L. – Бот. журн. 55 (5): 807–813.
[Romanova et al.] Романова Н.П., Шелаботин Г.П., Леонченко В.Г., Ханина Н.П. 1988. Методические рекомендации по применению цитологических методов в плодоводстве. М. 52 с.
[Shamrov] Шамров И.И. 2017. Морфологические типы семязачатков цветковых растений. – Бот. журн. 102 (2): 129–146. https://doi.org/10.1134/S0006813617020016
Verbeek-Reuvers A.A.M.L. 1977. The Northwest European Pollen Flora, 12. Grossulariaceae. – Rew. Palaeobot. Palynol. 24 (3): 107–116.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал