

Физиология человека, 2019, T. 45, № 1, стр. 5-18
Маскирующие эффекты нерелевантной зрительной информации в условиях базовой и суперординатной категоризации сложных изображений
Н. Ю. Герасименко 1, *, А. Б. Кушнир 1, Е. С. Михайлова 1
1 ФГБУН Институт высшей нервной деятельности и нейрофизиологии РАН
Москва, Россия
* E-mail: nataliagerasimenko@gmail.com
Поступила в редакцию 06.08.2018
После доработки 12.08.2018
Принята к публикации 01.10.2018
Аннотация
В психофизическом эксперименте участвовал 41 здоровый испытуемый. В исследовании анализировали точность и время реакции (ВР) при базовой и суперординатной категоризации изображений в условиях их прямой маскировки. В качестве целевых стимулов использовали изображения предметов и животных, в качестве масок – изображения предметов, животных, лиц и зданий. Сравнение психометрических характеристик базовой и суперординатной категоризации показало, что влияние семантического сходства маски и стимула на эффективность прямой маскировки зависело от типа выполняемой испытуемым экспериментальной задачи – уровня категоризации целевого стимула. При опознании на базовом уровне маскирующий эффект был более выражен при большем семантическом сходстве маски и целевого стимула. Напротив, при опознании на суперординатном уровне, больший маскирующий эффект наблюдали в ситуации, когда маска и стимул относились к разным категориям. Изображения лиц и зданий оказывали меньший маскирующий эффект по сравнению с изображениями животных и предметов независимо от типа задачи. Анализ распределения ВР показал, что при базовой категоризации рост ВР достигается за счет увеличения асимметрии распределения, а при суперординатной – за счет сдвига распределения в сторону больших значений. Результаты данного исследования свидетельствуют в пользу гипотезы о временном и пространственном разделении когнитивных процессов при базовой и суперординатной категоризации.
Зрительная категоризация – это способность человека отнести сенсорный стимул к какой-либо дискретной, поведенчески значимой категории [1]. Способность к категоризации существенно экономит наши когнитивные ресурсы. Если в новом объекте окружающего мира мы обнаруживаем информацию, позволяющую отнести его к определенной категории, мы можем без дополнительного обследования выдвигать относительно него релевантные категориальные суждения [2]. Категории имеют иерархическую структуру, образуя последовательность все более широких категорий, в каждую из которых предыдущая входит как составная часть. Один и тот же стимул на базовом уровне будет опознаваться как стул, стол или собака, на более высоком – суперординатном – уровне как животное или неодушевленный предмет, а на более узком – субординатном – как офисный стул или афганская борзая [3, 4].
В настоящее время нейробиологические основы категоризации изображений на разных уровнях, остаются до конца неясными [5]. Предполагается, что в процесс категоризации изображения в большей степени вовлечены нейроны вентрального пути передачи зрительной информации [6], который берет свое начало в первичной зрительной области (V1), продолжается через более высокие ретинотопически организованные зоны (V2, V3, V4) и достигает нижневисочной коры [5]. В нижневисочной коре обнаружены области, специфически отвечающие на стимулы наиболее экологически значимых категорий, таких как лица людей, части тел, локации и слова [5]. Но для многих других классов стимулов подобных областей не найдено. С учетом значительного разнообразия объектов окружающего нас мира, трудно представить, что каждому из них соответствует отдельная репрезентация. Поэтому сейчас выдвинуто предположение о существовании семантического континуума репрезентаций (continuous semantic space), внутри которого происходит плавный переход от одной категории к другой [7]. При этом в процессе категоризации важную роль играет префронтальная кора, модулирующие влияния которой “настраивают” нижневисочную кору, повышая селективность определенных групп нейронов. Показано, что при транскраниальной магнитной стимуляции префронтальной коры у испытуемых наблюдалось снижение селективности нейронов нижневисочной коры, выражающееся в перекрывании зон активации на лицевые стимулы и изображения зданий [8]. Наблюдения за пациентами с синдромом поражения лобной доли демонстрируют ведущую роль фронтальных отделов коры в категоризации на суперординатном уровне. Если пациентов спросить, чем сходны апельсин и банан, относящиеся к одной суперординатной категории “фрукты”, то они либо могут не увидеть их сходства вовсе (ответ испытуемого “один желтый, другой оранжевый”), либо относят их к одной категории руководствуясь ошибочными суждениями (“они оба сладкие”, “оба имеют кожуру”) [9].
На ход процесса категоризации в каждом отдельном случае влияет ряд факторов, среди которых фигурируют уровень внимания [10], тип задачи, выполняемой испытуемым [6], и контекст, предшествующий или совпадающий по времени со стимулом [11, 12]. В работе S.A. Nastase et al. в качестве стимулов использовали короткие фильмы, в которых испытуемый должен был обращать внимание либо на действия животных (летают, едят и т.д.), либо на их таксономическую принадлежность (приматы, птицы и т.д.). Сдвиг внимания испытуемого на таксоны животных приводил к увеличению дистанции между ними в континууме репрезентаций, выделенных по фМРТ-сигналу в нижневисочной коре, что может свидетельствовать об увеличении селективности ее ответов [10]. В работе L. Bugatus et al. сравнивали величину фМРТ-сигнала при выполнении испытуемым разных типов экспериментальных задач (одд-болл, задача на рабочую память и селективное внимание) при использовании стимулов, принадлежащих к различным категориям (лица, здания, автомобили, тела, псевдослова) [6]. Было показано, что тип задачи в большей степени влиял на активность вентральной части префронтальной коры, в то время как категориальная принадлежность стимула – на активность латеральной затылочно-височной и нижневисочной областей коры. Уровень внимания испытуемого оказывал значимое влияние на воспроизводимость (reproducibility) категориальной селективности ответов во всех тестируемых областях коры, что проявлялось в уменьшении воспроизводимости в ситуации невнимания (unattended conditions). В вентролатеральной префронтальной коре эта зависимость была максимально выраженной: категориальная селективность выявлялась только при условии привлечения внимания к данной категории изображений. В латеральной затылочно-височной и нижневисочной областях коры категориальная селективность наблюдалась и в том случае, когда данная категория стимулов находилась вне фокуса внимания испытуемого, увеличение уровня внимания к категории повышало селективность и в этих областях [6]. Также на процесс категоризации влияет сопутствующая стимулу зрительная информация. В случае, если изображение помещалось на неконгруэнтный фон (животное на фоне урбанистического пейзажа), его категоризация на суперординатном уровне ухудшалась по сравнению с предъявлением на конгруэнтном фоне [12].
В наших предыдущих работах для анализа влияния предшествующей зрительной информации мы использовали метод прямой зрительной маскировки, во время которой значимому стимулу предшествовало другое изображение, игнорируемое испытуемым по условию задачи. Нами было показано, что категоризация зрительных стимулов на базовом уровне затрудняется, если им предшествует семантически близкое незначимое изображение [13, 14]. Вместе с тем есть категории зрительных стимулов, которые при их использовании в качестве маскирующих изображений не вызывают ухудшения опознания целевого стимула. Одной из таких категорий являются лица. В модели прямой зрительной маскировки предшествующие целевому стимулу изображения лиц не снижали точность опознания и не увеличивали время двигательной реакции по сравнению с опознанием без маскировки [15].
Цель данной работы – в психофизическом эксперименте определить зависимость эффектов семантического сходства целевого и маскирующего изображения от типа выполняемой когнитивной задачи – категоризации изображений на базовом или суперординатном уровне. В качестве масок использовали изображения, относящиеся к тем же категориям, что и целевые стимулы (животные и предметы), а также изображения, относящиеся к обособленным категориям сложных зрительных стимулов – лица и здания.
МЕТОДИКА
Характеристика группы испытуемых. В исследовании принимали участие 41 испытуемый (22 женщины и 19 мужчин) с нормальным или скорректированным до нормы зрением в возрасте от 20 до 31 г. (24.4 ± 0.5 лет). Из них 5 чел по самоотчету были левшами, остальные – правшами. Исследование состояло из двух экспериментов. В первом испытуемых просили категоризовать изображения на базовом уровне, во втором – на суперординатном. Из всей группы испытуемых в эксперименте с опознанием на базовом уровне участвовали 29 чел (24.0 ± 0.5 лет), с опознанием на суперординатном уровне – 23 чел (25.2 ± 0.5 лет), в обоих экспериментах – 11 чел (24.8 ± 0.7 лет). Во всех случаях было получено письменное согласие испытуемых на проведение исследования, согласно протоколу, утвержденному этической комиссией Института высшей нервной деятельности и нейрофизиологии РАН. Участие в эксперименте оплачивали.
Стимулы. В качестве целевых стимулов использовали черно-белые фотографии животных и бытовых предметов, в качестве масок – черно-белые фотографии животных, бытовых предметов, зданий и лиц людей с эмоционально-нейтральным выражением. Фотографии лиц были взяты из базы лицевых стимулов Университета Неймегена [16]. Изображения животных, предметов и зданий выбрали нами из цветных фотографий, представленных в Интернете. Выбранные изображения вырезали из фона с помощью инструментов программы Adobe Photoshop CS4 и помещались по центру заранее созданного макета с белым фоном размером 800 × 600 пикселей. Размер самого изображения составлял 210 × 210 пикселей, разрешение – 72 точки на дюйм (72 dpi). Далее цветное изображение превращали в черно-белое переводя его из режима RGB в режим градаций серого. Для каждой из базовых категорий стимулов (стул, кувшин, собака и т.п.) выбрали по 4 разных изображений объекта. В табл. 1 приведен список предметов и животных, изображения которых использовали в качестве целевых стимулов и масок, а на рис. 1, А–Д примеры этих изображений.
Таблица 1.
Маскирующие и целевые стимулы, использованные в различных экспериментальных сериях
| Экспериментальная серия | Маскирующие стимулы | Целевые стимулы |
|---|---|---|
| Базовая категоризация животных | Животные: зебра (4), гепард (4), кролик (4), собака (4) | белка (4), лама (4), еж (4), носорог (4) |
| Предметы: расческа (4), автомобиль (4), ключ (4), кастрюля (4) | ||
| Лица с нейтральным эмоциональным выражением: женские (8) и мужские (8) | ||
| Здания (16) | ||
| Базовая категоризация предметов | Животные: бурундук (4), росомаха (4), волк (4), лошадь (4) | стол (4), шляпа (4), кувшин (4), замóк (4) |
| Предметы: настольная лампа (4), веник (4), чашка (4), вентилятор (4) | ||
| Лица с нейтральным эмоциональным выражением: женские (8) и мужские (8) лица | ||
| Здания (16) | ||
| Суперординатная категоризация | Животные: баран, медведь, бурундук, енот, собака, гепард, лошадь, коза, кролик, лев, олень, свинья, росомаха, тигр, волк, зебра, голубь, гусь, орел, петух, попугай, киви, утка, ворона, стрекоза, корова, улитка, краб, агама, окунь, карась, геккон | Животные: антилопа, барс, белка, верблюд, еж, хорек, лама, белый медведь, синица,
сова, воробей, бабочка, жук, черепаха, лягушка, рыба Предметы: бочка, ботинок, диван, домра, грузовик, колокольчик, корзина, кувшин, лейка, губная помада, туфля, стол, ведро, велосипед, зеркало, зонт |
| Предметы: барабан, бинокль, чайник, стул, чашка, ножницы, фен, флейта, фотоаппарат, гитара, расческа, кисть, утюг, кастрюля, ключ, клещи, коляска, настольная лампа, лодка, автомобиль, монитор, кухонная плита, пылесос, рояль, сковорода, совок, стиральная машина, трактор, ваза, веник, вентилятор, вертолет | ||
| Лица с нейтральным эмоциональным выражением: женские (16) и мужские (16) лица | ||
| Здания (32) |
Рис. 1.
Условия эксперимента. Примеры целевых и маскирующих изображений. А – целевые изображения, относящиеся к одной базовой категории “кувшин”, Б – стимулы четырех разных базовых категорий, использованные в одной экспериментальной серии, В – целевые стимулы суперординатной категории “животные”, Г – целевые стимулы суперординатной категории “предметы”, Д – маскирующие изображения четырех категорий: здания, лица, предметы и животные, Е – временная схема эксперимента.

Размер изображения на экране монитора – 8–10 см по ширине и по высоте, что в условиях эксперимента (120 см от экрана до глаз испытуемого) составляло 4–5 угл. град.
Описание эксперимента. В ходе эксперимента испытуемый сидел в удобном кресле в затемненной и звукозаглушенной комнате на расстоянии 120 см от экрана монитора Philips Brilliance 202P70/00 (диагональ 20", 60 Гц). Освещенность на уровне глаз испытуемого – 4 лк. Предъявление стимулов и регистрацию точности опознания и времени реакции (ВР) проводили с помощью программы E-Рrime 2.0 (Psychology Software Tools, Inc., США) с выносной клавиатурой Serial Response Box. Исследование проводили в дневное время (с 10 до 17 ч).
Временная схема эксперимента представлена на рис. 1, Е. Длительность предъявления маскирующего изображения – 102 мс, сразу после него на 85 мс предъявляли целевой стимул, который сменялся шумовой маской (50 мс). Такие временные параметры создавали максимальный маскирующий эффект и использовали в предыдущих исследованиях [13]. Межстимульный интервал – 3000 мс, во время него в центре экрана находилась фиксационная точка. При опознании стимула испытуемый нажимал на одну из клавиш выносной клавиатуры Serial Response Box. Испытуемых просили отвечать быстро, но стараясь не допускать большого количества ошибок. Для каждого испытуемого рассчитывали вероятность правильных ответов, или точность (в %), и среднее время реакции (ВР, мс) для правильных ответов.
Эксперимент с категоризацией на базовом уровне состоял из двух серий: опознание животных и опознание бытовых предметов при их прямой маскировке. В качестве маскирующих стимулов использовали изображения предметов, животных, а также лиц и зданий как объектов повседневной жизни. В каждой серии (опознание животных, опознание бытовых предметов) испытуемому предъявляли целевые стимулы, относящиеся к 4-м базовым категориям, для каждой из которых использовали по 4 разных фотографии, всего 16 изображений. В серии каждое изображение предъявляли по 16 раз, т.е. 256 предъявлений целевых стимулов. Из них в 64 случаях пары маска−целевой стимул были семантически близкими (животное−животное или предмет−предмет), в 64 случаях – семантически далекими (предмет−животное или животное−предмет), в 64 случаях маской было изображение лица, а еще в 64 случаях – изображение здания. В ходе эксперимента различные пары маска−стимул предъявляли в случайном порядке.
Эксперимент начинали с тренировочной серии для выработки моторного навыка, во время которой на экране в случайном порядке на 100 мс вспыхивали цифры от 1 до 4 (каждая по 12 предъявлений). При появлении цифры испытуемый должен был нажать на соответствующую ей кнопку клавиатуры.
Основную часть эксперимента начинали с появления на экране инструкции. Например, для задачи категоризации предметов на базовом уровне она выглядела так: “Во время опыта смотрите, пожалуйста, в центр экрана на маленький кружок. При появлении изображения стола нажмите на цифру 1. При появлении изображения шляпы нажмите на цифру 2. При появлении изображения кувшина нажмите на цифру 3. При появлении изображения замкá нажмите на цифру 4". Для серии с опознанием животных инструкция была аналогичной, с поправкой на используемые целевые стимулы. Каждую экспериментальную серию предваряли тренировкой, в ходе которой целевые стимулы предъявляли без маскировки. В тренировочной части длительность стимула составляла 85 мс, межстимульного интервала – 3000 мс, каждое целевое изображение предъявляли 1 раз, всего 16 предъявлений. После окончания тренировки на экране появлялась надпись "Основная часть” и начинали предъявление замаскированных целевых изображений (параметры предъявления изображений в основной части описаны выше).
Эксперимент с категоризацией на суперординатном уровне. Задачей испытуемого было отнести целевое изображение к категориям “животные” или “предметы”. Испытуемые категоризовали стимулы при их прямой маскировке четырьмя видами маскирующих изображений: предметы, животные, лица и здания. В качестве целевых стимулов использовали 16 изображений животных и 16 изображений предметов. Каждое изображение предъявляли по 8 раз, всего 256 предъявлений (по 32 пары животное−животное, предмет−предмет, предмет−животное, животное−предмет, лицо-животное, лицо−предмет, здание−животное, здание−предмет). Последовательность предъявления различных пар маска−стимул была случайной.
Эксперимент начинали с серии выработки моторного навыка. Основную часть эксперимента начинали с появления следующей инструкции: “Во время опыта смотрите, пожалуйста, в центр экрана на маленький кружок. При появлении изображения животного нажмите на цифру 1. При появлении изображения предмета нажмите на цифру 2”. При этом половина испытуемых по инструкции должны были нажимать на первую клавишу для категории “животное”, на вторую – для категории “предмет”. Для второй половины испытуемых номера клавиш меняли: первая клавиша – “предмет”, вторая – “животное”. Также как в серии с базовой категоризацией, вначале следовала тренировка, в ходе которой целевые изображения предъявляли без маски. После тренировки начиналась основная часть.
Длительность одной экспериментальной серии около 16 мин с одним небольшим интервалом посередине, продолжительность которого регулировал сам испытуемый.
Анализ распределения времени реакции. Согласно литературным данным, распределение времени реакции (ВР) отличается от нормального и подчиняется экс-Гауссову распределению (ex-Gaussian distribution) [17, 18]. Типичное распределение ВР ассиметрично и скошено вправо, т.е. имеет “хвост” больших значений ВР. Экс-Гауссова функция рассматривает распределение ВР как сочетание двух распределений: Гауссова (нормального), описывающего центральную часть распределения, и экспоненциального, характеризующего “хвост” больших значений. Для анализа экс-Гауссова распределения используют три параметра: µ и σ, описывающих среднее значение и стандартное отклонение нормальной части распределения соответственно, и τ, характеризующего среднее значение и стандартное отклонение экспоненциальной части [17]. Для вычисления параметров распределение ВР использовали метод непрерывной оценки максимального правдоподобия (continuous maximum likelihood estimation). Вычисление параметров µ, σ и τ проводили в программной среде RStudio 1.1.453 (RStudio, Inc.) с помощью скрипта, написанного A. Heathcote [19].
При статистической обработке данных использовали дисперсионный анализ – метод повторных измерений, ANOVA RM. Учитывали влияние факторов УРОВЕНЬ КАТЕГОРИЗАЦИИ, СТИМУЛ и МАСКА. Использовали несколько схем дисперсионного анализа. Анализ влияния уровня категоризации проводили с учетом факторов УРОВЕНЬ КАТЕГОРИЗАЦИИ (2 уровня: базовый, суперординатный), СТИМУЛ (2 уровня: животное, предмет) и МАСКА (2 уровня: семантически близкая к целевому стимулу и семантически далекая). Также проводили анализ точности и ВР отдельно для базового и суперординатного уровней категоризации с учетом влияния факторов СТИМУЛ (2 уровня: животные и предметы) и МАСКА (4 уровня: маска семантически близкая целевому стимулу и маска далекая от целевого стимула, маска-дом, маска-лицо). Проводили post-hoc-анализ с использованием теста Ньюмана-Кейлса. Статистический анализ проводили в программах SPSS 20.0 и Statistica 10.0.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Влияние уровня категоризации на психофизические характеристики опознания изображения. Для выявления зависимости времени реакции и вероятности правильного ответа от уровня категоризации (базовый или суперординатный) проведен дисперсионный анализ (ANOVA RM) с учетом факторов УРОВЕНЬ КАТЕГОРИЗАЦИИ, СТИМУЛ и МАСКА.
Для ВР выявлен значимый эффект УРОВНЯ КАТЕГОРИЗАЦИИ F(1, 10) = 15.98; p < 0.01 и взаимодействие УРОВЕНЬ КАТЕГОРИЗАЦИИ × МАСКА F(1, 10) = 30.06, p < 0.0005 и СТИМУЛ × МАСКА F(1, 10) = 6.01, p < 0.05. На рис. 2, А приведены средние значения ВР при выполнении испытуемыми двух вариантов категоризации изображений. Видно, что эффекты маскировки в этих двух ситуациях имеют противоположный характер. При базовой категоризации ВР больше при маскировке стимула семантически близкими изображениями, при суперординатной – семантическими далекими. Значимость эффектов была неодинаковой для животных и предметов. При базовой категоризации для животных уровень значимости эффекта МАСКИ соответствовал p < 0.01, для предметов отличия недостоверны, при суперординатной категоризации уровень значимости эффекта МАСКИ и для животных, и для предметов был одинаковым –p < 0.01 (здесь и далее критерий Ньюмана−Кейлса).
Рис. 2.
Влияние уровня категоризации на точность опознания, ВР и его распределение при различных типах маскировки. А – время реакции (мс), Б – вероятность правильного ответа (%). Слева приведены параметры опознания при базовой, справа – при суперординатной категоризации. Приведены средние по группе значения и ошибка среднего. На оси абсцисс цифрами обозначены пары маска−стимул: 1 – маска-предмет–стимул-животное (семантически далекая пара), 2 – маска-животное стимул-животное (семантически близкая пара), 3 – маска-животное–стимул-предмет (семантически далекая пара), 4 – маска-предмет–стимул-предмет (семантически близкая пара). Достоверность различий: * – p < 0.05, ** – p < 0.01 (критерий Ньюмана−Кейлса). В – график распределения индивидуальных ВР испытуемых при базовой категоризации, Г – график распределения индивидуальных ВР испытуемых при суперординатной категоризации, а – данные для семантически близких маски и стимуле, б – для семантически далеких. Приведены данные для опознания животных.
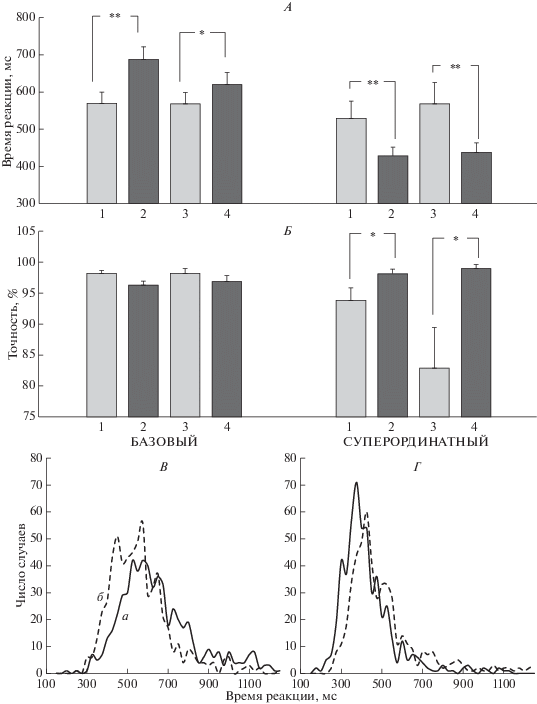
Для вероятности правильного ответа выявлены значимые взаимодействия УРОВЕНЬ КАТЕГОРИЗАЦИИ × МАСКА F(1, 10) = 11.7, p < 0.01 и СТИМУЛ × МАСКА F(1, 10) = 5.0, p < 0.05. На рис. 2, Б видно, что при базовой категоризации вероятность правильного ответа меньше при использовании семантически близкой маски. Для суперординатной категоризации зависимость обратная: точность выше при маскировке семантически близкими изображениями, чем семантически далекими (p < 0.05).
На рис. 2, В и Г приведены распределения индивидуальных ВР правильных ответов для базовой и суперординатной категоризации при разной семантической близости маски и целевого стимула. Анализ этих данных показал, что во всех случаях распределение не подчиняется закону нормального распределения (p < 0.0005, критерий Шапиро−Уилка). Для дополнительного анализа сочетанного эффекта варианта маскировки и характера выполняемой задачи (категоризация базовая или суперординатная) проведено сравнение параметров экс-Гауссова распределения ВР µ и σ, отражающих среднее значение и стандартное отклонение Гауссовой функции соответственно, и τ, отражающей среднее значение и стандартное отклонение экспоненциальной функции. Полученные результаты приведены в табл. 2.
Таблица 2.
Параметры экс-Гауссова распределения индивидуальных значений времени реакции
| Уровень категоризации | Маскирующий стимул | Целевой стимул | µ | σ | τ |
|---|---|---|---|---|---|
| Базовый | Животное | Животное | 543.4 (38.5) | 204.6 (22.8) | 159.0 (17.8) |
| Предмет | 469.8 (30.4) | 146.1 (28.1) | 114.8 (25.6) | ||
| Предмет | Предмет | 486.5 (26.7) | 220.0 (27.0) | 167.2 (20.1) | |
| Животное | 443.8 (35.9) | 153.2 (10.7) | 149.9 (17.6) | ||
| Суперординатный | Животное | Животное | 349.9 (11.3) | 117.5 (16.3) | 95.7 (16.7) |
| Предмет | 437.1 (40.5) | 167.3 (32.2) | 124.2 (20.6) | ||
| Предмет | Предмет | 377.0 (31.1) | 136.8 (35.5) | 114.9 (26.7) | |
| Животное | 463.5 (28.9) | 169.8 (32.9) | 114.6 (23.0) |
Проведен дисперсионный анализ ANOVA RM индивидуальных параметров распределения ВР. Для µ выявлен эффект УРОВНЯ КАТЕГОРИЗАЦИИ F(1, 8) = 5.71, p < 0.05 и взаимодействие СТИМУЛ × УРОВЕНЬ КАТЕГОРИЗАЦИИ F(1, 8) = 7.44; p < 0.05 и УРОВЕНЬ КАТЕГОРИЗАЦИИ × МАСКА F(1, 8) = 10.67; p < 0.05. При базовой категоризации µ был больше для семантически близких пар маска-стимул, при суперординатной, наоборот, для семантически далеких. Зависимость µ от семантической близости маски и целевого стимула более выражена для суперординатной категоризации (p < 0.05 и для животных, p = 0.08 для предметов) по сравнению с базовой.
Для характеристики σ обнаружено взаимодействие УРОВЕНЬ КАТЕГОРИЗАЦИИ × МАСКА F(1, 8) = 14.59, p < 0.01. Изменения параметра σ при разных уровнях категоризации имеют разнонаправленный характер. При базовой категоризации σ выше в случае семантически близкой маски по сравнению с далекой, при суперординатной, наоборот, выше при семантически далекой маске.
Для характеристики τ обнаружено взаимодействие УРОВЕНЬ КАТЕГОРИЗАЦИИ × МАСКА F(1, 8) = 13.94, p < 0.01. Изменения в параметре τ отчетливее в случае базовой категоризации, при которой τ больше для семантически близких пар маска-стимул по сравнению с семантически далекими.
Таким образом, при выполнении испытуемым задачи на базовую категоризацию эффективность маскировки выше при совпадении категориальной принадлежности стимула и маски. Напротив, при суперординатной категоризации стимулов больший маскирующий эффект оказывают изображения другой категории – животные при опознании предметов и предметы – при опознании животных. Анализ распределения ВР показал, что при суперординатной категоризации рост ВР сопровождается увеличением параметров, характеризующих Гауссову часть распределения (µ и σ). В случае базовой категоризации рост ВР сопровождается не столько увеличением µ, отражающим среднее значение, сколько увеличением σ, характеризующим дисперсию Гауссовой части, и τ, описывающем вклад в распределение “хвоста” больших значений.
В связи с выявлением достоверного эффекта фактора УРОВЕНЬ КАТЕГОРИЗАЦИИ и взаимодействия УРОВЕНЬ КАТЕГОРИЗАЦИИ × МАСКА в дальнейшем влияние фактора МАСКА было рассмотрено отдельно в случае базовой и суперординатной категоризации.
Базовая категоризация. Проведен дисперсионный анализ ВР и точности с учетом факторов СТИМУЛ (2 уровня) и МАСКА (4 уровня). Для ВР выявлен эффект фактора МАСКА F(3, 84) = 70.07, p < 0.0005 и взаимодействие СТИМУЛ × МАСКА F(3, 84) = 24.99, p < 0.0005. Как видно на рис. 3, А, при базовой категоризации стимулов большее ВР наблюдалось при маскировке изображениями семантически близкими целевому стимулу (p < 0.001 по сравнению с семантически далекой маской).
Рис. 3.
Влияние маски на ВР и вероятность правильного ответа при базовой категоризации изображений животных (слева) и предметов (справа). А – время реакции (мс), Б – вероятность правильного ответа (%). Приведены средние по группе значения и ошибка среднего. На оси абсцисс цифрами обозначены варианты маскировки: 1 – маска-здание, 2 – маска-лицо, 3 – семантически далекая маска (предметы для животных и животные для предметов), 4 – семантически близкая маска (животные для животных и предметы для предметов). Достоверность различий: *** – p < 0.001 (критерий Ньюмана−Кейлса). Остальные обозначения см. рис. 2.
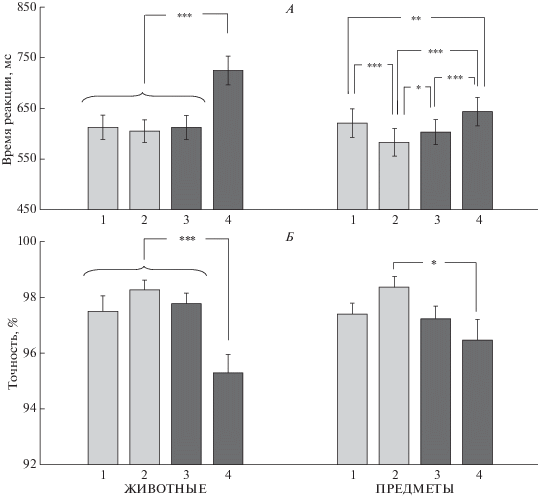
Для точности выявлен эффект фактора МАСКА F(3, 84) = 10.71, p < 0.0005. На рис. 3, Б видно, что большее маскирующее воздействие оказывают семантически близкие изображения, но при сравнении семантически близкой и семантически далекой масок значимое снижение точности наблюдается только для пары маска–животное, целевой стимул–животное (p < 0.001 по сравнению с маскировкой предметами).
Как видно на рис. 3, изображения животных более сильные маскеры как для изображений других животных, так и для предметов, но сами более устойчивы к маскировке изображениями других категорий, когда выступают в роли стимулов. Независимо от категории целевого стимула, наименьший маскирующий эффект оказывают изображения лиц, что проявляется в меньшем ВР (0.001 < p < 0.05 по сравнению с остальными масками для предметов и p < 0.001 по сравнению с семантически близкой маской для животных) и большей точности (p < 0.05 и p < 0.001 по сравнению с семантически близкой маской для предметов и животных, соответственно) при этом типе маски. Изображения зданий не оказывают отчетливого маскирующего влияния на изображения животных, но маскируют предметы (для ВР p < 0.05 по сравнению с маскировкой животными). Однако их маскирующий эффект меньше, чем для семантически близких масок (p < 0.01).
Суперординатная категоризация. Проведен дисперсионный анализ (ANOVA RM) с учетом факторов СТИМУЛ (2 уровня) и МАСКА (4 уровня). Для ВР выявлены значимый эффект МАСКИ F(3, 45) = 12.23, p < 0.0005 и взаимодействие СТИМУЛ × МАСКА F(3, 45) = 4.44, p < 0.05. Как видно на рис. 4, А, большее ВР наблюдается при маскировке изображениями, принадлежащими к другой семантической категории, а именно: животными для предметов (p < 0.001 по сравнению с масками той же категории) и предметами для животных (p < 0.01).
Рис. 4.
Влияние маски на ВР и вероятность правильного ответа при суперординатной категоризации изображений животных (слева) и предметов (справа). Обозначения см. рис. 3.
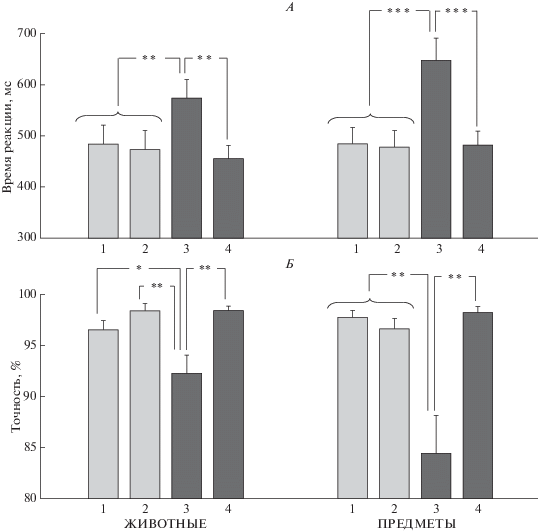
Для точности показан значимый эффект МАСКИ F(3, 45) = 9.77, p < 0.01. Вероятность правильного ответа снижается при маскировке семантически далекими изображениями (p < 0.01) (рис. 4, Б).
Как видно на рис. 4, изображения животных являются более сильной маской и более устойчивы к маскирующему воздействию, если используются как целевые стимулы. Наибольший маскирующий эффект наблюдается при маскировке предметов изображениями животных, он более выражен, чем для пары предмет−животное. Маскирующие эффекты маски-лица и маски-здания не отличаются от таковых для семантически близких изображений.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Сравнение психометрических характеристик базовой и суперординатной категоризации изображений животных и бытовых предметов показало, что влияние семантического сходства маски и стимула на эффективность прямой маскировки зависело от типа выполняемой когнитивной задачи. При базовом уровне категоризации, когда испытуемый должен был дать конкретное название объекту, маскирующий эффект был сильнее при семантическом сходстве маски и целевого стимула. Напротив, при опознании на суперординатном уровне, когда испытуемый должен был отнести объект к категориям животные или предметы, маскирующий эффект был выше в ситуации, когда маска и стимул были семантически различными, а нерелевантный стимул, принадлежавший той же категории, что и значимый, облегчал опознание целевого. Нужно подчеркнуть, что в данной работе не проводится сравнение абсолютных значений ВР при разных типах категоризации, так как количество альтернатив было различным: при базовой категоризации – четыре, а при суперординатной – две. При таких условиях эксперимента основным результатом является разнонаправленность эффектов семантической близости стимула и маски на ВР и точности при базовой и суперординатной категоризации.
Представляет интерес сравнение полученных нами результатов с данными других исследований, когда в задаче категоризации изображений значимая информация сочеталась с предшествующей или одновременно предъявляемой нерелевантной. Например, в работе O.R. Joubert et al. [12] исследовали влияние контекста на суперординатную категоризацию изображений животных и предметов. Изображения, помещенные на конгруэнтный фон (природный ландшафт для животных и урбанистический пейзаж для предметов) опознавались быстрее, чем стимулы, предъявляемые на неконгруэнтном фоне. Сходные результаты были получены при использовании экспериментальной модели совмещения изображения и слова (picture-word interference). Задачей испытуемых было категоризовать изображение в присутствии дистрактора – слова, предъявляемого визуально или аудиально. При категоризации изображения на базовом уровне семантически близкий дистрактор (например, слово “собака” для изображения кошки) вызывал увеличение ВР опознания. Авторы предполагают, что в этом случае дистрактор затрудняет выполнение задания по механизму лексического соревнования (lexical competition): он активирует свою репрезентацию и тормозит находящиеся рядом репрезентации семантически близких значимых стимулов [18, 20]. При суперординатной категоризации семантически близкий дистрактор (например, слово “лошадь” для изображения собаки), напротив, уменьшал ВР. Этот эффект авторы связывают с механизмом конгруэнции сообщения (message congruency): слово-дистратор конгруэнтно опознаваемому стимулу (например, слово “кошка” конгруэнтно изображению собаки) и соответствует той же суперординатной категории [18, 20].
В литературе при обсуждении нейробиологических основ категоризации изображений основная роль отводится структурам префронтальной и нижневисочной областей коры [5, 6, 21–25]. Функциональная значимость этих структур для операции категоризации неодинакова. В этом плане особенно убедительны результаты экспериментальных исследований D.J. Fridmann et al. [21–23] и J.A. Cromer et al. [26]. Регистрация нейронных ответов коры мозга приматов при выполнении ими задачи отставленного сопоставления категорий (delayed match-to-category task – DMС) показала, что нейроны префронтальной коры селективно реагируют на заранее выученные категории объектов. Многие нейроны демонстрируют бинарную категориальную настройку (binary-like categorical tuning), например, для объектов различных, но объединенных по принципу категориального сходства (кошка и собака различны, но объединены в категорию животных). Напротив, нейроны нижневисочной коры преимущественно реагируют селективно на стимулы, сходные в большей степени перцептуально, а не по принципу принадлежности к одной категории [23, 25, 27]. Используя прием морфинга изображений (машин различных марок, животных из различных семейств), D.J. Fridmann et al. [21, 22] и J.A. Cromer et al. [26] показали, что в DMC задаче нейроны префронтальной коры обезьян значительно раньше (первые 500 мс после начала эталонного стимула) реагируют на категориальное сходство, чем на различия физических характеристик (более 1000 мс после стимула). Т.е., категоризация осуществляется на уровне областей мозга, связанных с высшими поведенческими или когнитивными функциями, а именно префронтальной коры в большей степени, чем в областях, где кодируются отдельные характеристики, а именно в нижневисочной коре.
Исходя из изложенного выше, можно предположить, что разница наблюдаемых эффектов семантической близости маски и стимула при разных типах категоризации связаны c корковым уровнем, на котором происходит взаимодействие двух информационных посылок. При базовой категоризации на уровне нижневисочной коры возникает интерференция представительств семантически близких маскирующего и целевого стимулов. В работе M. Weber et al. показана прямая зависимость взаимного расположения зон активации на изображения животных от их семантической близости [28]. Позднее по данным фМРТ, зарегистрированным при просмотре испытуемыми видеофильмов, содержащих объекты и действия различных категорий (здания, люди, животные, предметы, бежать, говорить и т.д.), был построен непрерывный семантический континуум (continuous semantic space), внутри которого происходит плавный переход от одной категории к другой [7]. При этом воксели, активирующиеся на изображения родственных категорий, были расположены ближе друг к другу по сравнению с вокселями, активирующимися на неблизкие категории.
В настоящее время вопрос о том, на каком уровне (базовом или суперординатном) происходит первоначальная категоризация изображения остается открытым. Первоначально предполагалось, что все изображения сначала относятся к какой-либо базовой категории, а потом уже категоризуются на суперординатном уровне [29]. При этом базовый уровень был обозначен термином “уровень ввода” – entry level [30]. Вместе с тем, есть и другие данные. Анализ ВР опознания изображений показал, что при короткой длительности предъявления стимулы быстрее категоризуются на суперординатном уровне, при большей – на базовом [31, 32]. Однако существует и третья гипотеза, согласно которой изображения всегда первоначально категоризуются на суперординатном уровне [4]. В пользу этой гипотезы говорят данные о том, что в процессе развития ребенка суперординатные категории – “живое” и “неживое” - появляются раньше базовых [2]. В работе H. Banno и J. Saiki продемонстрирована неоднозначность такого “опережения” суперординатного уровня [33]. В работе проводили сравнение ВР в парадигме “Go/No-Go” при выполнении задач суперординатной категоризации сцен (природные пейзажи vs. урбанистические ландшафты и здания vs. сцены внутри помещений) и базовой категоризации отдельных объектов, принадлежавших природным пейзажам, сценам, ландшафтам (побережье vs. горы или автобан) [33]. Более быстрое опознание на суперординатном уровне наблюдалось только в случае категоризации природные пейзажи vs. урбанистические ландшафты, а категоризация здания vs. сцены внутри помещений происходила даже несколько медленнее, чем базовая категоризация. По мнению авторов, одно из возможных объяснений полученных результатов состоит в том, что для разделения природных и созданных человеком объектов (природные пейзажи vs. урбанистические ландшафты) достаточно низкоуровневых зрительных характеристик, так как в изображениях искусственных объектов статистически преобладают вертикальные и горизонтальные линии.
В нашей работе был проведен более детальный анализ показателя индивидуального ВР по характеристикам экс-Гауссова распределения [17, 18]. Согласно этой модели, параметры, характеризующие Гауссов компонент распределения (µ и σ), отражают время затраченное на кодирование стимула (stimulus encoding), или перцептивное звено, и организацию моторного ответа (motor response), или моторное звено, тогда как экспоненциальный компонент, характеризующийся параметром τ, преимущественно связан с когнитивной нагрузкой и центральным механизмом принятия решения [33]. Мы выявили, что при базовой категоризации увеличение маскирующего эффекта при семантическом сходстве маски и целевого стимула сопровождается увеличением не только величины µ и σ, но и τ, что говорит об увеличении нагрузки на центральное звено переработки информации. Эти данные хорошо сопоставимы с ранее полученными нами данными о возрастании интерквартильного интервала ВР при семантическом сходстве маскирующего и целевого стимулов [14]. Следовательно, в задаче базовой категоризации повышение τ в ситуации категориальной близости маски и целевого стимула может рассматриваться как показатель возросшей нагрузки на когнитивное звено переработки информации. Это предположение согласуется с мнением других авторов о большем объеме информации, необходимой для операции категоризации на базовом уровне по сравнению с суперординатным [34, 35].
В работе показано, что лица и здания при их использовании в качестве маскирующих стимулов, оказывали минимальный маскирующий эффект независимо от уровня категоризации целевого стимула. Вероятней всего, это связано с топографической обособленностью переработки этих стимулов в коре мозга. Есть сведения о существовании в мозге областей, селективно отвечающих на предъявление таких изображений: для лиц это fusiform face area (FFA), для зданий – parahippocampal place area (PPA) [5, 24, 36]. Многочисленные сведения указывают на уникальность лицевого стимула для зрительного восприятия человека. Так, например, опознание лица, в отличие от других изображений, устойчиво к обратной маскировке, эффекту мигания внимания (attention blink) и краудингу (crowding) [37, 38]. Согласно некоторым экспериментальным данным, лица являются особой категорией стимулов, восприятие которых не зависит от трудности основной задачи [39, 40]. Можно предположить, что восприятие лиц происходит автоматически и не зависит от их значимости для выполнения основной задачи, что позволяет предположить наличие механизма селективного внимания именно к лицам [39, 40]. В отличие от других категорий изображений, здания содержат в себе пространственную информацию и воспринимаются как “метки на местности” (landmarks), играя важную роль в навигации [41]. Таким образом, процесс обработки зрительной информации о маске-лицо и маске-здании не интерферирует с обработкой значимого стимула, и эти изображения не оказывают отчетливого маскирующего воздействия при опознании животных и предметов.
ЗАКЛЮЧЕНИЕ
В работе получены новые данные о влиянии типа выполняемой когнитивной задачи (базовый или суперординатный уровни категоризации изображений) на эффекты семантического сходства маски и целевого стимула. При базовой категоризации маскирующий эффект больше при семантической близости маскирующего и целевого изображений. При суперординатной категоризации семантическое сходство, напротив, улучшает опознание. Выявлена зависимость параметров экс-Гауссова распределения ВР от уровня категоризации. При базовой категоризации в отличие от суперординатной, распределение асимметрично, выше значения τ, характеризующего экспоненциальный компонент, что свидетельствует о большей нагрузке на когнитивное звено переработки информации в присутствии нерелевантного дистрактора и соответствует данным других авторов о большем объеме информации, необходимой для операции категоризации на базовом уровне. Обсуждаются различия в организации нервных сетей при опознании сложных изображений в присутствии нерелевантного дистрактора при базовом и суперординатном уровнях категоризации изображений.
Изображения лиц и зданий, принадлежащие к обособленным категориям стимулов, не оказывали выраженного маскирующего эффекта независимо от уровня категоризации. Вероятно, это связано с обособленностью обработки зрительной информации этих стимулов в нижневисочной коре, за счет чего явления интерференции маскирующих и целевых стимулов существенно меньше.
Работа выполнена по государственному заданию ИВНД и НФ РАН.
Список литературы
Freedman D.J., Assad J.A. Neuronal mechanisms of visual categorization: an abstract view on decision making // Annu. Rev. Neurosci. 2016. V. 8. № 39. P. 129.
Котов А.А. Индуктивный вывод в разных возрастах // Избранные разделы психологии научения: коллективная монография / Под. ред. Спиридонова В.Ф. М.: Издательский дом “Дело” РАНХиГС, 2017. С. 9.
Tanaka J., Luu P., Weisbrod M., Kiefer M. Tracking the time course of object categorization using event-related potentials // NeuroReport. 1999. V. 10. № 4. P. 829.
Large M.E., Kiss I., McMullen P.A. Electrophysiological correlates of object categorization: back to basics // Cogn. Brain Res. 2004. V. 20. P. 415.
Grill-Spector K., Weiner K.S. The functional architecture of the ventral temporal cortex and its role in categorization // Nat. Rev. Neurosci. 2014. V. 15. № 8. P. 536.
Bugatus L., Weiner K.S., Grill-Spector K. Task alters category representations in prefrontal but not high-level visual cortex // NeuroImage. 2017. V. 155. P. 437.
Huth A.G., Nishimoto S., Vu A.T., Gallant J.L. A continuous semantic space describes the representation of thousands of object and action categories across the human brain // Neuron. 2012. V. 76. P. 1210.
Miller B.T., Vytlacil J., Fegen D. et al. The prefrontal cortex modulates category selectivity in human extrastriate cortex // J. Cogn. Neurosci. 2010. V. 23. № 1. P. 1.
Garcin B., Volle E., Dubois B., Levy R. Similar or different? The role of the ventrolateral prefrontal cortex in similarity detection // PLoS ONE. 2012. V. 7. № 3. e34164.
Nastase S.A., Connolly A.C., Oosterhof N.N. et al. Attention selectively reshapes the geometry of distributed semantic representation // Cereb. Cortex. 2017. V. 27. № 8. P. 4277.
Damian M.F. Semantic negative priming in picture categorization and naming // Cognition. 2000. V. 76. № 2. P. B45.
Joubert O.R., Fize D., Rousselet A., Fabre-Thore M. Early interference of context congruence on object processing in rapid visual categorization of natural scenes // J. Vis. 2008. V. 8. № 13. P. 1.
Герасименко Н.Ю., Славуцкая А.В., Калинин С.А. и др. Опознание зрительных образов в условиях их прямой маскировки. Влияние категориальной близости значимого и маскирующих изображений // Журн. высш. нерв. деятельности. 2013. Т. 63. № 4. С. 419.
Калинин С.А., Герасименко Н.Ю., Славуцкая А.В., Михайлова Е.С. Поведенческие и электрографические характеристики опознания сложных изображений в условиях их прямой маскировки. Влияние категориальной близости целевого и маскирующего стимулов // Физиология человека. 2014. Т. 40. № 4. С. 5.
Герасименко Н.Ю., Славуцкая А.В., Калинин С.А., Михайлова Е.С. Влияние неосознаваемого восприятия эмоциональных лиц на опознание сложных зрительных образов // Физиология человека. 2013. Т. 39. № 3. С. 26.
Langner O., Dotsch R., Bijlstra G. et al. Presentation and validation of the Radboud Faces Database // Cogn. Emot. 2010. V. 24. № 8. P. 1377.
Balota D.A., Yap M.J., Cortese M.J., Watson J.M. Beyond mean response latency: Response time distributional analyses of semantic priming // J. Mem. Lang. 2008. V. 59. P. 495.
Roelofs A., Piai V. Distributional analysis of semantic interference in picture naming // Q. J. Exp. Psychol. 2017. V. 70. № 4. P. 782.
Heathcote A. Fitting Wald and ex-Wald distributions to response time data: An example using functions for the S-PLUS package // Behav. Res. Methods. Instrum. Comput. 2004. V. 36. № 4. P. 678.
Hantsch A., Jescheniak J.D., Madebach A. Naming and categorizing objects: Task differences modulate the polarity of semantic effects in the picture–word interference paradigm // Mem. Cognit. 2012. V. 40. № 5. P. 760.
Freedman D.J., Riesenhuber M., Poggio T., Miller E.K. Categorical representation of visual stimuli in the primate prefrontal cortex // Science. 2001. V. 291. P. 312.
Freedman D.J., Riesenhuber M., Poggio T., Miller E.K. Visual categorization and the primate prefrontal cortex: neurophysiology and behavior // J. Neurophysiol. 2002. V. 88. № 2. P. 929.
Freedman D.J., Riesenhuber M., Poggio T., Miller E.K. A comparison of primate prefrontal and inferior temporal cortices during visual categorization // J. Neurosci. 2003. V. 23. № 12. P. 5235.
Bunzeck N., Schutze H., Duzel E. Category-specific organization of prefrontal response-facilitation during priming // Neuropsychol. 2006. V. 44. P. 1765.
McKee J.L., Riesenhuber M., Miller E.K., Freedman D.J. Task dependence of visual and category representations in prefrontal and inferior temporal cortices // J. Neurosci. 2014. V. 34. № 48. P. 16065.
Cromer J.A., Roy J.E., Buschman T.J., Miller E.K. Comparison of primate prefrontal and premotor cortex neuronal activity during visual categorization // J. Cogn. Neurosci. 2011. V. 23. № 11. P. 3355.
Freedman D.J., Riesenhuber M., Poggio T., Miller E.K. Experience-dependent sharpening of visual shape selectivity in inferior temporal cortex // Cereb. Cortex. 2006. V. 16. № 11. P. 1631.
Weber M., Thompson-Schill S.L., Osherson D. et al. Predicting judged similarity of natural categories from their neural representations // Neuropsychol. 2009. V. 47. P. 859.
Rosch E., Mervis C.B., Gray W.D. et al. Basic objects in natural categories // Cogn. Psychol. 1976. V. 8. P. 382.
Jolicoeur P., Gluck M.A., Kosslyn S.M. Pictures and names: making the connection // Cogn. Psychol. 1984. V. 16. P. 243.
Mack M.L., Palmeri T.J. The dynamics of categorization: unraveling rapid categorization // J. Exp. Psychol. 2015. V. 144. № 3. P. 551.
Vanmarcke S., Calders F., Wagemans J. The time-course of ultrarapid categorization: the influence of scene congruency and top-down processing // I-Persept. 2016. V. 7. № 5. P. 2041669516673384.
Banno H., Saiki J. The processing speed of scene categorization at multiple levels of description: The superordinate advantage revisited // Perception. 2015. V. 44. № 3. P. 269.
Joubert O.R., Rousselet G.A., Fabre-Thorpe M., Fize D. Rapid visual categorization of natural scene contexts with equalized amplitude spectrum and increasing phase noise // J. Vision. 2009. V. 9. № 1. P. 1.
Greene M.R., Oliva A. Recognition of natural scenes from global properties: seeing the forest without representing the trees // Cogn. Psychol. V. 58. № 2. P. 137.
Downing P.E., Chan A.W.Y., Peelen M.V. et al. Domain specificity in visual cortex // Cereb. Cortex. 2005. V. 16. № 10. P. 1453.
Shelley-Tremblay J., Mack A. Metacontrast masking and attention // Psychol. Sci. 1999. V. 10. № 6. P. 508.
Mack A., Pappas Z., Silverman M., Gay R. What we see: inattention and the capture of attention by meaning // Conscious. Cogn. 2002. V. 11. № 4. P. 488.
Lavie N., Ro T., Russell C. The role of perceptual load in processing distractor faces // Psychol. Sci. 2003. V. 14. № 5. P. 510.
Neumann M.F., Mohamed T.N., Schweinberger S.R. Face and object encoding under perceptual load: ERP evidence // NeuroImage. 2011. V. 54. P. 3021.
Sala J.B., Rama P., Courtney S.M. Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory // Neuropsychol. 2003. V. 41. P. 341.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека