

Физиология человека, 2020, T. 46, № 1, стр. 22-35
Зависимость эффективности прямой маскировки социально значимыми изображениями от категории маски и ее пространственно-частотных характеристик
Н. Ю. Герасименко 1, *, А. Б. Кушнир 1, Е. С. Михайлова 1
1 ФГБУН Институт высшей нервной деятельности и нейрофизиологии РАН
Москва, Россия
* E-mail: nataliagerasimenko@gmail.com
Поступила в редакцию 22.05.2019
После доработки 28.06.2019
Принята к публикации 03.08.2019
Аннотация
В психофизиологическом эксперименте участвовали 20 здоровых испытуемых. Оценивали эффективность прямой маскировки бытовых предметов двумя категориями социально значимых изображений – зданиями и лицами людей, а также анализировали влияние низкочастотной фильтрации (НЧФ) маски. Стимулами и масками служили черно-белые фотографии бытовых предметов, зданий и лиц. Время реакции (ВР) опознания стимула зависело от категории маски: при маскировке лицами значимый стимул опознавался быстрее, чем при маске-здании. Анализ распределения индивидуальных значений ВР показал, что категория маски влияла на среднее значение ВР, а ее пространственная частота – на дисперсию распределения, которая увеличивалась при НЧФ. Точность опознания не зависела от категории маски. НЧФ маски снижала точность опознания только при маске-здание. Амплитуды ранних компонентов зрительных вызванных потенциалов зависели и от категории, и от фильтрации маски. При маскировке лицами амплитуды затылочно-височного N50 и лобного Р50 были больше, чем при маске-здании, НЧФ маски приводила к их снижению. Среднелатентные компоненты N150 и Р200 в большей степени зависели от фильтрации маски, чем от ее категории. НЧФ маски приводила к увеличению N150 в лобных и затылочно-височных областях, а также снижению Р200 в затылочных. Влияние категории проявлялось только в увеличении амплитуды затылочно-височного N150 при маске-лице. Можно предположить, что меньшее маскирующее воздействие лиц связано с их способностью увеличивать пространственное внимание. НЧФ приводила к затруднению переработки информации о маскирующем изображении, что усложняло разделение значимой и незначимой информации.
Одним из важных условий адекватного поведенческого ответа является четкое разделение значимых и незначимых сигналов. Нередко в качестве таких нерелевантных сигналов выступают социально значимые стимулы, например, лица людей [1]. Лица являются уникальной категорией изображений, восприятие которой происходит автоматически и не зависит от их значимости для выполнения основной задачи, что позволяет предположить наличие механизма селективного внимания к этой категории стимулов [2, 3]. Известно, что лица опознаются быстрее и точнее изображений других категорий [1]. Увеличение пространственного внимания проявляется не только в опознании самих лиц, но и в ответах на последующие зрительные стимулы. Так, например, в задаче на пространственное внимание тестовый стимул-квадрат выделялся быстрее, если он появлялся в том месте экрана, где до этого предъявлялся лицевой стимул [4]. Другая категория социально-значимых стимулов нашей ежедневной жизни – это здания. В отличие от других категорий изображений, здания содержат в себе информацию о пространстве и воспринимаются как “метки на местности” (landmarks), играя важную роль в навигации [5]. Ранее было показано, что в модели прямой зрительной маскировке лица и здания практически не оказывают маскирующего воздействия на опознание значимых изображений [6, 7]. Такая обособленность этих двух категорий может быть связана с существованием селективных для них областей: это fusiform face area для лиц и parahippocampal place area для изображений сцен и зданий [8–10].
Традиционная теория переработки зрительной информации предполагает иерархическую организацию зрительной системы и восходящую передачу сигнала от низкоуровневых к высокоуровневым областям мозга [11]. Вместе с тем, многочисленные работы подчеркивают важность нисходящих влияний для опознания изображения [12–14]. Согласно современным модельным представлениям, низкочастотная составляющая информации о стимуле быстро передается по магноцеллюлярному пути в орбитофронтальную кору, где на ее основании генерируется гипотеза о категориальной принадлежности стимула [12, 13]. Далее орбитофронтальная кора оказывает нисходящее влияние на нижневисочную кору, входящую в вентральный путь передачи зрительной информации, активируя репрезентацию, соответствующую гипотезе, выдвинутой на основании низкочастотной составляющей орбитофронтальной корой [12]. Мишенями такого нисходящего влияния может быть не только нижневисочная кора, но и другие области мозга, включая первичную зрительную кору [15].
Одна из теорий, описывающих механизм зрительной маскировки – это теория двух каналов, предложенная B. Breitmeyer и H. Ogmen [16]. Согласно этой теории, в механизме прямой маскировки важную роль играет зрительная информация о маске, передающаяся по вентральному пути, которая тормозит последующую переработку значимого стимула [16]. В дальнейшем данная теория была дополнена предположением о том, что точный механизм, лежащий в основе маскирующего воздействия в каждом конкретном случае, будет зависеть от временны́х параметров предъявления маски и стимула [16].
Цель данной работы – на модели прямой маскировки сравнить влияние нерелевантных к выполняемой задаче изображений лиц и зданий на опознание значимых стимулов – предметов. Для выявления роли различных частотных составляющих зрительной информации о маскирующем изображении в механизме прямой маскировки в данной работе наряду с интактными использовали низкочастотно-фильтрованные маски, т.е. изображения, в которых высокие пространственные частоты были ослаблены за счет применения низкочастотного пространственного фильтра. Анализировали поведенческие и нейрофизиологические характеристики опознания стимулов – точность, время реакции и его дисперсию, амплитуду ранних и среднелатетных компонентов зрительных вызванных потенциалов.
МЕТОДИКА
Характеристика группы испытуемых. В исследовании участвовали 20 испытуемых (10 женщин и 10 мужчин) с нормальным или скорректированным до нормы зрением в возрасте от 20 до 36 лет (средний возраст 24.9 ± 1.0 лет). Каждый испытуемый участвовал в двух экспериментальных сериях: 1) опознание предметов при их маскировке интактными изображениями зданий и лиц и 2) опознание предметов при их маскировке НЧФ изображениями зданий и лиц.
Стимулы. В качестве целевых стимулов использовали черно-белые фотографии бытовых предметов, в качестве масок – черно-белые фотографии зданий и лиц людей с эмоционально-нейтральным выражением. Фотографии лиц брали из базы лицевых стимулов Университета Неймегена [17]. Изображения предметов и зданий выбирали из цветных фотографий, представленных в Интернете. Выбранные изображения вырезали из фона с помощью инструментов программы Adobe Photoshop CS4 и помещали по центру заранее созданного макета с белым фоном размером 800 × 600 пикселей. Размер самого изображения составлял 210 × 210 пикселей, разрешение – 72 точки на дюйм (72 dpi). Далее цветное изображение превращали в черно-белое переводя его из режима RGB в режим градаций серого. Для преобразования изображений в НЧФ стимулы к ним применяли фильтр Гаусса с установленным значением размытия в размере 3-х пикселей на стимульное изображение. Размер изображения на экране монитора – 8–10 см по ширине и по высоте, что в условиях нашего эксперимента (120 см от экрана до глаз испытуемого) составляло 4–5 угл. град.
Для каждой из категорий стимулов (сумка, чашка, утюг и т.п.) выбирали по 4 разных изображения объекта. Для каждой из двух экспериментальных серий создавали 48 изображений: 32 маски (16 зданий и 16 лиц) и 16 стимулов. Из этих масок и стимулов составляли 96 уникальных пар маска-стимул, каждую из которых в ходе серии предъявляли по одному разу. На рис. 1, А, Б приведены примеры этих изображений.
Рис. 1.
Условия эксперимента. А – примеры стимулов четырех категорий, использованных в эксперименте. Б – примеры НЧФ и нефильтрованных маскирующих изображений лиц и зданий. В – временнáя схема эксперимента.

Ход эксперимента. Во время эксперимента испытуемый сидел в удобном кресле в затемненной (освещенность на уровне глаз испытуемого – 3 лк.) и звукозаглушенной комнате на расстоянии 120 см от экрана монитора Dell E1911c (диагональ 17 дюймов, частота обновления 60 Гц). Предъявление стимулов и регистрацию точности опознания и времени реакции (ВР) проводили с помощью программы E-Рrime 2.0 (Psychology Software Tools, Inc., США) с выносной клавиатурой Serial Response Box. Исследование проводили в дневное время (с 10 до 17 ч).
Временнáя схема эксперимента представлена на рис. 1, В. Длительность предъявления маскирующего изображения – 100 мс, сразу после него на 100 мс предъявляли целевой стимул. Межстимульный интервал изменяли в случайном порядке от 2.8 до 3.2 с, во время него в центре экрана находилась фиксационная точка. При опознании стимула испытуемый нажимал на одну из клавиш выносной клавиатуры Serial Response Box. Испытуемых просили отвечать быстро, но так, чтобы избегать большого количества ошибок. Для каждого испытуемого рассчитывали вероятность правильных ответов, или точность (в %), и среднее время реакции (ВР, мс) для правильных ответов.
Эксперимент начинали с тренировочной серии для выработки моторного навыка, во время которой на экране в случайном порядке на 100 мс, вспыхивали цифры от 1 до 4 (каждая по 12 предъявлений). При появлении цифры испытуемый должен был нажать на соответствующую ей кнопку клавиатуры.
Основную часть эксперимента начинали с появления на экране инструкции. Например, для серии с НЧФ масками она выглядела так: “На экране в случайном порядке будут появляться изображения предметов. Эксперимент будет состоять из двух частей. Первая – тренировочная. Вторая – основная. Ваша задача – опознать предмет и нажать на соответствующую клавишу. Во время эксперимента, пожалуйста, смотрите в центр экрана. Постарайтесь не моргать во время предъявления стимула. Моргать можно после нажатия кнопки. При появлении изображения туфли нажмите на цифру 1. При появлении изображения лампы нажмите на цифру 2. При появлении изображения зонта нажмите на цифру 3. При появлении изображения кувшина нажмите на цифру 4”. Для серии с интактными масками инструкция была аналогичной, с поправкой на используемые целевые стимулы. Каждую экспериментальную серию предваряли тренировкой, в ходе которой целевые стимулы предъявляли без маскировки. Тренировочная часть была необходима для создания твердой ассоциации между стимулом и нажимаемой в ответ на него клавишей выносной клавиатуры. В тренировочной части длительность стимула составляла 100 мс, межстимульного интервала – 3000 мс, каждое целевое изображение предъявлялось 1 раз, всего 16 предъявлений. После окончания тренировки на экране появлялась надпись “Основная часть”, после чего начиналось предъявление замаскированных целевых изображений (параметры предъявления изображений в основной части описаны выше). Длительность каждой экспериментальной серии около 7 мин.
Регистрация и анализ вызванных потенциалов. Регистрацию электроэнцефалограммы (ЭЭГ) проводили на 128-канальном электроэнцефалографе фирмы Geodesics (Electrical Geodesics Inc., США) с помощью шлема Hydro Cel Geodesic Sensor Net (Electrical Geodesics Inc., США). Частота квантования сигнала 500 Гц. Сопротивление электродов не превышало 50 кОм. При записи в качестве индифферентного электрода использовали вертекс (Cz). При off-line обработке записи в качестве референтного выбирали усредненный электрод. Данные ЭЭГ обрабатывали off-line в программе NetStation 4.5.4 (Electrical Geodesics Inc., США). Предварительно фильтрованные (0.5–45 Гц) записи ЭЭГ каждого испытуемого сегментировали на отрезки длительностью 1100 мс, содержащие интервалы 300 мс до предъявления маски и 800 мс после. Далее программно исключали реализации с выраженными глазными движениями, а при дальнейшем визуальном анализе исключали фрагменты с двигательными и электромиографическими артефактами. После этого для каждого испытуемого по безартефактным отрезкам ЭЭГ (от 25 до 45 реализаций, в среднем 36.5 ± 0.6) проводили усреднение вызванных потенциалов (ВП) на стимул для правильных ответов для разных категорий маскирующих изображений. Коррекцию базовой линии проводили по отрезку ЭЭГ 300 мс до предъявления маски. В результате для каждого испытуемого получили по 4 усредненных ВП: 1) маскировка интактными изображениями зданий, 2) маскировка интактными лицами, 3) маскировка НЧФ изображениями зданий и 4) маскировка НЧФ лицами.
Амплитуду выбранных для анализа компонентов индивидуальных вызванных потенциалов измеряли в программе NetStation 4.5.4. Замеряли адаптивный минимум или максимум во временнóм окне компонента, а именно средние значения амплитуды в интервале длиной 4 мс, соответствующем по 2 мс до и после пика компонента. Анализировали амплитуду компонентов ВП, усредненную по кластерам электродов. Усреднение амплитуды проводили в симметричных лобных (в левом полушарии сенсоры № 18, 21, 22, в правом – № 9, 10, 14), височных (в левом полушарии – № 58, 59, 65, в правом – № 90, 91, 96) и затылочных (в левом полушарии – № 66, 70, 71, в правом – № 76, 83, 84) кластерах электродов.
Анализ распределения времени реакции. Согласно литературным данным, распределение ВР отличается от нормального и подчиняется экс-Гауссову распределению (ex-Gaussian distribution) [18, 19]. Типичное распределение ВР ассиметрично и сдвинуто вправо, т.е. имеет “хвост” больших значений ВР. Экс-Гауссова функция рассматривает распределение ВР как сочетание двух распределений: Гауссова (нормального), описывающего центральную часть распределения, и экспоненциального, характеризующего “хвост” больших значений. Для анализа экс-Гауссова распределения используют три параметра: µ и σ, описывающих среднее значение и стандартное отклонение нормальной части распределения, соответственно, и τ, характеризующего среднее значение и стандартное отклонение экспоненциальной части [18]. Для вычисления параметров распределение ВР использовали метод оценки максимального правдоподобия (continuous maximum likelihood estimation). Вычисление параметров µ, σ и τ проводили в программной среде RStudio 1.1.453 (RStudio, Inc., США) с помощью скрипта, написанного A. Heathcote [20].
При статистической обработке данных использовали дисперсионный анализ – метод повторных измерений, ANOVA RM. При анализе поведенческих данных учитывали влияние факторов ФИЛЬТРАЦИЯ (2 уровня: интактная или НЧФ маска) и КАТЕГОРИЯ (2 уровня: маска-здание или маска-лицо). При анализе амплитуд компонентов ВП учитывали факторы ФИЛЬТРАЦИЯ, КАТЕГОРИЯ и ПОЛУШАРИЕ (2 уровня). Результаты ANOVA скорректированы поправкой Гринхауса–Гейссера. При внутригрупповых post-hoc сопоставлениях использовали тест Ньюмена–Кейлса на множественные сравнения. Статистический анализ проводили в программах SPSS 20.0 и Statistica 10.0.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Анализ поведения. Проведен дисперсионный анализ ANOVA RM ВР и точности опознания стимула с факторами КАТЕГОРИЯ и ФИЛЬТРАЦИЯ.
Для ВР выявлен значимый эффект фактора КАТЕГОРИЯ F(1, 19) = 11.85; p < 0.01. На рис. 2, А приведены средние значения ВР при различных типах маскирующих изображений. Видно, что при маскировке изображениями лиц ВР меньше, чем при маске-здании.
Рис. 2.
Влияние категориальной принадлежности и низкочастотной фильтрации маски на ВР, точность опознания и распределение ВР. А – время реакции (мс) в разных условиях эксперимента. Б – вероятность правильного ответа (%). Приведены средние по группе значения и ошибка среднего. Достоверность различий * – p < 0.05 (критерий Ньюмана−Кейлса). В – график распределения индивидуальных значений ВР при использовании разных маскирующих изображений. На А–В цифрами обозначены варианты маскирующего изображения: 1 – нефильтрованное изображение здания, 2 – НЧФ здание, 3 – нефильтрованное изображение лица, 4 – НЧФ лицо.
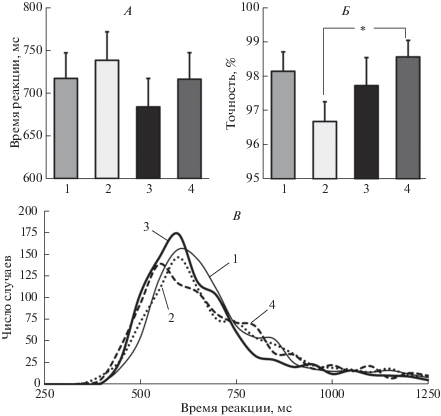
Для точности опознания выявлено значимое взаимодействие КАТЕГОРИЯ × ФИЛЬТРАЦИЯ F(1, 19) = 6.07, p < 0.05. На рис. 2, Б видно, что для изображений зданий точность опознания выше при использовании интактных изображений по сравнению с низкочастотно-фильтрованными. В случае лиц точность опознания стимула не зависела от фильтрации маски. Влияние категории маски выявлено при сравнении НЧФ изображений: точность выше для лиц по сравнению со зданиями (p < 0.05).
На рис. 2, В приведены графики распределения ВР индивидуальных ответов испытуемых при разных типах маскировки. Распределения ВР во всех случаях не подчиняется закону нормального распределения (p < 0.0005, критерий Шапиро−Уилка). Для дополнительного анализа сочетанного эффекта категории маски и ее фильтрации проведено сравнение параметров экс-Гауссова распределения ВР µ и σ, отражающих среднее значение и стандартное отклонение Гауссовой функции, соответственно, и τ, отражающей среднее значение и стандартное отклонение экспоненциальной функции. Полученные результаты приведены в табл. 1. Проведен дисперсионный анализ ANOVA RM индивидуальных параметров распределения ВР.
Таблица 1.
Параметры экс-Гауссова распределения индивидуальных значений времени реакции
| Маскирующий стимул | Фильтрация | µ | σ | τ |
|---|---|---|---|---|
| Дом | Интактное изображение | 593.9 (23.3) | 154.5 (16.3) | 122.3 (11.6) |
| Лицо | 577.6 (23.5) | 141.8 (18.5) | 105.4 (11.9) | |
| Дом | Низкочастотно-фильтрованное изображение | 604.0 (24.0) | 187.0 (18.5) | 133.3 (15.7) |
| Лицо | 574.2 (24.7) | 177.1 (16.8) | 141.1 (14.3) |
Для µ выявлен значимый эффект фактора КАТЕГОРИЯ F(1, 19) = 14.79, p < 0.001. Параметр µ больше при маскировке зданиями. И в случае интактных, и в случае низкочастотно-фильтрованных масок при маскировке зданиями распределение сдвинуто вправо в область больших значений по сравнению с маскировкой лицами (рис. 2, В).
Для параметра σ обнаружен достоверный эффект фактора ФИЛЬТРАЦИЯ F(1, 19) = 8.93, p < 0.01. Низкочастотная фильтрация увеличивает дисперсию той части экс-Гауссова распределения, которая описывается Гауссовой функцией. Эта закономерность выявляется как для масок-зданий, так и для масок-лиц.
Для параметра τ также обнаружен достоверный эффект фактора ФИЛЬТРАЦИЯ F(1, 19) = = 5.24, p < 0.05. Низкочастотная фильтрация маски приводит к увеличению τ. Этот параметр характеризует экспоненциальную часть распределения. Большая его величина при НЧФ отражает увеличение “хвоста” распределения в области больших значений (рис. 2, В).
Анализ вызванных потенциалов каудальных отделов коры. Компонент N50 регистрировали в задних отделах коры со средним значением латентности пика 52 ± 14 мс. Проведен дисперсионный анализ ANOVA RM с факторами ОБЛАСТЬ (2 уровня: затылочная и височная), ФИЛЬТРАЦИЯ, КАТЕГОРИЯ и ПОЛУШАРИЕ. Полученные результаты приведены в табл. 2.
Таблица 2.
Результаты дисперсионного анализа ANOVA RM амплитуды компонентов зрительных вызванных потенциалов (приведены только значимые эффекты)
| Схема ANOVA | Компонент | Область | Факторы и их взаимодействия | df | F | Значение p |
|---|---|---|---|---|---|---|
| ОБЛАСТЬ, ФИЛЬТРАЦИЯ, КАТЕГОРИЯ, ПОЛУШАРИЕ | N50 | Затылочная и височная | ОБЛАСТЬ | 1, 19 | 14.93 | 0.001 |
| ФИЛЬТРАЦИЯ | 1, 19 | 28.31 | 0.0005 | |||
| КАТЕГОРИЯ | 1, 19 | 31.84 | 0.0005 | |||
| ОБЛАСТЬ × ФИЛЬТРАЦИЯ | 1, 19 | 6.85 | 0.017 | |||
| ОБЛАСТЬ × КАТЕГОРИЯ | 1, 19 | 24.12 | 0.0005 | |||
| ОБЛАСТЬ × ФИЛЬТРАЦИЯ × × КАТЕГОРИЯ | 1, 19 | 6.24 | 0.022 | |||
| ОБЛАСТЬ × КАТЕГОРИЯ × × ПОЛУШАРИЕ | 1, 19 | 7.27 | 0.014 | |||
| ФИЛЬТРАЦИЯ, МАСКА, ПОЛУШАРИЕ | N150 | Затылочная | ФИЛЬТРАЦИЯ | 1, 19 | 7.21 | 0.015 |
| КАТЕГОРИЯ | 1, 19 | 7.05 | 0.016 | |||
| Височная | ФИЛЬТРАЦИЯ | 1, 19 | 12.84 | 0.002 | ||
| КАТЕГОРИЯ | 1, 19 | 5.94 | 0.025 | |||
| КАТЕГОРИЯ × ПОЛУШАРИЕ | 1, 19 | 4.91 | 0.039 | |||
| Р200 | Затылочная | ФИЛЬТРАЦИЯ | 1, 19 | 20.60 | 0.0005 | |
| ФИЛЬТРАЦИЯ × КАТЕГОРИЯ | 1, 19 | 6.07 | 0.023 | |||
| Р50 | Лобная | ФИЛЬТРАЦИЯ | 1, 19 | 4.59 | 0.045 | |
| КАТЕГОРИЯ | 1, 19 | 67.63 | 0.0005 | |||
| N150 | ФИЛЬТРАЦИЯ | 1, 19 | 8.22 | 0.01 |
Дисперсионный анализ выявил достоверные эффекты факторов ОБЛАСТЬ (p < 0.001), ФИЛЬТРАЦИЯ (p < 0.0005) и КАТЕГОРИЯ (p < 0.0005). Также были выявлены взаимодействия ОБЛАСТЬ × ФИЛЬТРАЦИЯ (p < 0.05), ОБЛАСТЬ × × КАТЕГОРИЯ (p < 0.0005), ОБЛАСТЬ × ФИЛЬТРАЦИЯ × КАТЕГОРИЯ (p < 0.05) и ОБЛАСТЬ × × КАТЕГОРИЯ × ПОЛУШАРИЕ (p < 0.05).
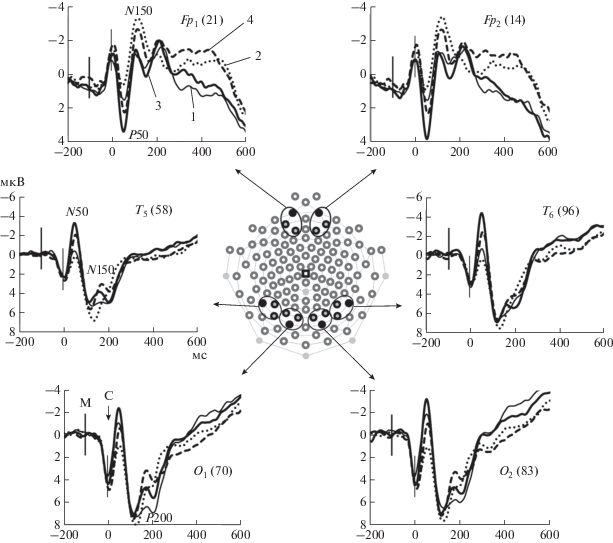
Как видно на рис. 3, амплитуда компонента N50 выше в височных областях по сравнению с затылочными. Однако достоверные отличия между областями наблюдаются только в правом полушарии при использовании в качестве масок лиц (p < 0.01 для интактных и p < 0.05 для НЧФ изображений).
Рис. 3.
Зрительные ВП затылочных О1 и О2 (электроды 70 и 83 соответственно), височных Т5 и Т6 (58 и 96) и лобных Fp1 и Fp2 (14 и 21) отведений при маскировке нефильтрованными и НЧФ изображениями зданий и лиц. В центре рисунка приведена схема расположения 128 регистрирующих электродов на поверхности головы. Черный квадрат – индифферентный электрод (Cz). Черным обведены электроды, принадлежащие лобному, височному и затылочному кластерам. Закрашены электроды, ВП которых приведены на графиках. По оси абсцисс – время (мс), по оси ординат – амплитуда (мкВ). Варианты маскирующих изображений: 1 – нефильтрованные здания, 2 – НЧФ здания, 3 – нефильтрованные лица, 4 – НЧФ лица. Вертикальные линии – моменты предъявления маски (толстая линия) и стимула (тонкая линия). Обозначены пики компонентов N50, P50, N150 и Р200.
Амплитуда N50 зависела от категориальной принадлежности маски: для изображений домов амплитуда меньше, чем для лиц (0.001 < p < 0.05) (рис. 4, А, Б). Данная зависимость более выражена в височных областях, где она выявлена как для интактных, так и для НЧФ масок (0.001 < p < 0.05). В затылочном кластере post-hoc выявил влияние категории маски только для НЧФ изображений в правом полушарии (p < 0.05) (рис. 4, Б).
Рис. 4.
Влияние категории и НЧФ маскирующего изображения на амплитуду компонентов N50, N150 и Р200 в затылочных и височных областях. А, Б – амплитуда компонента N50. В, Г – амплитуда компонента N150. Д, Е – амплитуда компонента Р200. Амплитуды усреднены по кластерам электродов. Верхний ряд – левое полушарие (А, В, Д), нижний – правое полушарие (Б, Г, Д). Цифрами обозначены варианты маски: 1 – нефильтрованные здания, 2 – НЧФ здания, 3 – нефильтрованные лица, 4 – НЧФ лица. Приведены средние по группе значения и ошибка среднего. * – p < 0.05, ** – p < 0.01 и *** – p < 0.001 (критерий Ньюмана−Кейлса).
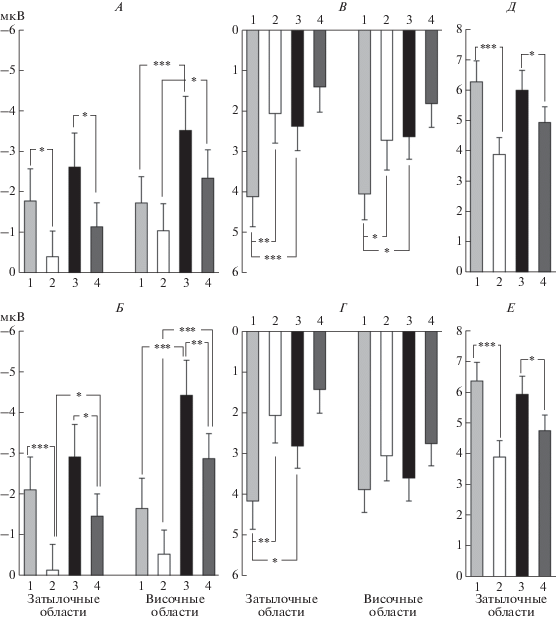
Как видно на рис. 4, А, Б, амплитуда данного компонента достоверно больше для интактных масок по сравнению с НЧФ (0.001 < p < 0.05). Фильтрация маски оказывает большее влияние на амплитуду компонента в затылочном кластере электродов по сравнению с височным. В затылочных областях достоверные отличия между маскировкой интактными и НЧФ изображениями наблюдаются для всех категорий масок (0.001 < p < < 0.05), в то время как в височных областях только для лиц в правом полушарии (p < 0.01).
Компонент N150 регистрировали в задних отделах коры со средним значением латентности пика 182 ± 37 мс. Амплитуду этого компонента проанализировали в затылочном и височном кластерах электродов с использованием дисперсионного анализа ANOVA RM (ФИЛЬТРАЦИЯ, КАТЕГОРИЯ, ПОЛУШАРИЕ). Полученные результаты приведены в табл. 2.
В затылочном кластере дисперсионный анализ выявил достоверные эффекты факторов ФИЛЬТРАЦИЯ (p < 0.05) и КАТЕГОРИЯ (p < 0.05). Как видно на рис. 4, В, Г, при маскировке лицами амплитуда больше, чем при маскировке зданиями, но достоверные отличия получены только для интактных изображений (0.001 < p < 0.05). Амплитуда больше для НЧФ маскирующих изображений, но достоверные отличия наблюдаются только при маскировке зданиями (p < 0.01).
В височном кластере были выявлены достоверные эффекты фактора ФИЛЬТРАЦИЯ (p < < 0.01), КАТЕГОРИЯ (p < 0.05) и взаимодействие КАТЕГОРИЯ × ПОЛУШАРИЕ (p < 0.05). В левом полушарии амплитуда N150 достоверно выше при интактной маске-лице по сравнению с нефильтрованными зданиями (p < 0.05). Так же, как в затылочном, в височном кластере НЧФ маски приводит к увеличению амплитуды данного компонента, достоверные отличия наблюдаются только в левом полушарии для маски-здания (p < 0.05) (рис. 4, В).
Компонент Р 200 регистрировали в затылочных отделах коры со средним значением латентности пика 225 ± 43 мс.
Дисперсионный анализ ANOVA RM (ФИЛЬТРАЦИЯ, КАТЕГОРИЯ и ПОЛУШАРИЕ) выявил достоверный эффект фактора ФИЛЬТРАЦИЯ (p < 0.0005) и взаимодействие ФИЛЬТРАЦИЯ × КАТЕГОРИЯ (p < 0.05). Как видно на рис. 4, Д, Е, низкочастотная фильтрация маски приводила к снижению амплитуды данного компонента, более выраженному для маски-здания (p < 0.0005) по сравнению с маской-лицом (p < 0.05).
Анализ вызванных потенциалов лобных отделов коры. Компонент Р50 регистрировали в передних отделах коры со средним значением латентности пика 44 ± 25 мс.
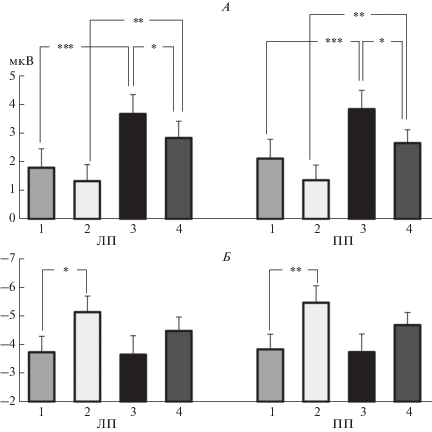
Дисперсионный анализ ANOVA RM (ФИЛЬТРАЦИЯ, КАТЕГОРИЯ, ПОЛУШАРИЕ) выявил достоверные эффекты факторов ФИЛЬТРАЦИЯ (p < 0.05) и КАТЕГОРИЯ (p < 0.0005) (табл. 2). Как видно на рис. 5, А, независимо от фильтрации маски, амплитуда компонента больше при использовании в качестве маскирующих изображений лиц по сравнению со зданиями (p < 0.001 для интактных и p < 0.01 для НЧФ изображений). НЧФ маски приводила к снижению амплитуды этого компонента ВП, которое было заметнее при маскировке лицами (p < 0.05).
Рис. 5.
Влияние категории и НЧФ маскирующего изображения на амплитуду компонентов Р50 и N150 в лобных областях. А – амплитуда компонента Р50. Б – амплитуда компонента N150. Амплитуды усреднены по кластерам электродов. ЛП – левое полушарие, ПП – правое полушарие. Остальные обозначения см. рис. 4.
Компонент N150 отчетливо регистрировали в передних отделах коры со средним значением латентности пика 121 ± 19 мс.
Дисперсионный анализ ANOVA RM (ФИЛЬТРАЦИЯ, КАТЕГОРИЯ, ПОЛУШАРИЕ) выявил достоверный эффект фактора ФИЛЬТРАЦИЯ (p < 0.01) (табл. 2). НЧФ маскирующего изображения приводила к увеличению амплитуды данного компонента ВП (рис. 5, Б). Более выраженным это увеличение было при маскировке зданиями (p < 0.05 в левом и p < 0.01 в правом полушарии).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Цель данной работы – на модели прямой маскировки сравнить влияние нерелевантных к выполняемой задаче изображений лиц и зданий на опознание значимых стимулов – предметов. Для выявления роли различных частотных составляющих зрительной информации о маскирующем изображении в данной работе наряду с интактными использовали НЧФ маски. В работе получены следующие результаты:
1) В модели прямой маскировки нерелевантные в выполняемой задаче изображения зданий и лиц не оказывали значимого влияния на точность опознания предметов, но различались по влиянию на ВР, которое меньше при маске-лицо, чем при маске-здание.
2) НЧФ маски приводила к увеличению дисперсии распределения индивидуальных значений ВР, а также в случае масок-зданий – к снижению точности опознания.
3) Амплитуда ранних компонентов зрительных ВП зависела от категории и пространственной частоты маски. При маскировке лицами амплитуда затылочно-височного компонента N50 и лобного компонента Р50 больше, чем при маске-здании. НЧФ маски приводит к снижению амплитуды N50 и Р50.
4) Амплитуда среднелатентных компонентов ВП в большей степени зависелa от пространственной частоты маски, чем от ее категории. НЧФ маски приводила к росту негативности N150 в лобных и затылочно-височных областях и снижению Р200 в затылочных. Категория маски влияла на амплитуду затылочно-височного компонента N150, которая выше при маске-лице.
Полученный в работе результат о меньшем ВР опознания значимого стимула при маскировке лицами по сравнению со зданиями соответствует результатам, полученным другими авторами [21–23], и нашим более ранним работам [6, 7]. Мы дополнили эти данные результатами анализа индивидуальных значений ВР. Обнаружено, что категория маски влияет на параметр µ, отражающий среднее значение Гауссовой части распределения, и связанный с перцептивным звеном переработки сигнала [24, 25]. Тот факт, что при маскировке лицами µ меньше, чем при зданиях, свидетельствует о том, что категория маски влияет на этап перцепции значимого стимула, а не на более позднюю когнитивную стадию [24, 25]. Показатели σ и τ, характеризующие дисперсию Гауссовой части распределения, а также среднее значение и дисперсию его экспоненциальной части, зависели от частотных характеристик маски. Значения этих показателей возрастали при использовании НЧФ масок, что согласно работам M. Sigman, S. Dehaene [24] и H. Banno, J. Saiki [25], отражает увеличение нагрузки на когнитивное звено переработки информации.
Точность опознания не зависела от категории маски. Влияние НЧФ маски показано только для маски-здания. Отсутствие эффекта фильтрации для лиц соответствует факту, что лицо увеличивает пространственное внимание, независимо от его пространственно-частотных характеристик [26].
Зрительные ВП в использованной нами модели прямой маскировки отражают результат интерференции переработки маскирующего и следующего за ним через 100 мс значимого изображений. Возникает закономерный вопрос, как эти два одновременно протекающих процесса отражаются в компонентах ВП. Мы исходим из предположения, что ранние компоненты ВП, а именно, N50 в задних отделах коры и Р50 – в передних, связаны с процессами переработки маскирующего изображения [27]. Их пики регистрируются через 150 мс после предъявления маски и всего лишь через 50 мс после появления релевантного стимула. Более поздние компоненты (N150 и P200) связаны с переработкой значимого стимула, на которую влияет “преднастройка” нейронов, возникшая ранее при анализе маскирующего изображения [27].
На амплитуду ранних компонентов ВП оказывала существенное влияние категория маски. Амплитуда N50 в затылочных и височных областях коры была больше при маске-лице, чем при маске-здании. Влияние категории маски на амплитуду N50 более отчетливо в височных областях, где различия между категориями показаны и для нефильтрованных, и для НЧФ изображений. Более высокая амплитуда компонента при маске-лице объясняется тем, что он регистрируется через 150 мс после предъявления маски, т.е., во временнóм окне лицо-специфического компонента N170, отражающего кодирование индивидуальных черт лица [28, 29].
Более высокая амплитуда при маске-лице показана и для компонента Р50 в лобных областях. Компонент Р50 отражает ранний этап переработки информации о маске, что позволяет сравнить его с позитивностью Р200, которая регистрируется 150−250 мс после предъявления изображения [30, 31]. Показано, что амплитуда этого компонента увеличивается при предъявлении релевантных стимулов, новых необычных изображений и связана с ростом экзогенного внимания [30–33]. Лица являются социально-значимой категорией изображений, к которым в эволюции сформировался механизм увеличения пространственного внимания даже при параллельном выполнении других перцептивных или когнитивных задач [34]. Следовательно, увеличение амплитуды Р50 при маскировке лицами может отражать усиленное внимание к данной категории изображений.
Амплитуда ранних компонентов ВП также зависела от пространственной частоты маски. НЧФ маски приводила к снижению амплитуды N50 в каудальных отделах коры, более заметному в затылочной области, где достоверные отличия выявлены для обеих категорий маскирующих изображений. В ряде работ продемонстрированно, что чем глубже переработка предшествующего незначимого изображения, т.е., чем полнее извлечена зрительная информация о нем, тем больше амплитуда ранней каудальной негативности [35, 36]. Таким образом, можно предположить, что в нашей работе компонент N50 отражает глубину переработки информации о маскирующем изображении, которая была больше в случае нефильтрованных масок, так как они содержат более полную зрительную информацию об изображении, чем НЧФ маски.
НЧФ приводила к снижению амплитуды ранней позитивности Р50 в лобных отделах коры. Связь амплитуды этого компонента и частотных характеристик дистрактора продемонстрирована в работе [33]. Авторы, меняя яркость и цвет дистрактора, показали, что амплитуда данного компонента будет меньше, если использовать дистрактор, обрабатывающийся преимущественно в магноцеллюлярном пути [33], что соответствует в нашей работе НЧФ маске. Согласно H. Gibbons et al. [35], позитивность в этом временнóм диапазоне (около 200 мс после прайма) связана с процессом нисходящего торможения информации о прайме. То есть, более высокая амплитуда Р50 при использовании интактных масок по сравнению с НЧФ может говорить о более успешном торможении переработки нерелевантной к выполняемой задаче маски, что приводит к улучшению опознания.
Анализ среднелатентных компонентов ВП показал, что на их амплитуду в большей степени влияла фильтрация маски, чем ее категориальная принадлежность. НЧФ маски вызывала увеличение N150 в лобных и каудальных отделах и снижение Р200 в затылочных. В работе C. Peyrin et al. испытуемым последовательно предъявляли два изображения, подвергавшиеся НЧФ и ВЧФ [15]. Показано, что предшествующее низкочастотное изображение оказывало выраженное модулирующее влияние на переработку значимого стимула, что выражалось в большей активации височно-теменных и левой лобной областей [15]. Можно предположить, что большая амплитуда компонента N150 в лобных отведениях при маскировке НЧФ изображениями отражает их модулирующее влияние на переработку значимого стимула. В работе T. Mu и S. Li при предъявлении испытуемым наложенные друг на друга двух разных изображения (ВЧФ и НЧФ) обнаружено, что ухудшение опознания при семантической конгруэнтности этих двух стимулов сопровождалось увеличением негативность N1 (латентность 122 мс) в лобных отведениях [37]. Данный эффект они связали с процессом интерференции низкочастотной и высокочастотной информации [37]. С учетом этих данных, наблюдаемое в нашей работе увеличение амплитуды лобного N150 при использовании НЧФ масок также может быть связано с интерференцией в лобных отделах информации о маске и значимом изображении.
В нашем исследовании НЧФ маски приводила к увеличению амплитуды компонента N150 в каудальных отделах коры. Похожие эффекты описаны в работе C. Peyrin et al. [15]. Авторы показали, что если низкочастотное изображение предшествовало высокочастотному, то во временнóм интервале негативности N150 (176–204 мс) в ВП на значимый стимул наблюдали большую активацию затылочных отделов коры, что отсутствовало в противоположной ситуации, когда высокочастотный стимул предшествовал низкочастотному. Авторы связывают это увеличение с влияниями вышележащих областей, активированных предшествующим низкочастотным изображением [15]. Сходный механизм может лежать в основе обнаруженного в нашей работе эффекта увеличения каудальной негативности N150 при НЧФ маски.
Напротив, амплитуда среднелатентного каудального компонента Р200 была больше при маскировке интактными изображениями по сравнению с НЧФ. Для обсуждения этого результата представляется интересным привлечь литературу о компоненте Рd (distractor positivity). Pd регистрируют во временнóм окне 200–350 мс при одновременном предъявлении стимула и дистрактора в контралатеральном по отношению к дистрактору полушарии и связывают с подавлением переработки зрительной информации о дистракторе [38–41]. При этом, чем больше внимания захватывает дистрактор, тем больше амплитуда Pd [40, 41], следовательно, увеличение амплитуды Pd свидетельствует об успешном подавлении переработки информации о дистракторе. Обнаруженная в нашем исследовании большая амплитуда позитивности при маскировке нефильтрованными изображениями может быть свидетельством более успешного подавления, в этом случае информации о маске, чем при использовании НЧФ изображений. В результате интактные маски оказывают меньшее маскирующее воздействие на опознание значимого стимула. Подобная связь выраженности маскирующего эффекта с амплитудой позитивности в сходном временнóм интервале (200–260 мс) показана в работе M. Del Zotto и A.J. Pegna при последовательном предъявлении стимула и маски [42].
Влияние категории маски было выявлено только в случае компонента N150 в каудальных отделах коры. При использовании в качестве масок изображений лиц амплитуда N150 была больше по сравнению с маскировкой зданиями. Показано, что лица увеличивают пространственное внимание испытуемого, облегчая опознание следующих за ними зрительных стимулов [4, 6, 7]. Можно предположить, что увеличение пространственного внимания при маске-лице приводило к росту амплитуды N150 на последующий значимый стимул.
ЗАКЛЮЧЕНИЕ
В работе получены новые данные о влиянии семантических и физических характеристик маски на эффективность прямой маскировки сложных изображений. Категориальная принадлежность маски влияла на ВР опознания изображения и, как показал анализ компонентов ВП, на ранний этап переработки зрительной информации. Авторы предполагают, что маска-лицо увеличивает пространственное внимание испытуемого, что облегчает переработку последующего значимого стимула. Низкочастотная фильтрация маски увеличивает дисперсию распределения индивидуальных ВР, что, предположительно, связано с возросшей нагрузкой на когнитивное звено переработки зрительной информации. При анализе ВП было выявлено, что фильтрация маски влияла на амплитудные характеристики как ранних, так и среднелатентных компонентов. Низкочастотная фильтрация маски ухудшала ее переработку, что затрудняло разделение значимой и незначимой зрительной информации. НЧФ в меньшей степени влияла на эффективность маскировки лицами, чем зданиями. НЧФ маски-здания приводила к снижению точности опознания значимого стимула, в случае маски-лица такого ухудшения при НЧФ не наблюдали. Анализ среднелатентных компонентов зрительного ВП показал, что НЧФ маски больше влияла на их амплитуду в случае зданий, чем в случае лиц. Таким образом, даже НЧФ лица увеличивали пространственное внимание испытуемого, что приводило к успешному торможению переработки этой категории маскирующих изображений и уменьшению интерференции значимой и незначимой зрительной информации.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены локальным биоэтическим комитетом Института высшей нервной деятельности и нейрофизиологии РАН (Москва).
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. Работа выполнена по государственному заданию ИВНД и НФ РАН.
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Guyader N., Chauvin A., Boucart M., Peyrin C. Do low spatial frequencies explain the extremely fast saccades towards human faces? // Vis. Res. 2017. V. 133. P. 100.
Lavie N., Ro T., Russell C. The role of perceptual load in processing distractor faces // Psychol. Sci. 2003. V. 14. № 5. P. 510.
Neumann M.F., Mohamed T.N., Schweinberger S.R. Face and object encoding under perceptual load: ERP evidence // NeuroImage. 2011. V. 54. № 4. P. 3021.
Bindemann M., Burton A.M., Langton S.R.H. et al. The control of attention to faces // J. Vision. 2007. V. 7. № 10. P. 1.
Sala J.B., Rama P., Courtney S.M. Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory // Neuropsychol. 2003. V. 41. № 3. P. 341.
Герасименко Н.Ю., Славуцкая А.В., Калинин С.А. и др. Опознание зрительных образов в условиях их прямой маскировки. Влияние категориальной близости значимого и маскирующих изображений // Журн. высш. нерв. деятельности. 2013. Т. 63. № 4. С. 419.
Герасименко Н.Ю., Славуцкая А.В., Калинин С.А. и др. Зрительное опознание в условиях прямой маскировки. Сравнительный анализ маскирующих эффектов лицевых и нелицевых стимулов // Журн. высш. нерв. деятельности. 2017. Т. 67. № 2. С. 180.
Downing P.E., Chan A.W.Y., Peelen M.V. et al. Domain specificity in visual cortex // Cereb. Cortex. 2005. V. 16. № 10. P. 1453.
Bunzeck N., Schutze H., Duzel E. Category-specific organization of prefrontal response-facilitation during priming // Neuropsychol. 2006. V. 44. № 10. P. 1765.
Grill-Spector K., Weiner K.S. The functional architecture of the ventral temporal cortex and its role in categorization // Nat. Rev. Neurosci. 2014. V. 15. № 8. P. 536.
Grill-Spector K., Malach R. The human visual cortex // Annu. Rev. Neurosci. 2004. V. 27. P. 649.
Bar M., Kassam K.S., Ghuman A.S. et al. Top-down facilitation of visual recognition // PNAS. 2006. V. 103. № 8. P. 449.
Rokszin A.A., Győri-Dani D., Nyúl L.G., Csifcsák G. Electrophysiological correlates of top-down effects facilitating natural image categorization are disrupted by the attenuation of low spatial frequency information // Int. J. Psychophys. 2016. V. 100. P. 19.
Dima D.C., Perry G., Singh K.D. Spatial frequency supports the emergence of categorical representations in visual cortex during natural scene perception // NeuroImage. 2018. V. 179. P. 102.
Peyrin C., Michel C.M., Schwartz S. et al. The neural substrates and timing of top–down processes during coarse-to-fine categorization of visual scenes: a combined fMRI and ERP study // J. Cogn. Neurosci. 2010. V. 22. № 12. P. 2768.
Breitmeyer B., Ogmen H. Visual masking. Time slices through conscious and unconscious vision. USA, N.Y.: Oxford University Press, 2006. 369 p.
Langner O., Dotsch R., Bijlstra G. et al. Presentation and validation of the Radboud Faces Database // Cognition and Emotion. 2010. V. 24. № 8. P. 1377.
Balota D.A., Yap M.J., Cortese M.J., Watson J.M. Beyond mean response latency: Response time distributional analyses of semantic priming // J. Mem. Lang. 2008. V. 59. № 4. P. 495.
Roelofs A., Piai V. Distributional analysis of semantic interference in picture naming // Q. J. Exp. Psychol. 2017. V. 70. № 4. P. 782.
Heathcote A. Fitting Wald and ex-Wald distributions to response time data: An example using functions for the S-PLUS package // Behav. Res. Methods. Instrum. Comput. 2004. V. 36. № 4. P. 678.
Mogg K., Bradley B.P. Some methodological issues in assessing attentional biases for threatening faces in anxiety: a replication study using a modified version of the probe detection task // Behav. Research Therapy. 1999. V. 37. № 6. P. 595.
Pourtois G., Grandjean D., Sander D., Vuilleumier P. Electrophysiological correlates of rapid spatial orienting towards fearful faces // Cerebral Cortex. 2004. V. 14. № 6. P. 620.
Jolij J., Lamme V.A.F. Repression of unconscious information by conscious processing: evidence from affective blindsight induced by transcranial magnetic stimulation // PNAS. 2005. V. 102. № 30. P. 10 747.
Sigman M., Dehaene S. Parsing a cognitive task: a characterization of the mind’s bottleneck // PLoS Biol. 2005. V. 3. № 2. P. e37.
Banno H., Saiki J. The processing speed of scene categorization at multiple levels of description: The superordinate advantage revisited // Perception. 2015. V. 44. № 3. P. 269.
Holmes A., Winston J.S., Eimer M. The role of spatial frequency information for ERP components sensitive to faces and emotional facial expression // Cogn. Brain Res. 2005. V. 25. № 2. P. 508.
Eddy M.D., Holcomb P.J. The temporal dynamics of masked repetition picture priming effects: Manipulations of stimulus-onset asynchrony (SOA) and prime duration // Brain Res. 2010. V. 1340. P. 24.
Nakashima T., Kaneko K., Goto Y. et al. Early ERP components differentially extract facial features: Evidence for spatial frequency-and-contrast detectors // Neurosci. Res. 2008. V. 62. № 4. P. 225.
Pesciarelli F., Sarlo M., Leo I. The time course of implicit processing of facial features: An event-related potential study // Neuropsychol. 2011. V. 49. № 5. P. 1154.
Potts G.F. An ERP index of task relevance evaluation of visual stimuli // Brain Cogn. 2004. V. 56. № 1. P. 5.
Daffner K.R., Alperin B.R., Mott K.K. Age-related differences in early novelty processing: Using PCA to parse the overlapping anterior P2 and N2 components // Biol Psychol. 2015. V. 105. P. 83.
Wang H., Sun P., Ip C. et al. Configural and featural face processing are differently modulated by attentional resources at early stages: An event-related potential study with rapid serial visual presentation // Brain Res. 2015. V. 1602. P. 75.
Carretié L., Kessel D., García-Rubio M.J. et al. Magnocellular bias in exogenous attention to biologically salient stimuli as revealed by manipulating their luminosity and color // J. Cogn. Neurosci. 2017. V. 29. № 10. P. 1699.
Carretie L. Exogenous (automatic) attention to emotional stimuli: a review // Cogn. Affect. Behav. Neuroscience. 2014. V. 14. № 4. P. 1228.
Gibbons H., Bachmann O., Stahl J. The more you ignore me the closer I get: An ERP study of evaluative priming // Cogn. Affect. Behav. Neurosci. 2014. V. 14. № 4. P. 1467.
Jost K., Wendt M., Luna-Rodriguez A. et al. Strategic control over extent and timing of distractor-based response activation // J. Exp. Psychol. Learn. Mem. Cogn. V. 43. № 2. P. 326.
Mu T., Li S. The neural signature of spatial frequency-based information integration in scene perception // Exp. Brain Res. 2013. V. 227. № 3. P. 367.
Hickey C., Di Lollo V., McDonald J.J. Electrophysiological indices of target and distractor processing in visual search // J. Cogn. Neurosci. 2009. V. 21. № 4. P. 760.
Hilimire M.R., Hickey C., Corballis P.M. Target resolution in visual search involves the direct suppression of distractors: Evidence from electrophysiology // Psychophysiol. 2012. V. 49. № 4. P. 504.
Gaspar J.M., McDonald J.J. Suppression of salient objects prevents distraction in visual search // J. Neurosci. 2014. V. 34. № 16. P. 5658.
Neumann M.F., Viska C.G., van Huis S., Palermo R. Similar distraction, but differential suppression, for faces and non-face objects: Evidence from behaviour and event-related potentials // Biol. Psychol. 2018. V. 139. P. 39.
Del Zotto M., Pegna A.J. Processing of masked and unmasked emotional faces under different attentional conditions: an electrophysiological investigation // Front. Psychol. 2015. V. 6. P. 1691.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека