

Физиология человека, 2020, T. 46, № 4, стр. 30-43
Характеристика латентных периодов и параметров вариабельности составных элементов простой зрительно-моторной реакции (электромиографическое исследование)
В. И. Соболев *
Крымский федеральный университет
имени В.И. Вернадского
Симферополь, Россия
* E-mail: v.sobolev@mail.ru
Поступила в редакцию 12.06.2018
После доработки 25.12.2018
Принята к публикации 10.11.2019
Аннотация
В работе, с участием девушек 18–19 лет, установлено, что время обнаружения стимула (sDT – stimulus detection time) и длительность метрической моторной реакции (MIT – movement initiation time), как составных компонентов простой зрительно-моторной реакции с латентностью 173.1 ± 0.36 мс (sRT – simple reaction time), измеренные методом анализа интерференционной электромиограммы, составили 141.8 ± 0.32 и 31.3 ± 0.05 мс при относительном “вкладе” каждого из них в общее время простой зрительно-моторной реакции соответственно 81.9 и 18.1%. Средняя величина колебаний латентности sDT-показателя в ходе выполнения теста (цепной ряд динамики, абсолютный прирост на цепной основе) составила 31.9 ± 1.17 мс, а метрической моторной реакции соответственно 1.63 ± 0.05 мс, или в 19.6 раза меньше; в относительных величинах (цепной темп прироста) степень флуктуации sDT-показателя была равна 21.1 ± 0.78%, а метрической моторной 5.24 ± 0.17%, или в 4.0 раза меньше; эти результаты указывают на высокую стабильность метрического моторного компонента в структуре простой зрительно-моторной реакции и крайне высокую вариабельность параметра “время обнаружения стимула” (sDT). По данным регрессионного анализа, при удлинении простой зрительно-моторной реакции (от 120 до 350 мс) амплитуда колебаний (флуктуации) времени обнаружения стимула в цепном ряду динамики (параметр ΔsDT) нарастает в 27 раз быстрее, чем в случае метрической моторной реакции (ΔMIT), что свидетельствует о преобладающем вкладе sDT-компонента в общую вариабельность психомоторной реакции.
Изучение особенностей функционального состояния центральной нервной системы (ЦНС) с помощью методов психометрии широко используется в исследованиях психофизиологического характера, в частности в разных областях прикладной физиологии, неврологии, психиатрии, геронтологии, педагогике и психологии [1–5]. В большой степени это связано с высокой информативностью и базовыми принципами построения такого рода реакций, реализация которых требует достаточно высокого уровня нейронной организации с подключением фундаментальных блоков функциональных систем организма [6–8].
Простая зрительно-моторная реакция (ПЗМР) относится к категории простейших и с точки зрения своей структуры [5], характеризуется тремя параметрами: латентным периодом реакции (sRT – simple reaction time), временем обнаружения стимула (sDT – stimulus detection time) и временем инициации двигательного акта (MIT – movement initiation time).
Наиболее часто при оценке общей скорости сенсомоторных реакций используют параметр sRT, который численно определяется как время с момента предъявления стимула до момента начала метрического сокращения мышцы. В литературе приводятся значения латентного периода ПЗМР, располагающиеся в диапазоне от 160 до 417 мс, при дисперсии выборок, различающихся на порядок. Прежде всего, выявлено, что значение sRT зависит от длительности стимула и его интенсивности, которое можно описать степенной функцией: чем слабее стимул, тем больше время реакции, но после того, как стимул достигает определенной силы, время реакции становится постоянным [9]. Уровень возбуждения (arousal), независимо от его природы, укорачивает sRT, но только при средней степени выраженности [10]. Время sRT-реакции зависит также от пола, и у мужчин больше, чем у женщин [11], удлиняясь с возрастом [5, 12]. В обширных исследованиях D.L. Woods et al. [5] на примере репрезентативной выборки из 1469 испытуемых в тестах с предъявлением зрительного стимула на экране монитора пришли к выводу об удлинении sRT с возрастом от 217.9 мс у 20.8-летних до 239.1 в группе 59–65-летних; приведены также доказательства ускорения ЗМР при более коротких (1000 мс) межстимульных паузах по сравнению с более длинными (1800 мс). Выраженное влияние на латентность sRT оказывает выбор области сетчатки, на которую попадает стимул [13]. Приводятся данные, свидетельствующие о том, что процедура предъявления разномодальных стимулов во многом определяет время реакции [14], которое удлиняется при последовательном выполнении обеих задач (эффект распределения ресурсов внимания). Имеются свидетельства ускорения реакции у тревожных личностей [15], а также под воздействием негативных комментариев экспериментатора относительно ошибок испытуемого [16] или при угрозе и нанесении электрошокового импульса тока [17]. Выявлена положительная связь между скоростью реакции и интеллектом [18]. U. Lindenberger [3], основываясь на анализе обширной литературы по проблеме когнитивного старения (human cognitive aging), указывает, что более высокая оценка состояния некоторых когнитивных функций мозга человека сочетается с более быстрой и точной реакцией. Процесс голодания не изменяет скорость сенсомоторных реакций [19], а при развитии утомления [4, 20] и под воздействием отвлекающих внимание факторов, например фонового шума [21], она, наоборот, падает. Время sRT в большой степени зависит от предварительного оповещения о предстоящем стимуле (укорачивается) [22]; при этом большое значение приобретает как длительность стимула-оповещения, так и интервал времени между его предъявлением и предъявлением тестового стимула [23].
Таким образом, все эти и другие подобные работы, в которых измерялась длительность sRT, свидетельствуют о том, что время выполнения ЗМР определяется многочисленными факторами как внутренней, так и внешней среды, являясь высоковариабельным параметром. Следует отметить работу Y.A. Dordonova et al. [24], в которой авторы на основании сравнительного анализа литературных данных пришли к мнению, что значения латентных периодов ЗМР, полученных в разных исследованиях, несравнимы из-за отсутствия стандартизации.
Составной частью ЗМР является параметр времени обнаружения стимула (sDT – stimulus detection time), который соответствует временнóму периоду от момента предъявления испытуемому стимула до момента начала моторного акта. По данным литературы [5, 23, 25], значения sDT колеблются в пределах 180–300 мс, что в значительной степени осложняет проведение сравнительного анализа данных разных авторов. Обычно показатель sDT измеряется хронорефлексометром по моменту остановки секундомера тем или иным способом. Используются также и другие приемы, в частности, D.L. Woods et al. [5] длительность sDT рассчитывали как разницу между значением sRT и временем, требующимся для активации механической кнопки пальцем кисти, которое предварительно вычислялось в отдельных исследованиях с использованием теппинг-теста. По данным D.L. Woods et al. [5], время sDT в смешанной выборке 20-летних мужчин и женщин, рассчитанное подобным образом, составляет 125.8 мс и увеличивалось в группе 50-летних до 133.5 мс.
Более точные результаты в отношении латентности sDT могут быть получены в электромиографических (ЭМГ) исследованиях. В 1966 г. J. Botwinick et al. [23] на основании анализа ЭМГ-записей определили время sDT, составившее 109.3 мс (при сигме 14.27). Позже [25], латентный период ЭМГ-ответа простой сенсомоторной реакции, который тождественен параметру sDT, был определен в пределах 180–200 мс. В еще более позднем исследовании [26] время sDT при предъявлении светового стимула, по данным латентности ЭМГ-ответа, оценивалось в 190 ± 26 мс у молодых (22.5 года) и 236 ± 26 мс у пожилых участников эксперимента (68 лет).
Широкий диапазон колебаний латентности sDT обусловлен сложной природой его формирования. Наиболее перспективные направления в этой области связаны с построением математических и логистических моделей, описывающих основные принципы организации и самоорганизации нейронных сетей или отдельных структур ЦНС при обработке сенсорной информации и формировании решения [27–31], а также работы с проведением картирования мозга средствами ЭЭГ и др. методами [32, 33], определением латентности вызванных потенциалов [1, 32–34], моделированием работы нейронных сетей при опознавании стимула [30] и др. Время sDT определяется временными параметрами процесса формирования ощущения. В связи с этим особый интерес представляют работы, в которых разрабатываются теории формирования ощущения, восприятия и сенсорного осознания, построенных на так называемых принципах “кольца” А.М. Иваницкого [8], “динамического ядра” G.M. Edelman [35], а также на результатах исследований В.Я. Сергина [27] и др.
Второй компонент ЗМР – время инициации метрического сокращения мышцы (MIT – movement initiation time) является составной частью общей латентности двигательного акта. Основные механизмы организации двигательного акта в целом известны и изложены в фундаментальных работах Н.А. Бернштейна [7]. Следует отметить, что, строго говоря, запуск двигательного акта в мышце начинается с момента достижения уровня деполяризации постсинаптической мембраны нервно-мышечного синапса критического уровня после прихода спайка нервных импульсов в рамках выполнения моторной программы. В реальных условиях эксперимента приемлемо измерение времени моторной реакции, начиная с момента лавинообразной генерации в мышце потенциалов действия, что легко осуществимо при использовании метода интерференционной ЭМГ. По данным не столь многочисленных ЭМГ-исследований, время метрической моторной реакции оценивается в широком диапазоне значений: 38 мс [23], 64.6 мс [17] и больше [26]. Большинство подобных работ выполнены с использованием теперь уже устаревшего ЭМГ-оборудования [17, 23], характеризуются применением специфического алгоритма опыта, например, подачей сигнала оповещения о предстоящей стимуляции [23], побудительной электрической стимуляции испытуемого [17], часто без достаточно полного статистического анализа вариационных рядов, а также с помощью принципиально различающихся условий и методик регистрации параметров собственно моторного ответа. Таким образом, нельзя с определенностью сделать вывод относительно латентности MIT-реакции, значение которой при использовании разных методических приемов колеблется в диапазоне от нескольких десятков до нескольких сотен миллисекунд [5, 23].
Анализируя литературные данные можно отметить, что в большинстве работ показатели, характеризующие латентности параметров ЗМР (sRT, sDT и MIT), определялись преимущественно порознь, редко в парах “sRT-sDT”, что затрудняет вычисление соотношения между ними, а также выявление характера взаимосвязей и степени их выраженности. Названные аспекты послужили основой для формирования первой задачи данной работы.
Вторая задача данного исследования связана с вопросом оценки стабильности выполнения ЗМР, а также вариабельности ее составных компонентов. По данным литературы, дисперсия выборок для значений времени ПЗМР, приводимая разными авторами, может различаться на порядок: от 20164 до 729 [5], хотя обычно меньше. В подобном случае разброс дисперсий вызван различиями в методике и условиях проведения эксперимента. В работах психофизиологической направленности вариабельность сенсомоторных реакций рассматривается преимущественно с точки зрения статистической характеристики центральных тенденций вариационного ряда. С учетом характера нашей работы, представляет интерес та часть вариабельности, которая проявляется в ходе проведения теста (“от реакции до реакции”), т.е. в динамике выполнения испытуемым алгоритма опыта. Такой методический подход послужил основой для формулировки второй задачи нашей работы.
Что касается природы вариабельности сенсомоторных реакций, то безусловно, она связана с фундаментальными механизмами организации поведенческого акта [3, 6–8, 28, 36], реализующимися на уровне различных отделов ЦНС путем образования соответствующей функциональной системы организма [6].
Все перечисленные задачи послужили основой для формулировки цели данной статьи: на основании анализа результатов ЭМГ-исследования измерить абсолютные и относительные значения составных компонентов ПЗМР (sDT – время обнаружения стимула и MIT – время моторной метрической реакции), выявить характер зависимости между ними, а также провести сравнительную оценку основных параметров их вариабельности.
МЕТОДИКА
В исследовании участвовали 16 практически здоровых девушек (средний возраст 18.1 ± 0.1 года, медиана 18 лет), обучающихся в Крымском федеральном университете им. В.И. Вернадского, в осенне-зимний период 2017–2018 гг.
Процедура исследования и оборудование. В эксперименте, в ходе которого испытуемым предъявляли световой стимул, проводили непрерывную регистрацию ЭМГ-активности мышц предплечья, участвующих в сокращении пальцев кисти. По результатам анализа ЭМГ-записей (рис. 1) вычисляли время ПЗМР (sRT), а также периоды ее составных элементов: время обнаружения стимула (sDT) и время метрической моторной реакции (MIT).
Рис. 1.
К методике вычисления параметров ПЗМР. Вертикальные отметки на шкале времени указывают на момент предъявления светового стимула (первая отметка) и момент активации кнопки “СТОП” в начале фазы метрической моторной реакции (вторая отметка); 174 мс – полное время ПЗМР (sRT); 143 мс – время обнаружения стимула (sDT); 31 мс – время метрической моторной реакции (MIT).
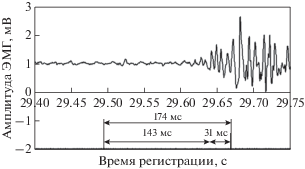
Метку “Начало реакции” на ЭМГ-записи наносили автоматически в момент предъявления испытуемому светового раздражителя, а метку “Конец реакции” – с помощью инициации механической кнопки “СТОП”, помещенной на выносной пульт “Samsung AA59-00741A”. Генерация метки на ЭМГ-записи (“Конец реакции”) осуществлялась в момент разрыва контактов кнопки при перемещении ее штока на расстояние не более 1 мм, т.е. практически в начале периода метрического сокращения мышц кисти. Во время эксперимента пульт с кнопкой фиксировали между большим (сверху) и четырьмя (снизу) пальцами правой руки. Благодаря хорошей эргономике пульта, в процессе выполнения моторной реакции при встречном движении пальцев (имитация хватательного рефлекса) существенно укорачивалось время срабатывания кнопки и, следовательно, сокращалось время генерации метки “Конец реакции” при инициации двигательного акта.
В качестве источника светового раздражителя использовали выносной точечный источник света на основе светодиода BL-L502UBC ярко-голубого свечения (Ultra Blue, 470 нм). Источник света располагали на расстоянии 40 см от кончика носа, а световой стимул предъявляли в оба глаза. Интенсивность светового раздражителя выбирали постоянной и предварительно аппаратными средствами устанавливали на приемлемом для глаза уровне 5000 мкд (IF = 20 мА) при длительности светового импульса 20 мс. Интервал между предъявляемыми стимулами колебался в диапазоне от 2 до 3 с и в автоматическом режиме задавался при помощи генератора случайных интервалов времени, построенного на основе функциональной комбинации четырех таймеров.
Для регистрации интерференционной ЭМГ использовали пластинчатые ЭМГ-электроды, биоусилитель, изготовленный на базе инструментального усилителя INA118, многоканальный цифровой USB-самописец (S-Recoder-L, Россия) и компьютер. Информацию записывали в виде alf-файлов при выбранной частоте семплирования (дискретизации) по каждому измерительному каналу 20 кГц (50 мкс). Электроды имели округлую форму диаметром 10 мм, изготовлялись из стандартных пластин электродов электрокардиографа и крепились на коже (4 см друг от друга) над мышцами внутренней поверхности предплечья, участвующих в сгибании кисти и ее пальцев, в частности в средней области поверхностного сгибателя пальцев (m. flexor digitorum superficialis).
Алгоритм опыта был следующим. После предварительного инструктажа о цели, задачах и ходе проведения эксперимента на внутреннюю поверхность правого предплечья накладывали ЭМГ-электроды, включали ЭМГ-запись и после серии из 15 тренировочных замеров скорости реакции без паузы осуществляли регистрацию основной серии тестов (40 измерений). Весь процесс осуществляли в автоматическом режиме.
В ходе анализа экспериментальных данных (alf-файлов) программными средствами АDClab на кривой записи ЭМГ (рис. 1) измеряли значение длительности двух показателей: время обнаружения стимула (sDT, мс) и время метрической моторной реакции (MIT, мс). В первом случае значение показателя соответствовало интервалу времени от момента предъявления светового стимула (первая отметка на шкале времени, рис. 1) до момента начала роста амплитуды ЭМГ относительно фона (начало процесса генерализованного возбуждения мышечных волокон). При измерении периода метрической моторной реакции программными методами измеряли интервал времени между точкой начала роста амплитуды ЭМГ (при достижении амплитуды ЭМГ выше фонового уровня на 3%) и моментом активации кнопки “СТОП” (вторая отметка). Сумма значений двух показателей (sDT и MIT) соответствовала полному времени ПЗМР (sRT). Следует отметить, что метрический компонент моторной реакции является составной частью общей двигательной реакции, продолжающейся и после операции “СТОП”, однако ее особенности в данной работе не изучали.
Статистический анализ. В ходе проведения анализа предварительно формировали 2 типа вариационных рядов. Первый из них (традиционный) представлял собой 16-ти вариантный ряд (по числу испытуемых) из значений индивидуальных средних арифметических величин исследуемых показателей, т.е. составленный по принципу: “один испытуемый – одна средняя арифметическая”.
Второй тип представлен сводными (объединенными) вариационными рядами, составленными путем объединения цепочкой 16-ти индивидуальных рядов (по числу 16-ти испытуемых). Поскольку такой способ применяется редко и требует соблюдения определенных правил, необходимо описание алгоритма подобной процедуры. На первом этапе для каждого из 16-ти испытуемых составляли по 3 индивидуальных ряда сочетанных значений sRT, sDT и MIT. Варианты в индивидуальных рядах располагали в хронологическом порядке, т.е. отражали динамику выполнения теста. На втором этапе в индивидуальных рядах каждого испытуемого проводили выбраковку сомнительных вариант (реакции фальстарта). Одновременно с удалением значения sRT удаляли и связанные (скрепленные) с ним значения sDT и MIT. В результате исходная длина индивидуальных рядов несколько укорачивалась, в среднем до 38 вариант (при сигме 0.44). На третьем этапе из индивидуальных рядов формировали сводные вариационные ряды путем их последовательного объединения. Поскольку в результате выбраковки сомнительных вариант длина индивидуальных рядов укорачивалась в среднем на 2 варианты, то конечная длина новообразованных объединенных рядов составляла 609 значений.
Таким способом были сформированы 3 сочетанных вариационных ряда для sRT, sDT и MIT-компонентов ПЗМР. Использованная процедура позволила сохранить в объединенных вариационных рядах весь спектр реакций экспериментальной группы испытуемых (в том числе и высоковариабельные значения), что улучшило репрезентативность выборок, а также позволило привлечь для их анализа более широкий набор статистических методов, в частности регрессионный и корреляционный анализы.
При статистической обработке данных использовали пакеты прикладных программ Excel и Statistica 7.0. После определения характера распределения данных в сводных вариационных рядах (тест Шапиро-Уилка) рассчитывали основные показатели описательной статистики. Статистическую значимость различий между двумя средними арифметическими величинами определяли с помощью двухвыборочного t-теста Стьюдента для выборок с различными дисперсиями при заданном уровне значимости p < 0.05. При оценке различий между двумя множествами использовали двухвыборочный F-тест для дисперсий. Для характеристики вариабельности значений центральной тенденции (средних арифметических) исследуемых показателей рассчитывали величины дисперсии выборок и коэффициенты вариации.
С целью проведения более широкого анализа степени стабильности выполнения ЗМР и оценки вариабельности ее основных компонентов дополнительно рассчитывали статистические параметры рядов динамики: абсолютный прирост на цепной основе (мс) и цепной темп прироста (%). Алгоритм расчета параметра “абсолютный прирост на цепной основе” (на примере sRT-показателя) был следующим. На первом этапе в индивидуальном хронологически выстроенном вариационном ряду (т.е. по ходу выполнения теста) вычисляли величину отклонения показателя sRT (мс) каждой последующей варианты от предшествующей (по модулю). В результате формировалось семейство из 16-ти новых индивидуальных вариационных рядов (по числу 16-ти испытуемых), представленных значениями sRT-отклонений (колебаний реакции) в динамике выполнения теста. На втором этапе путем последовательного объединения 16-ти отдельных индивидуальных вариационных рядов формировали сводный вариационный ряд sRT-отклонений. На третьем этапе для новообразованного сводного ряда рассчитывали основные параметры описательной статистики: среднюю арифметическую величину sRT-отклонений, т.е. “абсолютный прирост на цепной основе” со своей стандартной ошибкой, среднеквадратическим отклонением и дисперсией.
Эти же статистические параметры выражали и в относительных единицах (%, цепной темп прироста), что позволило нивелировать влияние чрезмерно различающихся средних арифметических величин исследуемых показателей.
Поскольку в наших исследованиях, благодаря небольшому числу замеров, эффект утомления еще не проявлялся, то измерение величины колебаний параметра времени “от реакции до реакции” в хронологическом ряду может оказаться полезным при оценке степени стабильности выполнения ЗМР. Метод анализа рядов динамики и прием объединения индивидуальных вариационных рядов позволил в дальнейшем провести более полную сравнительную количественную оценку статистической значимости различий степени стабильности (вариабельности) выполнения ЗМР и ее компонентов.
Во всех случаях сравнение анализируемых показателей и оценку статистической значимости различий проводили на основании проверки нулевой и альтернативной гипотез.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Анализ целостных вариационных рядов времени простой ЗМР и ее компонентов. На первом этапе описания результатов экспериментов были проанализированы 3 вариационных ряда, составленных традиционным способом, т.е. по принципу “один испытуемый – одна средняя арифметическая”. По данным табл. 1 видно, что значение sDT (148.7 ± 1.85) мс было существенно выше (в 4.8 раза), чем у MIT (31.12 ± 0.08) мс. Величины дисперсии, согласно двухвыборочному F-тесту для дисперсий, также различались, что выразилось в более высоком значении коэффициента вариации для показателя “время обнаружения стимула”. Характер распределения данных в 3-х описанных вариационных рядах соответствовал нормальному закону (тест Шапиро-Уилка). Однако дополнительно проведенные расчеты показали, что среди 48 индивидуальных выборок (16 для sRT-, 16 для SDT- и 16 для MIT-показателей) 31 нормальному закону не следовали. Такой результат закономерен, поскольку процедура усреднения индивидуальных данных, как правило, приводит к искусственному росту степени однородности ряда за счет нивелирования вариационного вклада высоковариабельных вариант.
Таблица 1.
Сравнительная характеристика значений различных компонентов ПЗМР
| Статистический показатель | Время ПЗМР (sRT), мс | Время обнаружения стимула (sDT), мс | Время метрической моторной реакции (MIT), мс | Отношение: sDT/MIT |
|---|---|---|---|---|
| Вариационный ряд, составленный из индивидуальных средних значений (n = 16) | ||||
| Среднее и ошибка средней | 178.6 ± 2.59 (100%) | 148.7 ± 1.85 (83.3) | 31.12 ± 0.08 (17.7%) | 4.8 ± 0.05 |
| Медиана | 181 | 150 | 31.2 | |
| Стандартное отклонение | 10.3 | 7.43 | 0.33 | |
| Дисперсия выборки | 107 | 55* | 0.11** | |
| Коэффициент вариации, % | 5.8 | 4.99 | 1.05 | |
| Соответствие нормальному закону распределения | Соответствует | Соответствует | Соответствует | |
| Количество испытуемых | 16 | 16 | 16 | |
| Объединенный (сводный) вариационный ряд (n = 609) | ||||
| Среднее и ошибка средней | 180.2 ± 1.34 (100%) | 149.0 ± 1.29 (82.7%) | 31.2 ± 0.06 (17.3%) | 4.8 ± 0.11 |
| Медиана | 172.8 | 141.5 | 30.9 | |
| Стандартное отклонение | 33.0 | 31.9 | 1.52 | |
| Дисперсия выборки | 1095 | 1017 | 2.3** | |
| Коэффициент вариации, % | 18.3 | 21.4 | 4.9 | |
| Соответствие нормальному закону распределения | Не соответствует | Не соответствует | Не соответствует | |
| Число измерений | 609 | 609 | 609 | |
Примечание: в круглых скобках приведен “вклад” (в %) соответствующего показателя в полное время ЗМР; * – различие между дисперсиями sDT и sRT статистически значимо (p < 0.05); ** – дисперсия MIT статистически значимо отличается (p < 0.05) от дисперсий sRT и sDT (двухвыборочный F-тест для дисперсий).
Таким образом, первоначальный анализ результатов экспериментов показал, что соотношение между sDT- и MIT-компонентами ПЗМР равно 4.8, а их вклад в полное время ЗМР составляет 83.3 и 17.7%; вариабельность sDT-показателя была несравненно выше, чем для MIT-реакции.
На следующем этапе были проанализированы объединенные ряды значений sRT-, sDT- и MIT-показателей, включающие весь спектр эмпирических вариант, в том числе и высоковариабельные. Прежде всего, выяснилось, что характер распределения данных в исходных эмпирических вариационных рядах для ПЗМР, а также для ее sDT- и MIT-компонентов ожидаемо не соответствовал нормальному закону (W-тест Шапиро-Уилка, табл. 1), что связано с присутствием в рядах значений запаздывающих реакций. Расчеты показали (на примере sDT), что всего 10 запаздывающих реакций (из 609) со временем более 250 мс (отличаются от средней на 38–81%) определяют 22% всей дисперсии выборки и формируют правостороннюю асимметрию, о чем может свидетельствовать сдвиг коэффициента асимметрии (Skewness) за счет указанных 10 вариант на 33%. Тем не менее, сохранение в объединенных рядах значений всего спектра эмпирических реакций улучшает репрезентативность выборок, адекватно отражая реальную вариабельность показателя времени выполнения ЗМР.
По данным табл. 1 видно, что средние арифметические для sDT и MIT были соответственно равны (149 ± 1.29) и (31.2 ± 0.06) мс, а значения медиан несколько отличались от средних арифметических (в пределах 1.0–5.0%). Относительный “вклад” показателя “время метрической моторной реакции” в полное время ПЗМР составил 17.3%, а показателя “время обнаружения стимула”, следовательно, 82.7%. Соотношение между периодами sDT и MIT было равным 4.8 ± 0.11. Значение величины дисперсий и коэффициентов вариации для вариационных рядов ПЗМР (sRT) и ее компонента sDT было приблизительно равным (соответственно 18.3 и 21.4%), а для периода MIT существенно отличалось в меньшую сторону (4.9%).
Сравнение результатов анализа двух типов вариационных рядов показывает, что в отношении принципиальных выводов о более высокой вариабельности sDT-компонента в сравнении с MIT-показателем, а также пропорций между ними противоречий не наблюдалось.
Анализ центрально-медианных секторов вариационных рядов времени ПЗМР и ее компонентов. Для выделения из вариационных рядов наименее вариабельной их части, которая предположительно может представлять собой множество наиболее статистически-вероятностных высокостабильных сенсомоторных ответов, генерируемых в центральных структурах зрительного анализатора, нами были использованы центрально-медианные сектора каждого из исследуемых рядов. Таким способом были сформированы 3 новых множества для sRT-, sDT- и MIT-показателей, состоящие из 127 вариант каждая (20% от общей длины ряда), и представляющие собой варианты 5–6 децилей. В такого рода усеченные выборки попали центрально-медианные сектора всех 16-ти индивидуальных вариационных рядов, что в значительной степени обеспечило репрезентативность исследуемых множеств. В этот же сектор попадают все варианты, определяющие в 16-ти индивидуальных рядах значение моды и средних арифметических.
Результаты анализа вновь сформированных вариационных рядов (табл. 2) позволили сделать следующие выводы. Во-первых, характер распределения данных во всех вариационных рядах, представляющих их центрально-медианные сектора (5–6 децили), с достаточной степенью статистической значимости (p < 0.05) стали описываться нормальным законом.
Таблица 2.
Сравнительная характеристика значений различных компонентов ПЗМР в центрально-медианной области вариационных рядов (5–6 децили вариационных рядов)
| Статистический показатель | Время ПЗМР (sRT), мс | Время обнаружения стимула (sDT), мс | Время метрической моторной реакции (MIT), мс | Отношение: sDT/MIT |
|---|---|---|---|---|
| Среднее и ошибка средней | 173.1 ± 0.36 (100%) | 141.8 ± 0.32 (81.9%) | 31.3 ± 0.05* (18.1%) | 4.53 ± 0.04 |
| Медиана | 173.2 | 141.7 | 31.25 | |
| Стандартное отклонение | 4.0 | 3.6 | 0.59 | |
| Дисперсия выборки | 16.1 | 13.1 | 0.35* | |
| Коэффициент вариации, % | 2.3 | 2.6 | 1.9 | |
| Эксцесс | –0.71 | –0.63 | –0.37 | |
| Асимметрия | 0.10 | 0.20 | 0.11 | |
| Соответствие нормальному закону распределения данных (тест Шапиро-Уилка) | Соответствует | Соответствует | Соответствует | |
| Число измерений | 127 | 127 | 127 | – |
Во-вторых, средние арифметические величины времени (мс) ЗМР sRT (173.1 ± 0.36), а также sDT-периода (141.8 ± 0.32) и метрической моторной реакции (31.3 ± 0.05) практически совпадали с их медианами, что свидетельствует о хорошей однородности вариационных рядов. Абсолютные значения средних арифметических для sRT и sDT были несколько ниже (3.9–4.1%), чем в случае полных вариационных рядов; для метрической моторной реакции такие различия не выявлены (табл. 1 и 2). Последнее можно рассматривать с точки зрения проявления высокой стабильности метрического моторного компонента ПЗМР (MIT), а также позволяет определить его абсолютное значение, равное (31.3 ± 0.05) мс.
В-третьих, соотношение между временем обнаружения стимула и метрическим моторным периодами составило 4.53 ± 0.04, а относительный вклад sDT и MIT в полное время ЗМР был соответственно равен (81.9 ± 0.12) и (18.1 ± 0.12)%.
В-четвертых, нормальный характер распределения эмпирических вариант, расположенных в центрально-медианных секторах (5 и 6 децили), позволяет при сравнительном анализе данных без всяких допущений применить методы параметрической статистики, в том числе и дисперсионный анализ. Следует особо подчеркнуть, что в случае анализа вариационных рядов центрально-медианного сектора значение дисперсий и соответственно стандартных отклонений и стандартных ошибок средних, а также коэффициентов вариации было существенно меньше, чем в случае с использованием целостных вариационных рядов (табл. 1 и 2), хотя общие тенденции и сохранились. Так, коэффициент вариации, по данным табл. 2, для sDT и MIT составил соответственно 2.5 и 1.9% при величинах дисперсии 13.1 и 0.35. Различия между величинами дисперсий, как показал двухвыборочный F-тест, были статистически значимы. Как видно, в центрально-медианном секторе исследуемых вариационных рядов, по данным расчета величин коэффициентов вариации и дисперсии, вариабельность значений времени обнаружения стимула была невысокой, а для метрической моторной реакции и вовсе незначительной.
Характеристика параметров вариабельности ПЗМР и ее компонентов методом анализа цепных рядов динамики. Результаты анализа динамических рядов (табл. 3) позволили сделать следующие выводы. Выявлено, что средняя (по модулю) величина отклонений (очередной варианты от предшествующей) для показателя “время обнаружения стимула” в хронологически выстроенном ряду составила (31.9 ± 1.17) мс, а для метрической моторной реакции соответственно (1.63 ± 0.05) мс, что было в 19.6 раза меньше (p < 0.001). Приведенный факт свидетельствует о более высокой стабильности метрической моторной реакции по сравнению со вторым компонентом ПЗМР – временем обнаружения стимула, sDT.
Таблица 3.
Сравнительная характеристика значений различных компонентов ПЗМР на основании анализа рядов динамики
| Статистический показатель | Метод расчета показателей цепных рядов динамики | Полное время ПЗМР (sRT) | Время обнаружения стимула (sDT) | Время метрической моторной реакции (MIT) |
|---|---|---|---|---|
| Среднее
арифметическое и его ошибка |
Абсолютный прирост на цепной основе, мс | 33.5 ± 1.21 100% |
31.9 ± 1.17 95.2% |
1.63 ± 0.05* 4.8% |
| Цепной темп прироста, % | 18.1 ± 0.66 | 21.1 ± 0.78 | 5.24 ± 0.17* | |
| Стандартное отклонение | Абсолютный прирост на цепной основе | 29.9 (897) | 28.9 (840) | 1.31 (1.73)* |
| Цепной темп прироста | 16.3 (267) | 19.4 (377) | 4.23 (17.9)* | |
| Число измерений | – | 608 | 608 | 608 |
Иллюстрацией к высказанному тезису являются результаты регрессионного анализа. Из рис. 2 и уравнений регрессии видно, что при удлинении ЗМР (sRT) в рамках исследованного диапазона 120–350 мс, значение времени обнаружения стимула (sDT) увеличивается несравненно более высокими темпами, чем время метрической моторной реакции – MIT:
Рис. 2.
Зависимость значений параметров sDT и MIT от общего времени выполнения ПЗМР. sDT – время обнаружения стимула, MIT – время метрической моторной реакции; sRT – полное время ПЗ-МР.
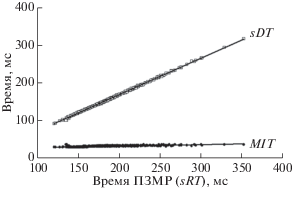
sRT = 0.96 [sTD] – 24.6 при bу/х = 0.96 ± 0.001 (p < 0.01) и коэффициенте корреляции Пирсона r = 0.99 ± 0.0012 (p < 0.01);
sRT = 0.036 [MIT] + 24.5 при bу/х = 0.036 ± 0.001 (p < 0.01) и r = 0.79 ± 0.025 (p < 0.01).
Приведенные значения коэффициентов регрессий в уравнениях прямых линий (bу/х) и их соотношение “sDT/MIT” (26.7 раз) свидетельствуют о высокой подвижности реакции “время обнаружения стимула”, как показателя длительности центральной задержки, и чрезвычайно стабильной метрической моторной реакции, как показателя двигательного акта, запускаемого моторной программой.
Еще одним свидетельством в пользу тезиса о высокой стабильности метрического моторного компонента и, наоборот, низкой – показателя времени обнаружения стимула, служат результаты анализа характера зависимости показателей рядов динамики от общей длительности ПЗМР (табл. 3, рис. 3 и 4).
Рис. 3.
Зависимость колебаний времени обнаружения стимула и метрической моторной реакции от общего времени выполнения ПЗМР. Абсолютный прирост на цепной основе определялся в динамике выполнения теста путем вычисления величины отклонения времени каждой последующей реакции от предыдущей. Остальные обозначения см. рис. 2.
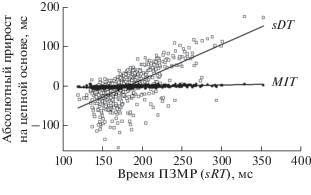
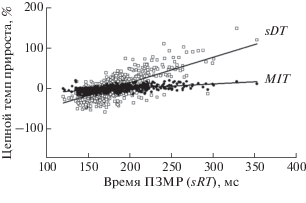
Рис. 4.
Зависимость колебаний времени обнаружения стимула и метрической моторной реакции, выраженных в относительных единицах (%), от общего времени выполнения ПЗМР. Цепной темп прироста в относительных единицах (%) определялся в динамике выполнения теста путем вычисления величины отклонения времени каждой последующей реакции от предыдущей. Остальные обозначения см. рис. 2.
Расчеты показали, что при удлинении простой зрительно-моторной реакции амплитуда колебаний времени обнаружения стимула (параметр ΔsDT – абсолютный прирост на цепной основе) нарастает в 27 раз быстрее (p < 0.001), чем в случае метрической моторной реакции. Такой вывод следует из сравнения соответствующих коэффициентов регрессии. Уравнения регрессии имеют следующий вид:
sRT = 0.89 [ΔsDT] – 160 при bу/х = 0.89 ± 0.039 (p < 0.01) и r = 0.67 ± 0.03 (p < 0.01), где [ΔsDT] – величина абсолютного прироста (колебаний) времени обнаружения стимула, рассчитанная на цепной основе, мс; bу/х – коэффициент регрессии; sRT – время простой зрительно-моторной реакции, мс;
sRT = 0.033 [ΔMIT] – 5.99 при bу/х = 0.033 ± ± 0.0022 (p < 0.01) и r = 0.52 ± 0.035 (p < 0.01), где [ΔMIT] – величина абсолютного прироста (колебаний) времени метрической моторной реакции, рассчитанная на цепной основе, мс.
Важно отметить, что поскольку средние арифметические величины абсолютных отклонений для sDT- и MIT-показателей чрезмерно различались (31.9 и 1.63 мс, табл. 3), для более корректной сравнительной оценки степени вариабельности двух названных параметров должны быть использованы величины, рассчитанные в относительных единицах (показатель “цепного темпа прироста”, %). Указанная процедура предназначена для нивелирования ошибки при интерпретации результатов, вызванной существенно различающимися средними арифметическими величинами. Так, по данным табл. 3 видно, что средняя величина параметра “цепной темп прироста, %” в случае показателя “время метрической моторной реакции” (5.24 ± 0.17)% оставалась статистически значимо ниже (в 4.0 раза, p < 0.001), чем для периода “время обнаружения стимула” (21.1 ± 0.78)%.
Рис. 4 иллюстрирует динамику зависимости показателей ΔsDTотн и ΔsDTотн от времени ПЗМР (sRT), а уравнения регрессии имели следующий вид:
sRT = 0.63 [ΔsDTотн] – 109 при bу/х = 0.63 ± 0.024 (p < 0.01) и r = 0.72 ± 0.028 (p < 0.01), где [ΔsDTотн] – величина относительного темпа прироста времени реакции обнаружения стимула, рассчитанная на цепной основе, %.
sSRT = 0.107 [ΔMITотн] – 19.1 при bу/х = 0.107 ± ± 0.007 (p < 0.01) и r = 0.53 ± 0.034 (p < 0.01), где [ΔMITотн] – величина относительного темпа прироста времени метрической моторной реакции, рассчитанная на цепной основе, %.
Можно видеть, что вместе с удлинением времени выполнения ЗМР размах колебаний значений sDT в соответствующем ряду динамики нарастал (bу/х = 0.63 ± 0.024) в 5.9 раза быстрее, чем метрической моторной реакции MIT (bу/х = 0.107 ± ± 0.007).
Обсуждая полученные результаты, прежде всего, следует отметить, что метод ЭМГ-исследования, использованный в данной работе, позволил вычислить абсолютное значение времени метрической моторной реакции, которое составило 31.3 ± 0.05 мс (18.1% от длительности всей ПЗМР). Данный параметр психомоторной реакции оказался высокостабильным и существенно не зависел от общей длительности ЗМР и методики статистического анализа. Такой факт свидетельствует о том, что после запуска программы моторного ответа из соответствующих структур ЦНС весь процесс ее реализации в эффекторном звене при условии нормы и максимальной мотивации испытуемого осуществляется с высокой степенью стабильности, что и предопределяет формирование стандартного двигательного акта. В иерархической структуре уровня организации движения, как считал H.А. Бернштейн [7], исследуемая сенсомоторная реакция относится к макродвижениям, т.е. одному из наиболее простых типов, которые обеспечиваются пирамидно-стриальным уровнем организации. Относительно простой уровень организации макродвижения в значительной мере упрощает его реализацию и в случае неизменных условий должно положительно отражаться на стабильности выполнения такого двигательного акта, что и было экспериментально подтверждено.
Метод ЭМГ-регистрации ЗМР позволил вычленить из нее и второй компонент – sDT, значение которого (при использовании целостного вариационного ряда и центрально-медианного сектора в области 6–7 децилей, табл. 1 и 2) составило соответственно (149.0 ± 1.29) и (141.8 ± 0.32) мс, что в 4–5 раз превышало значение MIT. Следовательно, относительный вклад sDT- и MIT-компонентов в общее время ПЗМР (sRT), независимо от метода расчета, составил 83 к 17% в первом случае, и 82 к 18%, во втором, т.е. практически не различался.
Сравнительный анализ значений sRT, определенных в наших исследованиях, с результатами других авторов показывает, что в одних случаях они совпадают [1, 36], а в других – нет [5]. С нашей точки зрения это может быть связано, помимо всего прочего, с конструктивными особенностями устройства для остановки отсчета времени sRT-реакции, что окажет влияние на общее время ЗМР. Более надежным и информативным показателем является параметр “время обнаружения стимула (sDT)”. Значения sDT, определенные в наших исследованиях в возрастной группе 18-летних девушек, близки к данным, приведенным для 20.8-летних испытуемых D.L. Woods et al. [5]. Однако необходимо учесть, что в отличие от нашей модели (диффузная вспышка света) в исследованиях D.L. Woods et al. в качестве светового стимула использовалась высококонтрастная мишень (high-contrast bulls eyes), а значение sDT рассчитывалось на основании косвенных данных. Прямые измерения sDT в силу методической сложности при массовых обследованиях, к сожалению, проводятся редко, но в тех случаях, когда они были выполнены [23, 26], абсолютные значение sDT были близки к нашим.
Различия в методике могут отражаться на абсолютных величинах показателей простой ЗМР, что не исключает сохранение общих закономерностей при ее реализации. Однако параметры вариабельности сенсомоторных реакций в силу ее (вариабельности) статистической природы изменяются в меньшей степени. Сравнительный анализ величин стандартных отклонений и коэффициентов вариации для sRT и sDT, приведенных в ряде публикаций [1, 5, 12, 37, 38], показал их хорошее соответствие с нашими результатами, вплоть до полного совпадения, например для коэффициентов вариации (18 и 18.3%).
Использованный в данной работе метод анализа цепных рядов динамики позволил вычислить в абсолютных (мс) и относительных (%) величинах степень стабильности выполнения ПЗМР и соответственно ее компонентов sDT и MIT-параметров. В этой связи можно сделать следующее заключение (табл. 3): величина колебаний времени sDT-реакции по ходу выполнения теста в абсолютных (мс) и относительных (%) единицах измерения была соответственно в 19.6 и 4 раза выше, чем MIT-реакции; основная доля вариабельности ПЗМР связана с высокодинамичным sDT-компонентом (95.2%), т.е. временем обнаружения стимула.
Несмотря на обширные исследования природы вариабельности психомоторных реакций [2, 12, 37] механизмы ее формирования во многом не ясны. Однако несомненно, что она является фундаментальным свойством любой саморегулирующейся системы (и не только саморегулирующейся) и обусловлена вариабельностью функционирования ее механизмов на разных уровнях организации ЦНС [39]. В последнее время интенсивно разрабатываются теории так называемого нейронного шума, как непременного условия моторного обучения, коррекции движения по feedback принципу и сохранения в рабочем состоянии нейронной сети, обеспечивающей функциональную готовность к выполнению освоенного ранее моторного акта [40]. Этот процесс отражает статистически-вероятностный характер функционирования нейронных сетей и, как можно полагать, вносит свою долю в общую вариабельность сенсорной реакции.
Хорошо обоснованными моделями нейронной организации сенсорного ответа являются так называемые модели накопления (accumulator models) [31], а также ее разновидности. D.P. Hanes et al. [41] пришли к выводу, что произвольное движение (саккады) начинается только тогда, когда нейронная активность в моторной области коры (Macaca mulatta) достигает уровня порога (принцип триггера), а время реакции, что важно, определяется вариабельностью стохастического процесса скорости достижения нейронной активности уровня такого порога. Согласно предложенной авторами модели, при низком пороге срабатывания нейронного триггера время реакции будет короче, чем при более высоком уровне. Модель характеризуется асимметричным распределением времени реакции (the skewed reaction-time distribution), что подтверждается результатами реального эксперимента, в том числе и нашего. В этой связи можно сослаться на представления Е.Н. Соколова и Н.И. Незлиной [42] о роли так называемых командных нейронов, участвующих в процессах организации функциональной связи между сенсорными и моторными компонентами при реализации условного рефлекса. Источником вариабельности в такой нейрональной системе служит процесс ритмического колебания уровня срабатывания нейрона-пейсмекера.
Наконец, колебание времени sDT-компонента может быть связано со следовыми эффектами в нейронных цепях, запустившимися предшествующей реакцией. В этом случае должна наблюдаться четкая зависимость между скоростью реакции, с одной стороны, и периодом межстимульных интервалов, а также стимулами предупредительного оповещения, с другой. Такая зависимость, как известно, давно установлена [23], и может служить основой для еще одного механизма вариабельности, поскольку продолжительность периода активности следового эффекта является величиной не постоянной.
Высокая вариабельность показателя sDT – времени обнаружения стимула может быть связана также с процессами формирования ощущения, в частности рассматриваться в свете теории кольцевого движения возбуждения (его повторного входа) как основы информационного синтеза [8]. Наряду с длинными кольцами циркуляции возбуждения с латентностью 100–200 мс, могут функционировать, по-видимому, и более короткие кольца с меньшей латентностью, а также механизмы, которые связывают с реверберацией возбуждения с центром в таламусе (гипотеза F. Crick) [43]. Известны также гипотезы “динамического ядра” G.M. Edelman [35] и многочисленные логистические теории, например, модель перцептивного связывания сенсорных событий и модель авто-отождествления В.Я. Сергина [27], которая оценивает латентность цикла осознания стимула около 50 мс.
Важным является общее свойство таких моделей – сенсорный ответ в той или иной форме формируется на основе статистически-вероятностного принципа обработки сенсорной информации и, следовательно, по самой природе должен быть вариабельным.
Однако, несмотря на то, что в основе вариабельности сенсомоторных реакций лежат нейрофизиологические механизмы, это не препятствует при объяснении ее природы принимать во внимание участие так называемых субъективных факторов, которые реализуются уже на другом уровне организации сенсорной системы. В работах психофизиологического характера [44] скорость сенсомоторных реакций рассматривается как один из показателей степени активности ЦНС и используется в качестве теста такой активности.
На уровне психофизиологического эксперимента устойчивость и способность к концентрации внимания, уровень мотивации испытуемого и другие психические проявления уже исходно сами по себе не являются стабильными, что приводит к закономерному росту вариабельности “реакции обнаружения стимула”, о чем свидетельствует отмеченный нами существенный размах колебаний sDT (от 100 до 320 мс).
Высокий уровень вариабельности sDT-показателя позволил предположить, что в структуре времени выполнения ЗМР может присутствовать достаточно стабильная “нейрофизиологически детерминированная составляющая”. Можно было предположить, что такого рода множество, отражающее статистически-вероятностное значение “установочной точки” в функциональной системе организации сенсомоторной реакции, можно обнаружить в центрально-медианном секторе исходного вариационного ряда. Проведенный статистический анализ позволил вычислить предполагаемое среднее нейрофизиологически детерминированное время параметра sDT, которое составило (141.8 ± 0.32) мс. Данный показатель центрально-медианной выборки характеризуется чрезвычайно низкой вариабельностью, различающейся по величине дисперсии относительно с sDT-реакцией целостного вариационного ряда в 77 раз (табл. 1 и 2). Этот факт предположительно можно рассматривать с точки зрения существования особого механизма, обеспечивающего генерацию и воспроизводство серии высокостабильных сенсорных ответов (в нашем случае 20% всех реакций). Если из времени предполагаемой “нейрофизиологически детерминированной реакции” (141.8 ± 0.32) мс вычленить период 20–25 мс (время достижения информации о стимуле в первичную зрительную кору) и время достижения спайка нервных импульсов из моторной коры к мышцам кисти (10 мс), то оставшееся время 110–120 мс хорошо согласуется со значением скорости формирования ощущения в гипотезе “повторного входа возбуждения как основе информационного синтеза” А.М. Иваницкого [8].
Значение “нейрофизиологически детерминированной составляющей”, отражающее наиболее вероятную скорость сенсомоторной реакции, при выполнении цикла ПЗМР может совершать спонтанные флуктуации вокруг значения своего “установочного” уровня (“set point”), отражая статистически-вероятностный характер процесса формирования сенсорного ответа. Такая “set point” во многом является аналогом нейронного триггера с меняющимся порогом срабатывания [41]. Значение “set point” не является величиной раз и навсегда установленной, но может сдвигаться на новый уровень в соответствии с изменившимися условиями внутренней и внешней среды, в частности и под влиянием субъективных факторов. Такое предположение согласуется с фундаментальными принципами организации функциональной системы [6], в которой “полезный результат” и является предметом регуляции, а его плавающий характер обусловлен текущими задачами и вероятностно-статистическими законами формирования сенсорного ответа в нейронных сетях соответствующих центральных структур зрительного анализатора.
ВЫВОДЫ
1. На основе метода интерференционной ЭМГ установлено, что время обнаружения стимула (sDT) и длительность метрической моторной реакции (MIT), как составных компонентов ПЗМР с латентностью 173.1 ± 0.36 мс (sRT), составили 141.8 ± 0.32 и 31.3 ± 0.05 мс при относительном “вкладе” каждого из них в общее время сенсомоторной реакции соответственно 81.9 и 18.1%.
2. Соотношение между значением sDT-показателя (“время обнаружения стимула”) и длительностью метрической моторной реакции (MIT) в случае анализа целостных вариационных рядов или их центрально-медианных секторов не различалось и колебалось в пределах 4.5–4.8.
3. Средняя величина колебаний латентности sDT-показателя в ходе выполнения теста (цепной ряд динамики, абсолютный прирост на цепной основе) составила 31.9 ± 1.17 мс, а метрической моторной реакции соответственно 1.63 ± 0.05 мс, или в 19.6 раза меньше; в относительных величинах (цепной темп прироста) степень флуктуации sDT-показателя была равна 21.1 ± 0.78%, а метрической моторной 5.24 ± 0.17%, или в 4.0 раза меньше; эти результаты указывают на высокую стабильность метрического моторного компонента в структуре ПЗМР и крайне высокую вариабельность параметра “время обнаружения стимула” (sDT).
4. По данным регрессионного анализа, при удлинении времени выполнения ПЗМР в диапазоне 120–350 мс темп нарастания длительности sDT-показателя и амплитуды его флуктуаций в абсолютных единицах измерения (мс) был в 26–27 раз выше, чем метрической моторной реакции, что свидетельствует о преобладающем вкладе sDT-компонента в общую вариабельность психомоторной реакции.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены локальным биоэтическим комитетом Гуманитарно-педагогической академии (филиал) Крымского федерального университета им. В.И. Вернадского (Ялта).
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Благодарности. Автор выражает благодарность заведующему кафедрой здоровья и реабилитации Института педагогики, психологии и инклюзивного образования Гуманитарно-педагогической академии (филиал) Крымского федерального университета им. В.И. Вернадского М.Н. Попову за помощь, оказанную при организации проведения данного исследования.
Конфликт интересов. Автор декларирует отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Айдаркин Е.К. Исследование особенностей взаимодействия зрительной и слуховой систем в условиях сенсомоторной интеграции // Валеология. 2006. № 3. С. 82.
Kochan N.A, Bunce D., Pont S. et al. Is intraindividual reaction time variability an independent cognitive predictor of mortality in old age? Findings from the Sydney Memory and Ageing Study // PLoS One. 2017. V. 12. № 8: e0181719.
Lindenberger U. Human cognitive ageing: Corriger la fortune? // Science. 2014. V. 346. № 6209. P. 572.
Özdemir R.A., Kirazcı S., Ugras A. Simple reaction time and decision making performance after different physical workloads: an examination with elite athletes // Int. J. Human Sciences. 2010. V. 7. № 2. P. 655.
Woods D.L., Wyma J.M., Yund E.W. et al. Factors influencing the latency of simple reaction time // Front. Hum. Neurosci. 2015. V. 9. № 131. P. 131.
Анохин П.К. Философские аспекты теории функциональной системы: избр. труды. М.: Наука, 1978. 399 с.
Бернштейн Н.А. Физиология движений и активность. М.: Наука (Серия “Классики науки”), 1990. 497 с.
Иваницкий А.М. Мозговая основа субъективных переживаний: гипотеза информационного синтеза // Журн. высш. нерв. деят. 1996. Т. 46. № 2. С. 241.
Luce R.D. Response Times: Their Role in Inferring Elementary Mental Organization. N.Y.: Oxford University Press, 1986. 576 p.
Masanobu A., Choshi K. Contingent muscular tension during a choice reaction task // Percept. Mot. Skills. 2006. V. 102. № 3. P. 736.
Der G., Deary I.J. Age and sex differences in reaction time in adulthood: Results from the United Kingdom health and lifestyle survey // Psychol. Aging. 2006. V. 21. № 1. P. 62.
Silverman I.W. Simple reaction time: It is not what it used to be // Am. J. Psychol. 2010. V. 123. № 1. P. 39.
Ando S., Kida N., Oda S. Practice effects on reaction time for peripheral and central visual fields // Percept. Mot. Skills. 2002. V. 95. № 3. P. 747.
Hsieh S. Tasking shifting in dual-task settings // Percept. Mot. Skills. 2002. V. 94. № 2. P. 407.
Robinson M.C., Tamir M. Neuroticism as mental noise: a relation between neuroticism and reaction time standard deviations // J. Pers. Soc. Psychol. 2005. V. 89. № 1. P. 107.
Panayiotou G., Vrana S.R. The role of self-focus, task difficulty, task self-relevance, and evaluation anxiety in reaction time performance // Motiv. Emotion. 2004. V. 28. № 2. P. 171.
Weiss A.D. The locus of reaction time change with set, motivation and age // J. Gerontol. 1965. V. 20. № 1. P. 60.
Deary I.J., Der G., Ford G. Reaction times and intelligence differences: A population-based cohort study // Intelligence. 2001. V. 29. № 5. P. 389.
Cheatham R.A., Roberts S.B., Das S.K. et al. Long-term effects of provided low- and high-glycemic-load low-energy diets on mood and cognition // Physiol. Behav. 2009. V. 98. № 3. P. 374.
Cote K.A., Milner C.E., Smith B.A. et al. CNS arousal and neurobehavioral performance in a short-term sleep restriction paradigm // J. Sleep Res. 2009. V. 18. № 3. P. 291.
Trimmel M., Poelzl G. Impact of background noise on reaction time and brain DC potential changes of VDT-based spatial attention // Ergonomics. 2006. V. 49. № 2. P. 202.
Brebner J.T., Welford A.T. Introduction: an historical background sketch / Ed. Welford A.T. N.Y.: Reaction Times. Academic Press, 1980. P. 1.
Botwinick J., Thompson L. Premotor and motor components of reaction time // J. Exp. Psychol. 1966. V. 71. № 1. P. 9.
Dordonova Y.A., Dordonov Y.S. Is there any evidence of historical slowing of reaction time? No, unless we compare apples and oranges // Intelligence. 2013. V. 41. № 5. P. 674.
Stull A.G., Kearney J.T. Effects of variable fatigue levels on reaction-time components // J. Mot. Behav. 1978. V. 10. № 3. P. 223.
Hinder M.R., Fujiyama H., Summers J.J. Premotor-Motor Interhemispheric Inhibition Is Released during Movement Initiation in Older but Not Young Adults // PLoS One. 2012. V. 7. № 12. e52573.
Сергин В.Я. Авто-отождествление паттернов нейронной активности как физиологический механизм осознания // Журн. высш. нерв. деят. 2016. Т. 66. № 3. С. 259.
Zandbelt B., Purcell B.A., Palmeri T.J. et al. Response times from ensembles of accumulators // Proc. Natl. Acad. Sci. 2014. V. 111. № 7. P. 2848.
Ratcliff R., Tuerlinckx F. Estimating parameters of the diffusion model: Approaches to dealing with contaminant reaction times and parameter variability // Psychon. Bull Rev. 2002. V. 9. № 3. P. 438.
Engel T.A., Chaisangmongkon W., Freedman D.J., Wang X.-J. Choice-correlated activity fluctuations underlie learning of neuronal category representation // Nat. Commun. 2015. V. 6. P. 6454.
Heathcote A., Love Jonathon. Linear deterministic accumulator models of simple choice // Front. Psychol. 2012. V. 3. P. 292.
Strelets V.B. EEG mapping in emotional and cognitive pathology / Conceptual Advances in Russian Neurosciences: Complex Brain Functions // Eds. Miller R., Ivanitsky A.M., Balaban P.M. UK: Harwood Academic Publishers. Reading, 1999. P. 151.
Иваницкий А.М., Ильюченок И.Р. Картирование биопотенциалов мозга при решении вербальной задачи // Журн. высш. нерв. деят. 1992. Т. 42. № 4. С. 627.
Николаев А.Р., Иваницкий Г.А., Иваницкий А.М. Исследование корковых взаимодействий в коротких интервалах времени при поиске вербальных ассоциаций // Журн. высш. нерв. деят. 2000. Т. 50. № 1. С. 44.
Edelman G.M., Tononi G.A. Universe of Consciousness. N.Y.: Basic Books, 2000. 274 p.
Welford A.T. Choice reaction time: Basic concepts / Ed. Welford A.T. N.Y.: Reaction Times. Academic Press, 1980. P. 73.
Dykiert D., Der G., Starr J.M., Deary I.J. Age differences in intra-individual variability in simple and choice reaction time: systematic review and meta-analysis // PLoS One. 2012. V. 7. № 10. e45759.
Hultsch D.F., MacDonald S.W.S., Dixon R.A. Variability in Reaction Time Performance of Younger and Older Adults // J. Gerontol: Series B. 2002. V. 57. № 2. P. 101.
MacDonald C.J, Meck W.H. Systems-level integration of interval timing and reaction time // Neurosci. Biobehav. Rev. 2004. V. 28. № 7. P. 747.
Rokni U., Richardson A.G., Bizzi E. et al. Motor Learning with Unstable Neural Representations // Neuron. 2007. V. 54. № 4. P. 653.
Hanes D.P., Schall J.D. Neural Control of Voluntary Movement Initiation // Science. 1966. V. 274. № 5286. P. 427.
Sokolov E.N., Nezlina N.I. [Conditioned reflex: detector and command neuron] // Zh. Vyssh. Nerv. Deiat. Im. I.P. Pavlova. 2007. V. 57. № 1. P. 5.
Crick F. The Astonishing Hypothesis: The Scientific Search for the Soul. N.Y.: Charles Scribner’s Sons, 1994. 317 p.
Ильин Е.П. Психомоторная организация человека. СПб.: Питер, 2003. 384 с.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека