

Физиология человека, 2021, T. 47, № 2, стр. 63-71
Феномен коактивации мышц при модуляции ходьбы с помощью электрической стимуляции спинного мозга
Т. Р. Мошонкина 1, *, Н. Д. Шандыбина 1, С. А. Моисеев 2, А. А. Гришин 1, Ю. П. Герасименко 1
1 ФГБУН Институт физиологии имени И.П. Павлова РАН
Санкт-Петербург, Россия
2 ФГБОУ ВО Великолукская государственная академия физической культуры и спорта
Великие Луки, Россия
* E-mail: moshonkina@infran.ru
Поступила в редакцию 27.05.2020
После доработки 07.06.2020
Принята к публикации 30.09.2020
Аннотация
Впервые проведено исследование коактивации мышц-антагонистов на фоне неинвазивной электрической стимуляции спинного мозга. Во время ходьбы здоровых добровольцев по беговой дорожке проводили чрескожную электрическую стимуляция (ЧЭС) дорзальных корешков спинного мозга, чтобы во время фазы переноса адресно активировать моторные пулы мышц-сгибателей, а во время фазы опоры – мышц-разгибателей. Такая стимуляция меняла кинематику походки. Значения коэффициента коактивации (КК) мышц в фазах опоры и переноса при ходьбе со стимуляцией сравнивали с этими значениями при ходьбе без стимуляции. Было получено, что ЧЭС увеличивает значения КК мышц бедра на ~10% и голени на ~5%. Стимуляция также меняет соотношение между величинами КК мышц бедра и голени в фазах шага. При стимуляции в фазе опоры зарегистрировано достоверное отличие между КК мышц бедра и КК мышц голени в фазе опоры, отсутствовавшее при ходьбе без стимуляции. При стимуляции в фазе переноса не было отличия между КК мышц голени в фазе переноса и КК мышц голени фазе опоры, которое регистрировали при ходьбе без стимуляции, т.е. при такой стимуляции голеностопный сустав был зафиксирован в фазе переноса в недоразогнутом положении, в том же, в каком он находился в фазе опоры. При ходьбе, сопровождавшейся чередующейся стимуляцией: моторных пулов мышц-сгибателей в фазе переноса и моторных пулов мышц-разгибателей в фазе опоры, – жесткость голеностопного сустава в фазе переноса была такой же, как и в фазе опоры. Следовательно, величина и направленность изменений КК зависят от пространственно-временны́х характеристик стимуляции и коррелирует с изменениями походки, которые вызывает адресная стимуляция дорзальных корешков спинного мозга.
Коактивация мышц – это синхронное сокращение мышц-антагонистов, осуществляющих тягу в одном и том же суставе. Как правило, коэффициент коактивации (КК) рассчитывают за один шагательный цикл. Понятие коактивации обсуждают в контексте биомеханики движений и участия разных отделов мозга в управлении движением. Увеличение коактивации связывают с увеличением жесткости сустава; принято считать, что управление коактивацией происходит на супраспинальном уровне [1]. Неоднократно продемонстрировано, что у пациентов с двигательными нарушениями после реабилитационных мероприятий наблюдают уменьшение значений КК мышц, обеспечивающих движения, на которые были направлены реабилитационные процедуры [1]. Так у детей с ДЦП после курса чрескожной электрической стимуляции (ЧЭС) спинного мозга на фоне локомоторной механотерапии зарегистрированы как достоверное уменьшение КК мышц бедра и голени, так и достоверное увеличение объема движений в суставах ног [2].
В процессе разработки “нейропротеза” – устройства и технологии для облегчения ходьбы пациентов, перенесших инсульт, – проведены исследования с участием здоровых добровольцев в качестве испытуемых. В процессе ходьбы ЧЭС корешков спинного мозга обеспечивала пространственно-временнýю адресную активацию моторных пулов: мышц-сгибателей – во время фазы переноса, мышц-разгибателей – во время фазы опоры [3]. При ходьбе по беговой дорожке ЧЭС сгибателей достоверно увеличивала подъем стопы над опорой, а ЧЭС разгибателей уменьшала длительность фазы опоры, увеличивала частоту шагов.
Задача настоящего исследования – определить влияние пространственно-временнóй адресной активации моторных пулов методом ЧЭС во время равномерной ходьбы на коактивацию мышц-антагонистов при адресном воздействии на эти мышцы в фазах опоры и переноса.
МЕТОДИКА
В исследовании участвовали здоровые добровольцы (n = 6). Все испытуемые мужского пола (средний возраст ~25 лет). Исследование выполняли на базе НИИ Проблем спорта и оздоровительной физической культуры ФГБОУ ВО “Великолукская государственная академия физической культуры и спорта” (ВЛГАФК). В процессе исследования испытуемые шли по беговой дорожке (h/p/cosmos gaitway®) со скоростью ~1.5 км/ч. После 15 с ходьбы начинали ЧЭС правого дорзального корешка спинного мозга, связанного с моторным пулом мышц-разгибателей (активация разгибателей) во время фазы опоры правой ноги, через 15 с эту стимуляцию прекращали и начинали ЧЭС правого дорзального корешка спинного мозга, связанного с моторным пулом мышц-сгибателей (активация сгибателей), во время фазы переноса правой ноги, через 15 с начинали чередующуюся ЧЭС разгибателей и сгибателей во время фаз опоры и переноса правой ноги соответственно (рис. 1). Исследование повторяли в той же последовательности, стимулируя корешки слева в зависимости от фаз шага левой ноги.
Рис. 1.
Принцип пространственно-временнóй чрескожной стимуляции спинного мозга в нейропротезе. А – схема расположения катодов и последовательность ЧЭС во времени; белые кружки – неактивные катоды; серые кружки – катоды, активные во время фазы переноса (Th11–12) и опоры (L1–2) ипсилатеральной ноги; пунктирная линия – осевая линия позвоночника; Th11–12 и L1–2 – позвонки. Б – алгоритм ЧЭС на примере последних 15 с исследования и принцип анализа ЭМГ на примере m. tibialis anterior (ТА); ЧЭС в зависимости от фаз шага, определенной по перемещению первого пальца стопы (кривая под электродами); ЭМГ инвертирована и нормированы к 100% (точечная кривая ТА, а), затем построена огибающая (б); ЭМГ синхронизирована с данными кинематики (в – фаза опоры, г – фаза переноса); испытуемый Д.Г., ходьба со скоростью 1.5 км/ч, правая нога.
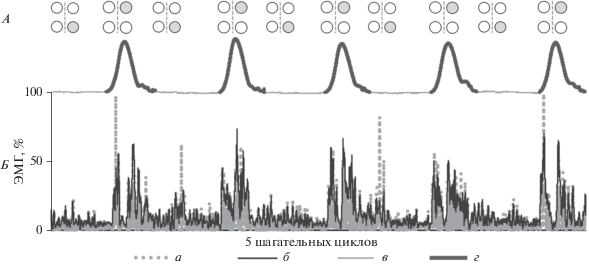
Возможность адресной активации моторных ядер сгибателей и разгибателей методом эпидуральной электрической стимуляции спинного мозга была показана в исследованиях с участием пациентов с травмой спинного мозга в [4]. В исследованиях с участием здоровых добровольцев продемонстрировали, что ЧЭС на 2 см правее или левее центральной оси позвоночника на уровне позвонков L1–L2 увеличивала силу разгибания в коленном и голеностопном суставах на стороне стимуляции [5]. Для ЧЭС фиксировали накожно 4 катода (ø 3 см): 1) для активации разгибателей на ~2.5 см правее и левее оси позвоночника по ходу корешков спинного мозга между позвонками L1–L2, 2) для активации сгибателей – на ~2.5 см правее и левее оси позвоночника по ходу корешков спинного мозга между позвонками T11–T12 (рис. 1, А). Два анода (5 × 10 см2) размещали накожно симметрично над гребнями подвздошных костей. Для стимуляции использовали Биостим/Неостим-5 (ООО “Косима”, Россия). ЧЭС проводили монополярными модулированными импульсами тока (длительность 1 мс, модуляция 5 кГц) с частотой 15 Гц (активация разгибателей) и 30 Гц (активация сгибателей). Диапазон использованных токов 15–90 мА. Подробнее материалы и метод ЧЭС описаны в [3].
ЧЭС корешков на уровне T11–12 и L1–2 проводили во время фаз переноса и опоры ипсилатеральной ноги, соответственно. Для детекции фаз шага на передние поверхности бедер эластичными повязками крепили датчики-гироскопы, в состав которых входили трехосный гироскоп и трехосный акселерометр в одной микросхеме LSM6DSL (ST Microelectronics, Швейцария), микроконтроллер MSP430F5510 (Texas Instruments, США), радиомодуль, аккумулятор. Момент постановки стопы на опору и отрыва от опоры определяли по смене знака производной угловой скорости бедра, которую регистрировал гироскоп.
Для регистрации кинематических характеристик движений ног использовали видеосистему (Qualisys, Швеция). На двух ногах регистрировали поверхностную ЭМГ сгибателя и разгибателя коленного сустава – мышц бедра (m. gastrocnemius lat. (GL), m. tibialis ant. (TA)) и сгибателя и разгибателя голеностопного сустава – мышц голени (m. biceps fem (BF), m. vastus lat. (VL)). Для этого использовали телеметрический электронейромиограф (ME 6000 MegaWin, Финляндия). Сигналы ЭМГ были синхронизированы с отметками стимуляции и с сигналами видеосистемы. Для анализа ЭМГ фазы шага определяли по перемещениям маркера-отражателя, закрепленного на обуви над пятым пальцем стопы. Материалы и методы электромиографии и видеофиксации ходьбы, а также детали их синхронизации подробно описаны в работе [6].
При ЧЭС корешков спинного мозга ЭМГ зашумлена артефактами стимуляции. Все данные ЭМГ, в том числе и зарегистрированные без ЧЭС, фильтровали с помощью оригинального программного обеспечения. Принцип работы фильтра состоит в поиске паттерна, характерного для артефакта стимуляции, и замена фрагмента ЭМГ, содержащего наводку, на динамическое среднее. При этом вариационный ряд ЭМГ оставался практически не затронут. Дальнейшую обработку и анализ ЭМГ проводили в среде RStudio. Сигналы ЭМГ-активности инвертировали в положительную область и нормировали, затем строили огибающую, ЭМГ синхронизировали для каждой фазы шага и считали среднее нормированной ЭМГ (рис. 1, Б) [7]. Огибающую строили методом скользящего среднего (окно 60) и нормировали методом шкалирования к 0 : 100. Нормированную огибающую каждого цикла аппроксимировали на 100 точек методом линейной интерполяции [8].
Так как золотой стандарт в расчетах КК отсутствует [1], применяли две хорошо известные формулы КК: модифицированную формулу Рудольфа КК(1) [9, 10] и формулу Латаша с коллегами КК(2) [1].
Формула КК(2), в отличие от формулы КК(1), не чувствительна к среднему уровню активности мышц – большие значения КК(2) отражают только одновременную активность мышц в сравниваемой паре, а маленькие значения – низкий уровень коактивации мышц в паре. Однако если разница между средней активностью мышц в сравниваемой паре не очень большая, то доля случайности в том, активность какой мышцы будет учтена в числителе КК(2), увеличивается, следовательно, вариабельность значений, рассчитанных по КК(2), велика. Последнего недостатка нет у формулы КК(1).
Во всех фазах шага отношение сгибателя и разгибателя бедра (BF и VL), сгибателя и разгибателя голени (TA и GL) рассчитывали по среднему значению ЭМГ за фазу. Данные, полученные при ЧЭС корешков справа и слева, анализировали вместе. Для определения эффекта активации корешков определяли относительную разницу (дельту) между коэффициентами коактивации при ходьбе с ЧЭС и при ходьбе без ЧЭС для каждого исследования (попарная разница, n = 15). Статистический анализ проводили с использованием t-критерия Стьюдента для связанных выборок. Достоверными считали различия при p < 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Коактивация мышц при ходьбе без стимуляции. Значения КК при ходьбе без ЧЭС, рассчитанные по обеим формулам, представлены на рис. 2, А, Б. Независимо от способа расчета получено, что в фазе переноса коактивация мышц голени достоверно меньше коактивации мышц бедра; в фазе опоры коактивация мышц голени и бедра не отличались. Коактивация мышц голени в фазе переноса достоверно меньше, чем в фазе опоры; значения КК мышц бедра не зависели от фазы шага.
Рис. 2.
Коактивация мышц-антагонистов при ходьбе по беговой дорожке. Коэффициенты коактивации, рассчитанные по формулам КК(1), КК(2), в фазах опоры и переноса при ходьбе без ЧЭС (А, Б), при ходьбе с ЧЭС на уровне L1 в фазе опоры (В, Г), при ходьбе с ЧЭС на уровне T11 в фазе переноса (Д, Е) и при чередующейся стимуляции на уровне L1 в фазе опоры и на уровне T11 в фазе переноса, как на рис. 1, Б (Ж, З). TA/GL и VL/BF – КК соответствующих пар мышц; показаны средние и доверительные интервалы; n = 15; * – p < 0.05.
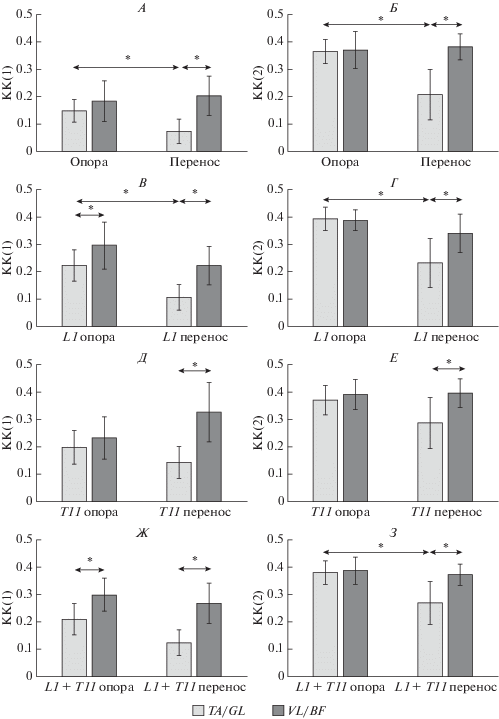
Результаты аналогичных исследований коактивации мышц голени и бедра в зависимости от фаз шага при ходьбе здоровых испытуемых по беговой дорожке опубликованы в [11]. Также получено, что КК мышц бедра были больше значений КК мышц голени и значения КК мышц голени в фазе опоры были больше, чем в фазе переноса. Однако в [11] значения КК мышц бедра, также, как и мышц голени, зависели от фазы шага и были больше в фазе опоры, чем в фазе переноса. Вероятно, причина различий в том, что в [11] испытуемые ходили со скоростью ~4.5 км/ч, в нашем исследовании – со скоростью в 3 раза меньше. Возможно, ходьба с меньшей скоростью по беговой дорожке связана с уменьшением амплитуды движений в коленном суставе, что проявляется в том, что значения КК мышц бедра (показатель жесткости коленного сустава) в фазе переноса не уменьшаются по отношению КК в фазе опоры. На основании анализа изменений КК в фазах шага при разной скорости ходьбы можно предположить, что произвольная регуляция скорости ходьбы происходит за счет регуляции амплитуды движений в коленном суставе.
Коактивация мышц при ходьбе с активацией разгибателей в фазе опоры. При ЧЭС на уровне L1 в фазе опоры коэффициенты коактивации мышц бедра и голени, рассчитанные по КК(1), были достоверно больше коэффициентов в фазе опоры при ходьбе без ЧЭС (рис. 3, А), при расчете по КК(2) положительный прирост соответствующих значений был недостоверным (рис. 3, Б). При ЧЭС на уровне L1 соотношения между величинами коэффициентов в фазах шага, рассчитанными по формулам КК(1) и КК(2), оставались такими же, как и при ходьбе без ЧЭС (рис. 2, А–Г). При расчете по КК(1) в фазе опоры (во время ЧЭС) усредненные коэффициенты мышц голени были достоверно меньше усредненных значений КК мышц бедра (рис. 2, В), что не наблюдали без ЧЭС (рис. 2, А). Такое же соотношение коэффициентов в фазе опоры наблюдали ранее при ходьбе здоровых испытуемых по беговой дорожке с большей, чем в наших исследованиях, скоростью [11]. Пропорция значений КК(1) мышц голени и бедра, соответствующая пропорции, полученной в [11], отражает увеличение частоты шагов (уменьшение длительности фазы опоры при постоянной скорости ходьбы) при ЧЭС на уровне L1 в фазе опоры [3]. Независимо от использованной для расчета формулы, значения КК в фазе переноса при ЧЭС на уровне L1 в фазе опоры достоверно не отличались от значений КК в фазе переноса при ходьбе без ЧЭС (рис. 3), что косвенно демонстрирует точность детекции фаз шага “нейропротезом” [3].
Рис. 3.
Влияние пространственно-временнóй чрескожной стимуляции спинного мозга на коактивацию мышц. А, Б – разница (дельта) между относительным изменением КК, при ходьбе с ЧЭС и при ходьбе без ЧЭС, сосчитанными по формулам КК(1) и КК(2), попарное сравнение; L1 – ЧЭС на уровне L1 в фазе опоры; T1 – ЧЭС на уровне T11 в фазе переноса; L1 + T1 – ЧЭС на уровне L1 в фазе опоры и на уровне T11 в фазе переноса (рис. 1, Б); TA/GL и VL/BF – КК соответствующих пар мышц. n = 15. Показаны средние и доверительные интервалы. * – p < 0.05.

Коактивация мышц при ходьбе с активацией сгибателей в фазе переноса. При ЧЭС на уровне Т11 в фазе переноса коэффициенты коактивации мышц бедра и голени были достоверно больше коэффициентов в фазе переноса без ЧЭС при расчете по КК(1) (рис. 3, А), при расчете по КК(2) прирост значений достоверен для мышц голени, а для мышц бедра не достоверен (рис. 3, Б). Достоверное увеличение значений КК, рассчитанных по обеим формулам, демонстрирует, что при активации моторных ядер мышц-сгибателей в фазе переноса увеличивалась жесткость в голеностопном суставе. Соотношения между величинами коэффициентов, рассчитанными по обеим формулам КК, для ходьбы с ЧЭС на уровне Т11 отличаются от соотношений, которые наблюдали при ходьбе без ЧЭС, – отсутствует различие между коэффициентами мышц голени в фазе опоры и в фазе переноса; значения КК мышц голени при ЧЭС в фазе переноса такие же, как в фазе опоры без ЧЭС (рис. 2, Д, Е). Следовательно, при ЧЭС корешков сгибателей в фазе переноса жесткость в голеностопном суставе в этой фазе такая же, как и в фазе опоры при ходьбе без стимуляции – при этом способе ЧЭС при переносе ноги голеностопный сустав зафиксирован в недоразогнутом положении, в том же, в каком он был в фазе опоры без стимуляции. Вероятно, что активация моторных ядер мышц-сгибателей в фазе переноса у пациентов, перенесших инсульт, может преодолеть проблему отвисания стопы во время ходьбы, свойственную походке этих пациентов. Коэффициенты в фазе опоры при таком способе ЧЭС достоверно не отличались от коэффициентов в фазе опоры при ходьбе без ЧЭС (рис. 3), что еще раз доказывает точность определения фаз шага “нейропротезом”. Это важный результат, так как для детекции фаз шага в расчетах КК и в “нейропротезе” использованы разные технические устройства и методы.
Коактивация мышц при ходьбе с активацией сгибателей в фазе переноса и разгибателей в фазе опоры. При ходьбе, которая сопровождалась ЧЭС на уровне L1 в фазе опоры и ЧЭС на уровне Т11 в фазе переноса (как на рис. 1, Б), коэффициенты мышц голени и бедра в фазе опоры, рассчитанные по КК(1), были достоверно больше соответствующих коэффициентов при ходьбе без стимуляции, а значения КК(1) мышц голени в фазе переноса также достоверно отличались от соответствующих коэффициентов без ЧЭС (рис. 3, А). При расчете по формуле КК(2) достоверные отличия отсутствовали (рис. 3, Б). При расчете по КК(1) значения коэффициента мышц голени были достоверно меньше значений КК мышц бедра как в фазе переноса, так и в фазе опоры, а достоверные отличия между значениями коэффициента для мышц голени в фазе опоры и в фазе переноса, которые получили при ходьбе без ЧЭС, не обнаружили (рис. 2, А, Ж). Соотношения между коэффициентами мышц бедра и голени в фазах шага при ЧЭС, рассчитанными по формуле КК(2), были такими же, как и при ходьбе без ЧЭС (рис. 2, Б, З).
Достоверные отличия отсутствовали между коэффициентами коактивации при сравнении в парах ЧЭС L1/T11, L1/(L1 + T11), T11/(L1 + T11) как в фазе опоры, так и в фазе переноса. Отсутствие различий связано с тем, что при стимуляции в одной из фаз шага, меняя кинематику движения в этой фазе, вызывали также компенсационные изменения кинематики и в другой фазе шага, потому что скорость ходьбы была постоянной и была задана скоростью беговой дорожки.
Значения КК(1) в фазе опоры при ЧЭС корешков мышц-разгибателей в фазе опоры и в фазе переноса при ЧЭС корешков мышц-сгибателей в фазе переноса были достоверно больше, чем значения КК(1) в соответствующих фазах при ходьбе без ЧЭС как для мышц голени, так и для мышц бедра (рис. 3, А). Прирост КК(1) для мышц голени составил 5–7%, для мышц бедра – 11–12%. Как было отмечено [9], большие значения КК(1) соответствуют большой усредненной активности обеих мышц в анализируемой паре. Было доказано, что ЧЭС корешков разгибателей увеличивает амплитуду рефлекторного ответа мышц разгибателей со стороны стимуляции [5], у спинальных пациентов было показано, что эпидуральная электрическая стимуляция сегментов спинного мозга, в которых находятся ядра сгибателей, увеличивает активность мышц-сгибателей и также стимуляция сегментов спинного мозга, в которых находятся ядра разгибателей, увеличивает активность мышц-сгибателей для разгибателей [4]. Значения КК(2) при ЧЭС в тех же условиях увеличились недостоверно (рис. 3, Б); это связано с большой вариативностью значений, вычисляемых по этой формуле, которая проявляется при малой разнице между амплитудами ЭМГ сравниваемых мышц (см. Методика). Следовательно, изменения значений КК(2) при ЧЭС косвенно свидетельствуют о том, что во время стимуляции корешков активность мышц в анализируемой паре отличалась незначительно. Возможно, что увеличение усредненной активности мышц-антагонистов связано с явлением “наведения” (cross talking), т.е. с возможной передачей возбуждения между соседними мышцами при регистрации ЭМГ поверхностными электродами [12]. С одной стороны, для большинства мышц голени во время ходьбы испытуемых с разной скоростью показано, что разница между результатами регистрации методами поверхностной и внутримышечной электромиографии минимальна или недостоверна, т.е. явление “наведения” отсутствует [13, 14]. С другой стороны, для m. soleus и m. tibialis ant. в тех же условиях поверхностная электромиография регистрировала активность, а внутримышечная – демонстрировала отсутствие активности [14], и авторы исследования сделали вывод о том, что надо с осторожностью интерпретировать результаты поверхностной ЭМГ этих мышц. Следовательно, нельзя исключить, что явление “наведения” между соседними мышцами внесло свой вклад в прирост величин коактивации мышц-антагонистов, который мы зарегистрировали при ЧЭС.
ЗАКЛЮЧЕНИЕ
В результате исследования коактивации мышц-антагонистов в фазах шага в процессе адресного воздействии на исследуемые мышцы ЧЭС получено, что при ЧЭС моторных пулов мышц-разгибателей в фазе опоры коактивация мышц голени была достоверно меньше коактивации мышц бедра, что не наблюдали без стимуляции. Основываясь на опубликованных ранее данных, предполагаем, что это связано с зарегистрированным увеличением частоты шагания (уменьшение длины шага при постоянной скорости ходьбы [3]).
При ЧЭС на уровне моторных пулов мышц-сгибателей в фазе переноса получено, что коактивация мышц голени в фазе переноса такая же, как и в фазе опоры при ходьбе без стимуляции. Следовательно, при таком адресном воздействии ЧЭС голеностопный сустав зафиксирован в фазе переноса в недоразогнутом положении, в том же, в каком он был в фазе опоры без стимуляции.
При ходьбе, которая сопровождалась чередующейся ЧЭС: моторных пулов мышц-сгибателей в фазе переноса и моторных пулов мышц-разгибателей в фазе опоры, – жесткость голеностопного сустава в фазе переноса была такой же, как и в фазе опоры.
Независимо от способа расчета коактивации, значения КК в фазе переноса при ЧЭС во время фазы опоры не отличались достоверно от значений КК в фазе переноса при ходьбе без ЧЭС и, наоборот, при ЧЭС во время фазы переноса не отличались достоверно от значений КК в фазе опоры при ходьбе без ЧЭС. Это демонстрирует точность технологии детекции фаз шага в “нейропротезе”.
Впервые была определена коактивация мышц-антагонистов во время движений на фоне неинвазивной электрической стимуляции спинного мозга и выявлено увеличение коактивации всех исследуемых пар мышц по сравнению с движениями без стимуляции. В связи с этим необходимо проводить дальнейшие исследования, чтобы выяснить, связано ли это с “наведением” (cross talking) возбуждения между мышцами или с увеличением жесткости суставов, так как увеличение жесткости суставов связывают с риском падения, нарушения устойчивости при походке [11].
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены локальным биоэтическим комитетом Великолукской государственной академии физической культуры и спорта (ВЛГАФК) (Великие Луки).
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. НИР с ООО “Косима” № 2/2019 от 01.12.2019 в рамках проекта НТИ “Разработка нейростимуляционного устройства для регуляции двигательных функций у больных с последствиями острого нарушения мозгового кровообращения” и программы фундаментальных исследований Президиума РАН № 1.42.
Благодарности. Авторы благодарны Р.М. Городничеву, А.М. Пухову, Е.А. Пивоваровой, С.М. Иванову, В.В. Маркевич за помощь в экспериментальной работе.
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Latash M.L. Muscle coactivation: definitions, mechanisms, and functions // J. Neurophysiol. 2018. V. 120. № 1. P. 88.
Solopova I.A., Sukhotina I.A., Zhvansky D.S. et al. Effects of spinal cord stimulation on motor functions in children with cerebral palsy // Neurosc. Lett. 2017. V. 639. P. 192.
Городничев Р.М., Пухов А.М., Моисеев С.А. и др. Регуляция фаз шагательного цикла при неинвазивной электрической стимуляции спинного мозга // Физиология человека. 2021. Т. 47. № 1. С. 73. Gorodnichev R.M., Pukhov A.M., Moiseev S.A. et al. Regulation of stepping phases during noninvasive electrical spinal cord stimulation // Human Physiology. 2021. V. 47. № 1. P. 60.
Wagner F.B., Mignardot J.B., Le Goff-Mignardot C.G. et al. Targeted neurotechnology restores walking in humans with spinal cord injury // Nature. 2018. V. 563. № 7729. P. 65.
Calvert J.S., Manson G.A., Grahn P.J., Sayenko D.G. Preferential activation of spinal sensorimotor networks via lateralized transcutaneous spinal stimulation in neurologically intact humans // J. Neurophysiol. 2019. V. 122. № 5. P. 2111.
Gerasimenko Y., Gad P., Sayenko D. et al. Integration of sensory, spinal, and volitional descending inputs in regulation of human locomotion // J. Neurophysiol. 2016. V. 116. № 1. P. 98.
Guerrero J.E., Macias-Diaz J.A. Biosignal EMG: tools for electromyogram signals (EMG) analysis. R. Packag, 2015. Интернет ресурс: https://CRAN.R-project.org/package=biosignalEMG
Becker R. The new S language. CRC Press, 2018. ISBN 9 781 315 895 888.
Rudolph K.S., Axe M.J., Snyder-Mackler L. Dynamic stability after ACL injury: who can hop? // Knee Surgery, Sports Traumatology, Arthroscopy. 2000. V. 8. № 5. P. 262.
Martino G., Ivanenko Y.P., Serrao M. et al. Locomotor patterns in cerebellar ataxia // J. Neurophysiol. 2014. V. 112. № 11. P. 2810.
Acuña S.A., Francis C.A., Franz J.R., Thelen D.G. The effects of cognitive load and optical flow on antagonist leg muscle coactivation during walking for young and older adults // J. Electromyogr. Kinesiol. 2019. V. 44. P. 8.
De Luca C.J., Merletti R. Surface myoelectric signal cross-talk among muscles of the leg // Electroencephalogr. Clin. Neurophysiol. 1988. V. 69. № 6. P. 568.
Vieira T.M., Botter A., Muceli S., Farina D. Specificity of surface EMG recordings for gastrocnemius during upright standing // Scientific Reports. 2017. V. 7. № 1. P. 1.
Péter A., Andersson E., Hegyi A. et al. Comparing surface and fine-wire electromyography activity of lower leg muscles at different walking speeds // Frontiers in physiology. 2019. V. 10. Article 1283. https://doi.org/10.3389/fphys.2019.01283
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека