

Физиология человека, 2021, T. 47, № 4, стр. 115-123
Потенциалы, связанные с событиями, в тесте на сравнение пар слов в зрительной и слуховой модальности
И. С. Никишена 1, *, В. А. Пономарев 1, Ю. Д. Кропотов 1
1 ФГБУН Институт мозга человека имени Н.П. Бехтеревой РАН
Санкт-Петербург, Россия
* E-mail: nikishena@mail.ru
Поступила в редакцию 14.07.2020
После доработки 16.09.2020
Принята к публикации 25.11.2020
Аннотация
Целью исследования было изучение особенностей работы мозга при сравнении речевых сигналов в условиях кроссмодального взаимодействия. Исследовались связанные с событиями потенциалы (ПСС) у 166 здоровых испытуемых в двухмодальном двустимульном GO-NOGO тесте, в котором первым стимулом было напечатанное слово, а вторым – произнесенное слово. Показано, что при ожидании произносимого слова после предъявления напечатанного слова регистрируется активация лобной-височной зоны левого полушария (F7, F3), предположительно, связанная с формированием в памяти фонологического образа слова. После предъявления второго (произносимого) слова, при его совпадении с напечатанным, регистрируется активация с максимумом в задневисочных отведениях (Т5, Т6) в интервале времени 370–500 мс, а в случае несовпадения – в интервале 590–800 мс с максимумом в затылочных отведениях (О1, О2). Эти колебания потенциала, предположительно, связаны с процессами сличения вновь полученной информации от слуховой системы с фонологическим образом слова, хранящимся в рабочей памяти.
Потенциалы, связанные с событиями (ПСС), представляет собой суммарную электрическую активность головного мозга в ответ на стимул. Метод признан удобным, поскольку дает хорошее временнóе разрешение при анализе процессов обработки стимула в зависимости от решаемой задачи. В литературе представлено значительное количество данных на основе анализа ПСС, связанных фактически со всеми этапами обработки информации (восприятие, память, внимание и прочие) [1, 2].
Начальным этапом в подготовке исследований методом ПСС является выбор дизайна теста. Предъявляемый участнику исследования тест моделирует ситуацию, в которой будут задействованы те или иные мозговые системы. В частности, широкое распространение получила GO-NOGO-парадигма [1–4]. В этой парадигме должны быть соблюдены некоторые условия: во-первых, наличие GO-стимула, требующего ответной реакции, вероятность этого стимула должна быть достаточно высока; во-вторых, наличие стимула, после которого необходимо сдержать импульсивную ответную реакцию; в-третьих, у выполняющего тест человека должно быть время на подготовку ответа после стимула. Такие тесты используются для изучения мозговых систем обеспечения внимания и контроля поведения, включающих рабочую память.
Существует двустимульный вариант GO-NOGO-парадигмы, в котором в каждой пробе предъявляются последовательно два типа стимулов. Первый стимул может быть как просто предупреждающим о начале пробы, так и несущим более сложную информацию для последующего сравнения со вторым стимулом в пробе. Второй стимул в пробе несет информационную нагрузку и требует ответной реакции участника исследования. Такие тесты дают возможность изучать два вида феноменов – ПСС между двумя стимулами в пробе и ПСС после запускающего стимула.
К первой группе феноменов относится медленное отрицательное колебание – условно негативная волна (УНВ, contingent negative variation – CNV) [5]. Это колебание регистрируется перед предъявлением второго стимула, при достаточно длинном межстимульном интервале в нем выделяются две разные волны – ранняя О-волна УНВ и поздняя Е-волна УНВ. Ранняя УНВ связана с ориентировочной реакцией на первый стимул и обработкой содержащейся в нем когнитивной информации, регистрируется в проекции лобных отделов мозга [5]. Е-волну УНВ определяют как волну ожидания или готовности к действию после второго стимула, это колебание модулируется вниманием, этот компонент ПСС имеет максимальную величину в центральных и затылочных отведениях [6–9]. В случае тестового задания, без подготовки к двигательному ответу, может быть зарегистрирована низкоамплитудная волна “stimulus preceding negativity” (SPN), которую связывают с готовностью к восприятию информации о втором стимуле в паре, например, сравнению с использованием рабочей памяти [10–14].
Ко второй группе феноменов ПСС, регистрируемых после второго стимула, относятся компоненты, связанные с восприятием стимулов и контролем поведения [2], включая вовлечение или подавление действий.
Помимо этого, наличие в условии теста двух стимулов дает возможность изучать процессы сличения информации и формирования ответной реакции при сравнении двух сигналов, подаваемых на разные сенсорные входы, например, в зрительной и звуковой модальности.
Процессы запоминания и сличения информации являются основополагающими механизмами работы мозга [15–22]. Усложняет понимание этих механизмов то, что информация поступает по разным сенсорным каналам, а в случае человека – еще и в виде символов (букв, слов, знаков). Вопрос о том, как мозговые системы осуществляют кроссмодальное взаимодействие, имеет не только теоретическое, но и прикладное значение [23, 24]. Можно выделить несколько режимов сличения информации, во-первых, мультисенсорное взаимодействие, во-вторых, сличение отдельных характеристик стимулов разной модальности со следом, хранящимся в рабочей памяти, в-третьих, сравнение категорий стимулов на основе категоризации. Мультисенсорное взаимодействие относится к процессам восприятия, когда перцептивные системы связывают и интегрируют входные данные для создания единого образа [25–28]. Под категоризацией подразумевается процесс отнесения стимула к какому-либо классу по совокупности признаков [29]. По-видимому, существует еще один вариант, суть которого состоит в том, что до категоризации второго стимула может происходить сравнение в рабочей памяти характеристик разномодальных стимулов, которые разделены по времени. Примером такого сличения информации является сравнение речевых сигналов, первый их которых – напечатанное слово, а второй – произнесенное. В этом случае можно предположить, что после первого стимула происходит формирование в памяти его “фонологического аналога” [15, 16, 23, 24]. И тогда одной из возможных “оптимальных” стратегий выполнения задачи, будет сравнение второго стимула с “фонологическим аналогом” первого. К сожалению, физиологические механизмы этого сличения, если оно действительно существует, мало изучены.
В данной статье представлены результаты пилотных исследований, полученные при сравнении ПСС при предъявлении напечатанного слова на экране монитора (первый, “word”-стимул) и произнесенного слова (второй, “sound”-стимул). Задачей испытуемого было запомнить первый стимул (“word”) и сравнить с произнесенным словом. Если слова были одинаковыми, следовало нажать на кнопку, если разными – на кнопку не нажимать. Можно предположить, что для того, чтобы запомнить слово, человек его читает и повторяет при помощи внутренней речи, чтобы создать и запомнить фонологический образ напечатанного слова. После предъявления второго стимула (“sound”-стимул) испытуемый сравнивает его со следом в памяти для формирования ответной реакции. Таким образом, предметом исследования были события, регистрируемые непосредственно перед вторым стимулом и события после второго стимула, предшествующие ответной реакции. Эти результаты представляют интерес, поскольку полученные данные показывают особенности мозговой организации ответа, связанной с функциями сохранения речевого образа и сличения информации.
МЕТОДИКА
В исследовании принимали участие 166 здоровых испытуемых в возрасте от 7 до 16 лет (мужского пола – 91, женского пола – 75 чел.). Испытуемых подбирали по следующим критериям: 1) перинатальный период протекал без осложнений, 2) отсутствие неврологических и психиатрических заболеваний, травм мозга, судорожной активности в анамнезе, 3) нормальное психическое и физическое развитие, без нарушений слуха, нормальное или скорректированное зрение, 4) средняя или лучшая успеваемость в школе, 5) отсутствие медикаментозной терапии на момент исследования.
Двухмодальный двустимульный тест. Участникам исследования предъявляли два типа стимулов – зрительные и слуховые. Испытуемого располагали в кресле перед монитором, на экране которого демонстрировались зрительные стимулы, слуховые стимулы воспроизводилиcь через динамики. В тесте было 20 слов в печатном виде и 20 слов, произнесенных вслух. Тест выполнялся на родном языке участников исследования. Аудиофайлы были записаны носителем языка мужским голосом. Изображения с угловыми размерами, порядка 3.8°, предъявляли в центре экрана на белом фоне. Между предъявлением изображений на экране демонстрировался белый фон. Презентация изображений не сопровождалась миганием экрана монитора. Для предъявления стимулов использовали программу Psytask (Россия).
Тест состоял из 200 проб. Продолжительность теста составляла 6 мин 20 с. В каждой пробе стимулы предъявляли парами. Первым в паре предъявляли зрительный стимул в виде напечатанного слова (“word”-стимул). Вторым в паре был слуховой стимул в виде произнесенного слова (“sound”-стимул). Длительность предъявления зрительного стимула – 200 мс, интервал между стимулами в паре – 1000 мс, длительность слухового стимула 700 мс, а интервал между началами пар стимулов – 3100 мс.
Все пробы были разделены на две равные группы, в зависимости от сочетания стимулов в паре. Использовали следующие сочетания пар стимулов: одинаковые (“word”-стимул – парный “sound”-стимул), различающиеся (“word”-стимул – отличающийся “sound”-стимул). Испытуемые нажимали на кнопку правой рукой как можно точнее и быстрей, если произнесенное слово соответствовало печатному – GO-условие, таких пар было 50%. В случае, если произнесенное слово отличалось от напечатанного, испытуемый должен был воздержаться от нажатия кнопки (NOGO-условие 50% проб). Слова в парах были подобраны следующим образом: они различались одной начальной буквой в написании слова и одной фонемой в начале слова при произнесении слова. Пример GO-условия – DINO-DINO, пример NOGO-условия – DINO-KINO. Пробы предъявляли в квазислучайном порядке.
Пробы, в которых испытуемые неправильно выполняли задание, исключали из анализа. Перед началом исследования волонтерам предоставляли несколько десятков проб для тренировки. В ходе теста делали одну паузу длительностью 1–2 мин для отдыха.
ЭЭГ была записана с помощью компьютерного электроэнцефалографа “Мицар-ЭЭГ” (Россия). Для постановки электродов использовали международную систему 10-20. Референтные электроды крепили на мочки ушей, использовали объединенные электроды. Заземляющий электрод был закреплен в точке Fpz. Сопротивление электродов не превышало 5 кОм. Частота квантования сигналов составляла 250 Гц. Параметры ФВЧ и ФНЧ – 0.53 (τ = 0.3 с) и 50 Гц соответственно. Сетевую электромагнитную помеху подавляли с использованием цифрового режекторного фильтра с полосой непропускания 45–55 Гц.
Данные обрабатывали с использованием программы WinEEG. Коррекцию артефактов моргания выполняли с помощью метода, предложенного R.N. Vigário [30], использующего разложение на независимые компоненты. Также пробы, содержащие артефакты, исключали из дальнейшего анализа. В частности было принято (эмпирически подобрано), что пробы, в которых: 1) потенциалы более 50 мкВ для электродов Fp1 и Fp2 и более 100 мкВ для остальных электродов; 2) медленные волны в диапазоне от 0.53 до 1 Гц, с амплитудой более 50 мкВ; 3) быстрые колебания сигнала в интервале частот от 20 до 35 Гц, с амплитудой более 35 мкВ содержали артефакты. ПСС были рассчитаны методом усреднения отдельно для каждого из двух условий. Все компоненты, описываемые в результатах, были выделены при помощи t-критерия для парных измерений. Для снижения шума, записи, в которых было менее 30 проб для каждого из условий, удаляли из исследования.
Зрительный двустимульный тест. При сравнении и обсуждении полученных данных использовали зрительный GO-NOGO тест, результаты обработки которого публиковались в работах [31, 32]. Зрительный GO-NOGO тест представляет собой двустимульный тест, в котором в качестве стимулов используются изображения, относящиеся к разным категориям – животные (Ж), растения (Р). Варианты предъявления стимулов в пробе: Ж-Ж, Ж-Р, Р-Р, Р-Ж. Все пары стимулов предъявляли с равной вероятностью, порядок пар был квазислучайным. Интервал между первым и вторым стимулов составлял 1000 мс, длительность предъявления зрительного стимула на экране – 100 мс, длительность пробы 3000 мс. Проба, требующая ответного нажатия на кнопку – Ж-Ж. Если участник исследования отвечал неправильно, такую пару исключали из обработки данных. Для каждого из четырех вариантов предъявления проб ПСС вычисляли отдельно. Записи, в которых было менее 30 проб для каждого из условий, удаляли из исследования.
Для сравнения в настоящем исследовании были выбраны условия Ж-Ж (GO-условие), Ж-Р (NOGO-условие). Из описания зрительного GO-NOGO теста видно, что условия двух тестов не были идентичными, поэтому сравнивался не весь компонентный состав после первого стимула, а только события, непосредственно предшествующие второму стимулу. В этот временнóй период характеристики первого стимула влияют в наименьшей степени, и происходят процессы подготовки к приему и сравнительной обработке второго стимула в соответствии (что особенно важно) с поставленной задачей.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Поведенческие показатели. Количество пропусков значимых стимулов – 9.63% (SD = 7.93), время реакции – 876 мс (SD = 134); количество ложных тревог – 3.29% (SD = 2.88).
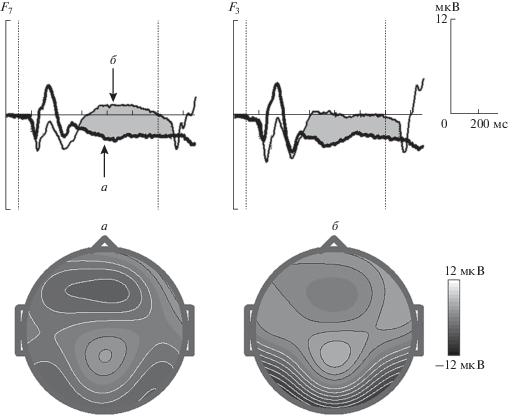
Компоненты потенциалов, связанных с событиями после предъявления первого стимула. Перед предъявлением “sound”-стимула наблюдаетcя медленное отрицательное отклонение потенциала в интервале 500–1100 мс от начала первого стимула, максимально выраженное в отведениях F7, F3 и Fz (рис. 1) со статистической значимостью p < 10–6, которая оценивалась по отношению к среднему значению электрического потенциала в интервале перед первым стимулов.
Рис. 1.
Усредненные по группе ПСС для электродов F7, F3 в GO-условии после первого стимула в двух тестах. а – Двухмодальный тест, б – зрительный тест. Стрелками указано позднее отрицательное колебание перед вторым стимулом. По оси абсцисс – время, мс, по оси ординат – величина потенциала, мкВ. Момент начала стимула указан вертикальной пунктирной линией. Топографии, определенные для максимальных значений потенциала в двухмодальном (а) и зрительном (б) тестах.
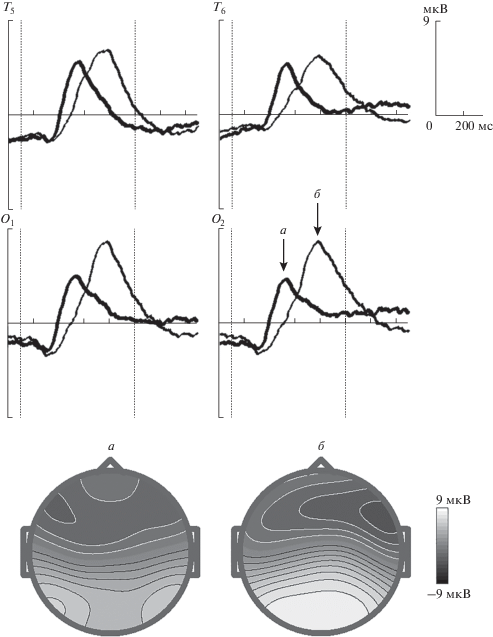
Компоненты потенциалов, связанных с событиями после предъявления второго стимула. На рис. 2 представлены усредненные потенциалы в референтном монтаже для обоих условий теста. На рис. 2 видно, что различия после второго стимула регистрируются в интервале 350–800 мс. Ответ на совпадение слов регистрируется раньше, чем на несовпадение. При совпадении напечатанного слова с услышанным, возникает положительное отклонение потенциала в интервале 370–500 мс, с максимумом в 428 мс в задневисочных отведениях Т5 и Т6. В случае несовпадения регистрируется положительная волна, достигающая максимума позднее, в интервал времени 590–800 мс, с максимумом в 680 мс в затылочных отведениях (О1, О2). Статистическая значимость этих волн по отношению к среднему значению в интервале перед первым стимулом p < 10–6. Статистическая значимость различия сигналов в GO- и NOGO-условиях в интервале 300–450 мс p < 10–5 для затылочных отведений и p < 10–6 – для височных.
Рис. 2.
Усредненные по группе ПСС после второго стимула в двухмодальном тесте. а – GO-условие, совпадение слов, б – NOGO-условие, несовпадение слов для электродов Т5, Т6, О1, О2. Стрелками указаны волны в GO- и NOGO-условии. По оси абсцисс – время, мс, по оси ординат – величина потенциала, мкВ. Начало и конец второго стимула указаны вертикальными пунктирными линиями. Топографии, определенные для максимальных значений в GO-условии (а), в NOGO-условии (б).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Существует несколько теорий относительно процесса сравнения кросс-модальных стимулов [15, 24]. Например, трехкомпонентная модель рабочей памяти предполагает отдельное хранение вербальной, пространственной информации и информации об объектах. В соответствии с трехкомпонентной моделью рабочей памяти, после предъявления зрительного образа слова, стимул преобразуется в фонологический аналог (articulatory rehearsal) для дальнейшей обработки и удержания в рабочей памяти [24, 33].
Авторы настоящей статьи сконструировали двустимульный тест, требующий от участника исследования сравнения двух стимулов в каждой пробе и ответной реакции. В двухмодальном тесте было зарегистрировано отрицательное отклонение после первого стимула в интервале 500–1100 мс (рис. 1). Фокус активности приходится на лобное центральное отведение (Fz), но в дополнение к этому наблюдается активация в проекции лобных отделов левого полушария (F3, F7) (рис. 1). В GO-NOGO тестах подобного дизайна после первого стимула в интервале 500 мс и до предъявления второго стимула развивается негативное отклонение УНВ [5, 9]. О-волна УНВ не имеет выраженной латерализации. Что касается поздней УНВ, то Е-волна УНВ может иметь асимметрию, которая приходится на центральные отделы и зависит от используемой в ответе руки [8], а отрицательное колебание SPN регистрируется в лобных и затылочных отделах в слуховой и зрительной модальностях соответственно, но не имеет асимметрии в локализации [34]. Следовательно, ни одно из этих колебаний в полной мере не соответствует полученному нами медленному отрицательному колебанию ПСС после первого стимула в “двухмодальном” тесте.
Физиологический смысл этого отрицательного колебания с максимумом в проекции лобных отделов левого полушария можно объяснить следующим образом. В конструкции двухмодального теста первый стимул был зрительным, и предположительно, для создания модели стимула, происходит его перевод в фонологический образ (articulatory rehearsal, внутреннее произнесение слова), и соответственно задействуются отделы мозга, связанные с артикуляционным аппаратом и лежащие в проекции лобных отведений слева (зона Брока).
Результаты исследования показали, что различия в обработке сигнала после второго стимула в GO- и NOGO-условиях регистрируются в виде положительных колебаний электрического потенциала в ответ на второй стимул с латентным периодом 428 и 680 мс. Это значительно раньше, чем время реакции (876 ± 20.6 мс). Важно учитывать, что в предложенном тесте длина второго стимула составляет 700 мс, поэтому наблюдается увеличение латентного периода когнитивных компонентов по сравнению с данными других авторов. Топографии рассматриваемых положительных отклонений потенциала после второго стимула отличаются от стандартной GO-NOGO-парадигмы. В качестве примера можно привести топографию компонента Р3 в NOGO-условии (рис. 3) в зрительном двустимульном тесте у детей [31, 32]. Из рис. 3 видно, что этот NOGO-компонент имеет центрально-теменное распределение, в то время как в двухмодальном тесте при несовпадении слов регистрируется положительное колебание в затылочных областях. На основе сравнения представленных в этом исследовании экспериментальных данных о локализации компонент ПСС, а также данных литературы и собственных [32], можно предположить, что полученные нами компоненты не соответствуют по локализации Р3NOGO-компонентам.
Рис. 3.
Усредненные по группе ПСС после второго стимула в зрительном тесте. а – GO-условие, совпадение зрительных образов, б – NOGO-условие, несовпадение зрительных образов для электродов Cz и Pz. По оси абсцисс – время, мс, по оси ординат – величина потенциала, мкВ. Стрелками указаны волны Р3GO и Р3NOGO. Начало и конец второго стимула указаны вертикальными пунктирными линиями. Топографии, определенные для максимальных значений GO-условии (а), NOGO-условии (б).

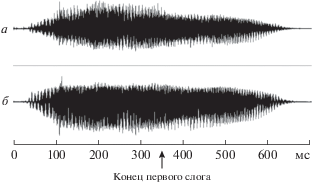
Длина второго стимула составляет 700 мс. На рис. 4 представлены все 20 вариантов слов, наложенных друг на друга, стрелкой указан конец первого слога. Из рис. 4 видно, что окончание первого слога приходится примерно на 350 мс, начало первого из рассматриваемых колебаний потенциала приблизительно соответствует этому времени, т.е. еще до окончания слова. Значит полный образ слова, и, следовательно, категория объекта (действия) еще не могут быть сформированы. Эти данные показывают, что, вероятно, наблюдаемые феномены не являются вариантом компонента P3, а связанны с операцией сравнения предъявляемого стимула с внутренней репрезентацией, хранящейся в рабочей памяти.
Рис. 4.
Изображение, полученное наложением друг на друга 20-и графиков оцифрованных слов (“sound”-стимулов). Стрелкой указан конец первого слога. По оси абсцисс – время, в мс. а – левый динамик, б – правый динамик.
По данным литературы и исследованиям авторов настоящей статьи, в задаче на детекцию рассогласования стимулов в теменных отведениях в тесте GO-NOGO регистрируется колебание Р2 (c пиковой латентностью 200–300 мс). В литературе описан компонент Р2 в NOGO-условии в тесте на предъявление зрительных стимулов с разными физическими характеристиками, но относящиеся к одной категории (GO-условие), или к разным категориям (NOGO-условие) [18]. Этот компонент наблюдается в задневисочных отведениях и отражает процессы обнаружения конфликта при сравнении входящей информации с образом, хранящимся в рабочей памяти. В двухмодальном тесте различия наблюдаются в проекции височно-затылочных отделов мозга в GO-условии. Можно предположить, что для сравнения речевых стимулов на раннем этапе сравнения, используются те же механизмы мозга, что и неречевых зрительных и слуховых образов, и основная роль принадлежит указанным или близлежащим отделам.
Сравнимые данные были получены С.В. Синицыным [35], в тесте на сравнение зрительных паттернов и фонологических образов у детей, процессы обработки стимулов отражались в положительных колебаниях потенциала в задних теменных и височных отделах в интервале 248–528 мс, автор также связывает эти колебания с процессами в рабочей памяти.
В соответствии с описанием двухмодального теста, отдельной задачи понять смысл слова не ставилось, поэтому одной из возможных стратегий, которую, вероятно, принимали большинство испытуемых, была следующей. После первого стимула участник исследования, вероятно, переводил напечатанное слово в фонологический “проговариваемый” вариант, после второго стимула достаточно было сравнения звукового сочетания, без учета смысловой нагрузки. Это утверждение, естественно, не исключает дальнейшей или параллельной обработки смысловых характеристик сигнала, которая могла продолжаться и после нажатия на кнопку. Такая стратегия поведения подтверждается коротким временем реакции – около 763 мс, окончание произносимого слова приходилось на 650 мс (рис. 4). Это позволило давать ответ быстрее, но привело к большому количеству ошибок по сравнению с NOGO-условием.
Латентный период положительного компонента в NOGO-условии больше, чем GO, т.е. формирование мозгового ответа на NOGO-условие требует более длительной обработки сигнала. В NOGO-условии было допущено меньше ошибок (GO-условие 9.63% vs NOGO 3.31%). Большее количество пропусков одинаковых пар нельзя объяснить неточным опознанием, тогда и разные пары слов опознавались как одинаковые, а количество ложных нажатий на NOGO-пробы было бы сравнимо с количеством пропусков значимых стимулов. Возможно, эволюционно выгодно рассматривать схожие стимулы в условиях дефицита времени как разные и формировать на них ориентировочную реакцию.
ЗАКЛЮЧЕНИЕ
Результаты анализа ПСС в двухмодальном тесте с предъявлением слов показали несколько процессов, которые протекают при обработке стимула в виде слова. После предъявления первого “word”-стимула и перед предъявлением второго “sound”-стимула наблюдается активация в проекции лобных отделов левого полушария в зоне Брока. Эта активация связана с формированием в рабочей памяти фонологического аналога предъявленного слова.
После второго стимула в двух условиях регистрируются положительные компоненты ПСС. Эти колебания предположительно связанны с механизмами сличения сигналов в рабочей памяти. Компонент в GO-условии регистрируется раньше по времени и имеет задневисочную локализацию. Компонент ПСС в NOGO-условии регистрируется позднее с максимумом в затылочных отведениях. Эти характеристики отражают особенности сличения сигналов в рабочей памяти в ситуации вовлечения в действие и подавления действия.
Полученные результаты заслуживают внимания, поскольку затрагивают сложные вопросы кросс-модального взаимодействия, памяти и речевых функций, в частности, возможности участия рабочей памяти на достаточно ранних этапах обработки слова до его смыслового анализа и категоризации. Данные могут быть полезны исследователям, занимающимся указанными вопросами. Однако полученные результаты являются не окончательными и предполагают продолжить исследования в данном направлении.
Этические нормы. Исследование проведено в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены Комитетом по этике Института мозга человека им. Н.П. Бехтеревой РАН (Санкт-Петербург) и Местным комитетом по этике кантона Граубюнден (Швейцария).
Информированное согласие. Законный представитель каждого участника исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. Работа выполнена в рамках государственного задания Минобрнауки по теме № АААА-А19-119101890066-2.
Благодарности. Авторы выражают благодарность Dr. Andreas Muller, Детский центр г. Кур (Швейцария) за предоставленные записи ПСС в GO-NOGO тесте.
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Кропотов Ю.Д. Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия / Донецк: Издатель Заславский Ю.А., 2010. 512 с.
The Oxford handbook of event-related potential components / Eds. Luck S.J., Kappenman E.S. Oxford: Oxford University Press, 2011. 642 p.
Verleger R., Paehge T., Kolev V. et al. On the relation of movement-related potentials to the go/no-go effect on P3 // Biol Psychol. 2006. V. 73. № 3. P. 298.
Vuillier L., Bryce D., Szücs D., Whitebread D. The maturation of interference suppression and response inhibition: ERP analysis of a cued Go/Nogo Task // PLoS One. 2016. V. 11. № 11. P. e0165697
Walter W.G., Cooper R., Aldridge V.J. et al. Contingent negative variation: an electric sign of sensorimotor association and expectancy in the human brain // Nature. 1964. V. 203. P. 380.
Brunia C.H., van Boxtel G.J. Wait and see // Int. J. Psychophysiol. 2001. V. 43. № 1. P. 59.
Tecce J.J. Contingent negative variation (CNV) and psychological processes in man // Psychol. Bull. 1972. V. 77. № 2. P. 73.
Gomez C.M., Delinte A., Vaquero E. et al. Current source density analysis of CNV during temporal gap paradigm // Brain Topogr. 2001. V. 13. № 3. P. 149.
Taylor B.K., Gavin W.J., Davies P.L. The test-retest reliability of the visually-evoked contingent negative variation (CNV) in children and adults // Dev. Neuropsychol. 2016. V. 41. № 3. P. 162.
Brunia C.H. Movement and stimulus preceding negativity // Biol. Psychol. 1988. V. 26. № 1–3. P. 165.
Qiao Z., Geng H., Wang Y., Li X. Anticipation of uncertain threat modulates subsequent affective responses and covariation bias // Front. Psychol. 2018. V. 11. № 9. A. 2547.
Johnen A.-K., Harrison N.R. Level of uncertainty about the affective nature of a pictorial stimulus influences anticipatory neural processes: An event-related potential (ERP) study // Neuropsychologia. 2020. V. 146. A. 107 525.
Michalowski J.M., Pane-Farre C.A., Low A., Hamm A.O. Brain dynamics of visual attention during anticipation and encoding of threat- and safe-cues in spider-phobic individuals // Soc. Cognit. Affect. Neurosci. 2015. V. 10. № 9. P. 1177.
Tanovic E., Pruessner L., Joormann J. Attention and anticipation in response to varying levels of uncertain threat: an ERP study // Cognit. Affect Behav. Neurosci. 2018. V. 18. № 6. P. 1207.
Baddleley A.D., Hitch G.J. Working memory / Recent advances in learning and motivation // Ed. Bower G. N.Y.: Academic Press, 1974. V. 8. P. 47.
Baddeley A.D. Working memory: theories, models, and controversies // Annu. Rev. Psychol. 2012. V. 63. P. 1.
Miller E.K., Lundqvist M., Bastos A.M. Working memory 2.0 // Neuron. 2018. V. 24. № 100(2). P. 463.
Кропотов Ю.Д., Пономарев В.А., Пронина М.В., Полякова Н.В. Эффекты повторения и рассогласования стимулов в сенсорных зрительных компонентах потенциалов, связанных с событиями // Физиология человека. 2019. Т. 45. № 4. С. 5. Kropotov Y.D., Ponomarev V.A., Pronina M.V., Polyakova N.V. Effects of Repetition and Stimulus Mismatch in Sensory Visual Components of Event-related Potentials // Human Physiology. 2019. V. 45. № 4. P. 349.
Chai W.J., Abd Hamid A.I., Abdullah J.M. Working memory from the psychological and neurosciences perspectives: A review // Front. Psychol. 2018. V. 9. P. 401.
D’Esposito M., Postle B.R. The cognitive neuroscience of working memory // Rev. Psychol. 2015. V. 3. № 66. P. 115.
Nee D.E., D’Esposito M. The representational basis of working memory // Curr. Top. Behav. Neurosci. 2018. V. 37. P. 213.
Clark R.E. A history and overview of the behavioral neuroscience of learning and memory / Behavioral Neuroscience of Learning and Memory // Eds. Clark R.E., Martin S. Curr. Top. Behav. Neurosci. Cham: Springer, 2017. V. 37. P1.
Ruchkin D.S., Berndt R.S., Johnson R.Jr. et al. Modality-specific processing streams in verbal working memory: evidence from spatio-temporal patterns of brain activity // Brain Res. Cogn. Brain Res. 1997. V. 6. № 2. P. 95.
Peters J., Suchan B., Zhang Ya., Daum I. Visuo-verbal interactions in working memory: evidence from event-related potentials // Brain Res. Cogn. Brain. Res. 2005. V. 25. № 2. P. 406.
Hidaka S., Teramoto W., Sugita Y. Spatiotemporal processing in crossmodal interactions for perception of the external world: A review // Front. Integr. Neurosci. 2015. V. 22. № 9. P. 62.
Stock A.K., Gohil K., Huster R.J., Beste C. On the effects of multimodal information integration in multitasking // Sci Rep. 2017. V. 7. № 1. A. 4927.
Tang X., Wu J., Shen Y. The interactions of multisensory integration with endogenous and exogenous attention // Neurosci. Biobehav. Rev. 2016. V. 61. P. 208.
Sanders P., Thompson B., Corballis P., Searchfield G. On the timing of signals in multisensory integration and crossmodal interactions: A scoping review // Multisens. Res. 2019. V. 1. № 32(6). P. 533.
Kropotov J.D., Ponomarev V.A. Differentiation of neuronal operations in latent components of event-related potentials in delayed match-to-sample tasks // Psychophysiology. 2015. V. 52. № 6. P. 826.
Vigário R.N. Extraction of ocular artefacts from EEG using independent component analysis // Electroencephalogr. Clin. Neurophysiol. 1997. V. 103. № 3. P. 395.
Кропотов Ю.Д. Современная диагностика и коррекция синдрома дефицита внимания. СПб.: ЭЛБИ-СПб, 2005. 148 с.
Kropotov J.D., Ponomarev V.A., Tereshchenko E.P. et al. Effect of aging on ERP components of cognitive control // Front. Aging Neurosci. 2016. V. 8. P. 69.
Smith E.E., Jonides J. Working memory: A view from neuroimaging // Cogn. Psychol. 1997. V. 33. № 1. P. 5.
Brunia C.H., van Boxtel G.J.M. Anticipatory attention to verbal and non-verbal stimuli is reflected in a modality-specific SPN // Exp. Brain Res. 2004. V. 156. № 2. P. 231.
Синицын С.В. Отражение в связанных с событием потенциалах процессов рабочей памяти у детей младшего школьного возраста // Физиология человека. 2008. Т. 34. № 2. С. 128. Sinitsyn S.V. Working memory in young schollchildren: A study using event-related potentials // Human Physiology. 2008. V. 34. № 2. P. 250.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека