

Доклады Российской академии наук. Науки о Земле, 2020, T. 493, № 1, стр. 89-93
ВЛИЯНИЕ ПРОГРЕССИРУЮЩЕГО РАСПАДА ДРЕВОСТОЯ НА УГЛЕРОДНЫЙ ОБМЕН ЕЛОВЫХ ЛЕСОВ
Д. В. Карелин 1, 2, *, Д. Г. Замолодчиков 2, А. В. Шилкин 3, А. С. Куманяев 4, С. Ю. Попов 4, Н. О. Тельнова 1, М. Л. Гитарский 5
1 Институт географии Российской академии наук
Москва, Россия
2 Центр по проблемам экологии и продуктивности лесов Российской академии наук
Москва, Россия
3 НПО “Тайфун” Росгидромета Российской Федерации
Обнинск, Калужская область, Россия
4 Московский государственный университет
имени М.В. Ломоносова
Москва, Россия
5 Филиал "Угреша" Университета “Дубна”
Дзержинский, Московская область, Россия
* E-mail: dkarelin7@gmail.com
Поступила в редакцию 08.04.2020
После доработки 14.04.2020
Принята к публикации 20.04.2020
Аннотация
Многолетнее воздействие распада древостоя ели на углеродный баланс изучали в таежных лесах Валдая (Новгородская обл.). Оценивали эмиссию CO2 из почвы, дыхание древесного дебриса, валовое дыхание, нетто-потоки CO2 и водяного пара. Гибель древостоя еловых насаждений в результате воздействия ксилофагов и сапротрофов, а также ветровалов не приводит к снижению удельного валового дыхания на таких участках. Это связано с тем, что сокращение выделения CO2 от дыхания живых деревьев компенсируется приростом эмиссии от валежа, а также резким (3–3.5 кратным) и продолжительным (не менее 6 лет) увеличением дыхания почвы под сухостоем елей (“hotspots”). Вклад в поток валового дыхания с единицы площади погибшего елового древостоя из почвы под сухостоем (14.4%) превышает вклад разложения крупного древесного дебриса (9%). За период наблюдений отмечено трехкратное снижение годового нетто-стока CO2 с –300 до –95 гС м–2 год–1, которое сопровождается снижением эвапотранспирации (0.0116 г H2O м–2 сек–1; май–октябрь 2018) по сравнению с аналогичным периодом 2010 г. (0.0142 г H2O м–2 сек–1). Наблюдаемое снижение нетто-поглощения связано как с ростом валового дыхания в результате увеличения площадей с распавшимся древостоем, так и со снижением первичной продукции елей. Дальнейшее увеличение площади распада древостоя до 27% превращает нетто-баланс еловой тайги в источник СО2 для атмосферы, что можно учитывать при дистанционных прогнозах.
В современной биосфере, несмотря на растущее антропогенное влияние, связанное с землепользованием, леса остаются одним из важнейших регуляторов газового состава атмосферы и альбедо, что имеет прямое влияние на климат [1]. Важнейшей структурно-функциональной компонентой лесов, в том числе бореальных, фитомасса которых составляет 14.9% от лесов мира [2], является древостой. Он определяет характерные скорости продукции и разложения органических веществ на лесных территориях, а также содержание органического углерода в почвах лесной экосистемы. Поэтому вопросы, связанные с климатическим (в том числе антропогенно обусловленным) или прямым антропогенным влиянием на С-баланс лесных экосистем через состояние их древостоя, относятся к важнейшим в области глобальной экологии.
В основу исследования положен ряд многолетних (с 2009 г. по н.в.) наблюдений за таежными экосистемами полигона Валдайского филиала Государственного гидрологического института Росгидромета (Новгородская обл., 57°57′43.0″ с.ш., 33°20′19.4″ в.д., 225.8 м н.у.м.). Подробнее изучаемые экосистемы и методы исследований описаны в коллективной монографии [3]. Преобладают перестойные высокобонитетные еловые леса на моренных отложениях с присутствием сосны и березы при максимальной высоте древостоя до 40 м. Для исследуемого полигона характерны массовые ветровалы сухостоя елей, погибших в 2010–2013 гг. в результате атак ксилофагов (в частности, Ips typographus L) и грибных гнилей. Это усыхание относится к последней общеевропейской вспышке массовой гибели елей [4]. Наиболее существенное ослабление древостоя произошло после аномальной летней засухи 2010 г., которую часто рассматривают как проявление глобального потепления климата [5, 6]. За 2009–2016 гг. запас крупного древесного дебриса на полигоне увеличился втрое, тогда как запас живой древесины снизился от 572 до 312 м3 га–1 [3].
Было оценено влияние прогрессирующего распада древесного яруса на основные показатели С-цикла таежного леса, в частности, почвенную эмиссию, валовое дыхание и нетто-потоки СО2. В качестве основы для их площадной оценки использовали зону отклика (т.н. “футпринт”) автоматической пульсационной установки (АПУ) для измерения потоков СО2 и паров воды. При высоте размещения оборудования 42 м над поверхностью почвы и 5 м над пологом леса максимальный радиус “футпринта” оценен как 500 м, при площади 0.785 км2 [3, 7]. На постоянных пробных площадях и трансектах измеряли запасы основных резервуаров древесины и надземной фитомассы, а также потоки СО2 от крупного древесного дебриса (валежа и сухостоя), а также из почвы.
За период наблюдений увеличилась как общая доля участков с ослабленным или поврежденным древостоем ели (с 5 до 30%), так и средняя на единицу площади эмиссия СО2 из почвы на участках с распадом древостоя: на 16.5% в 2014 г. и на 19% в 2017 г. Дополнительная эмиссия связана с повышенными выбросами углекислого газа из почвы вокруг сухостойных елей на расстоянии не более 0.5 м от стволов [8, 9]. Этот эффект обнаруживается через 1–2 года после гибели дерева и поддерживается на постоянном уровне не менее 6 лет. Степень усиления эмиссии СО2 рядом с сухостоем, по сравнению с межстволовыми участками, составляет высокую и достаточно постоянную величину (3–3.5 раза), которая мало меняется от года к году. Механизм явления остается до конца не ясен. Наиболее вероятно это связано с повышенной активностью грибов-ксилотрофов и с облегченным транспортом газа в прикомлевой зоне почвы под сухостоем. Вторым дополнительным слагаемым эмиссии СО2 на участках распада древостоя является эмиссия от разложения растущего запаса валежа. В общий поток валового дыхания на этих участках может входить дыхание сохранившихся живых деревьев. Для сравнения уровней валового дыхания (ВД) леса, не затронутого усыханием, и на участках с полным распадом древостоя были рассчитаны величины основных слагаемых ВД, включая эмиссию из почвы (с подстилкой) на межстволовых участках, в пристволовых зонах сухостоя, от крупного древесного дебриса и живых деревьев. На участках распада древостоя наибольший вклад в суммарный поток 815 ± 24.6 гC м–2 год–1 предоставлен почвой межстволовых участков (76.6%), на втором месте находится вклад зон почвы под сухостоем (14.4%), на третьем – эмиссия от валежа (9%). В незатронутом усыханием ельнике на втором месте по участию в ВД после эмиссии из почвы находится вклад дыхания живого древостоя. Согласно блоковой модели, суммарный поток ВД в здоровом лесу меняется в пределах 922.1–970.7 гC м–2 год–1. Отметим, что на участках распада древостоя вклад эмиссии от разложения дебриса существенно зависит от его количества и плотности. Так, по нашим ранним оценкам ВД на модельных участках распада древостоя в 2013 г. составило 1480 гC м–2 год–1 [3]. Поэтому полученное различие между ВД участка распада и незатронутого усыханием ельника нельзя считать существенным. Убыль дыхания автотрофов при усыхании древостоя компенсируется увеличением дыхания гетеротрофов на сходную величину.
Для С-баланса леса в целом важно, что участки распада длительно функционируют как источник углерода, занимая при этом значительную территорию. Оценить их вклад для большой площади можно с помощью пульсационного метода. Как показывает сравнение оценок годового нетто-баланса С в период наиболее активной деградации елового древостоя, в 2010–2011 гг. баланс в экосистеме составил –300 гC-СО2 м–2 год–1 (сток С из атмосферы), тогда как в 2018 г. сток снизился до –95 гC-СО2 м–2 год–1. Это произошло не только в результате роста валового дыхания, но и снижения первичной продукции ослабленного елового древостоя, что подтверждается значимым падением эвапотранспирации за май–октябрь в сравниваемые годы; 0.0142 ± 0.0003 гH2O м–2 сек–1 (n = 6475) в 2010 г. и 0.0116 ± 0.0002 гH2O м–2 сек–1 (n = 7677) в 2018 г. (отличия значимы: медианный тест, p = 0.027).
Сопоставление средних величин нетто-потоков С по секторам направлений ветра по данным АПУ с мая по октябрь 2010 и 2018 гг. с присутствием в этих секторах различных биотопов показало, что наибольшая положительная корреляция отмечается между источником С и площадями, занятыми усыхающим еловым древостоем и безлесными участками в пределах от 30–40 до 300–350 м от измерительной установки (рис. 1, 2). При этом в 2018 г. эта связь становится значимой (rp = = +0.77, p = 0.026).
Рис. 1.
Средний нетто-поток углерода (мг C-CO2 м–2 сек–1, N = 6475) с мая по октябрь 2010 (а), и (б) доля площади, занятая усыхающим еловым древостоем и безлесными участками по направлениям ветра (1 – север, 2 – северо-восток, … 8 – северо-запад) в зоне от 40 до 300 м от автоматической пульсационной установки. Отрицательные величины нетто-потока обозначают сток С-СО2 из атмосферы, положительные – источник.
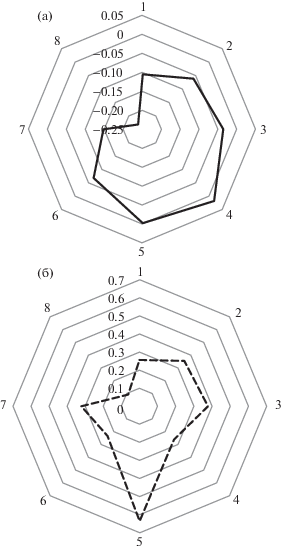
Рис. 2.
Средний нетто-поток углерода (мг C-CO2 м–2 сек–1, N = 7677) с мая по октябрь 2018 (а), и (б) доля площади, занятая усыхающим еловым древостоем и безлесными участками в зоне от 30 до 350 м от автоматической пульсационной установки. Отрицательные величины нетто-потока обозначают сток С-СО2 из атмосферы, положительные – источник.
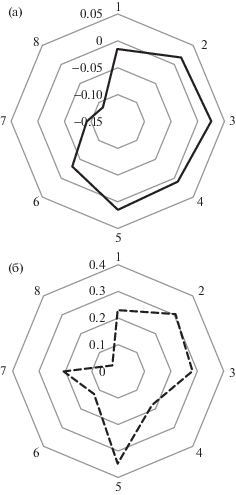
Таким образом, прогрессирующее ослабление и гибель елового древостоя, связанное с современным потеплением, приводит к трехкратному снижению стока С на значительных территориях, несмотря на сопровождающие этот процесс восстановительные сукцессии. Снижение стока связано с сохранением величины валового дыхания на фоне снижения продукции. Расчеты показывают, что увеличение площади полного распада древостоя до 27% превращает территорию бореального леса в источник СО2 для атмосферы, что может быть использовано при дистанционных оценках влияния климатических или антропогенных воздействий на С-баланс лесов.
Список литературы
Bonan G.B. // Science. 2008. V. 320. P. 1444–1449.
Pan Y., Birdsey R.A., Phillips O., Jackson R.B. // Ann. Rev. Ecol. Evolution and Systematics. 2013. V. 44. P. 593–622.
Мониторинг потоков парниковых газов в природных экосистемах / Ред. Д.Г. Замолодчиков, Д.В. Карелин, М.Л. Гитарский, В.Г. Блинов. Саратов: Амирит. 2017. 279 с.
Štursová M., Šnajdr J., Cajthaml T., Bárta J., Šantrůčková H., Baldrian P. // ISME Journal. 2014. V. 8. P.1920–1931.
Данилов-Даниэлян В.И. // Экология и жизнь. 2010. № 10. С. 20–27.
Мохов И.И., Семенов В.А. // Метеорология и гидрология. 2016. № 2. С. 16–28.
Замолодчиков Д.Г., Гитарский М.Л., Шилкин А.В., Марунич А.С., Карелин Д.В., Блинов В.Г., Иващен-ко А.И. // Фундаментальная и прикладная климатология. 2017. № 1. С. 54–68.
Карелин Д.В., Замолодчиков Д.Г., Исаев А.С. // ДАН. 2017. Т. 475. № 4. С. 473–476.
Карелин Д.В., Почикалов А.В., Замолодчиков Д.Г. // Известия РАН. Cер географическая. 2017. № 2. С. 60–68.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о Земле