

Доклады Российской академии наук. Науки о Земле, 2020, T. 494, № 1, стр. 5-8
Новая находка докодонта (mammaliaformes, docodonta) в нижнем мелу Западной Сибири
Академик РАН А. В. Лопатин 1, 2, *, А. О. Аверьянов 3, 4, И. Т. Кузьмин 5, Е. А. Бойцова 5, П. Г. Сабуров 5, С. В. Иванцов 6, П. П. Скучас 5
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
2 Московский государственный университет
имени М.В. Ломоносова
Москва, Россия
3 Зоологический институт Российской академии наук
Санкт-Петербург, Россия
4 Казанский (Приволжский) федеральный университет
Казань, Россия
5 Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
6 Томский государственный университет
Томск, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 20.12.2019
После доработки 30.06.2020
Принята к публикации 01.07.2020
Аннотация
Описана первая находка докодонта в местонахождении Большой Кемчуг 3 (илекская свита, нижний мел) в Красноярском крае – нижний моляр Sibirotherium sp. Sibirotherium rossicum, один из позднейших докодонтов в геологической летописи, известен из илекской свиты местонахождения Шестаково 1 (Кемеровская область). Ранее в местонахождениях илекской свиты в Красноярском крае (Большой Терехтюль 2 и Большой Илек) находили только неопределимые остатки докодонтов.
Докодонты – разнообразная группа мезозойских маммалиаформов (архаичных млекопитающих, неродственных современным таксонам), включавшая наземные, роющие, древесные и полуводные экоморфы [1–4]. Для докодонтов было характерно псевдотрибосфеническое строение коренных зубов (с крупным внутренним выступом на верхних зубах и противолежащим ему углублением на нижних), значимое для эффективной обработки пищи при жевании. Благодаря этому они стали одной из доминантных групп в сообществах мелких позвоночных средней – поздней юры Северного полушария.
В России известен комплекс позвоночных из среднеюрских отложений Красноярского края (местонахождение Березовский карьер), в котором докодонты были весьма разнообразны и доминировали среди маммалиаформов [5]. Также в России известны самые поздние докодонты в геологической летописи: Sibirotherium из раннего мела Кемеровской области (местонахождение Шестаково 1) и Khorotherium из раннего мела Якутии (местонахождение Тээтэ) [6–8].
В последние годы также были сделаны находки неопределимых докодонтов в отложениях илекской свиты (нижний мел, баррем–апт) в Красноярском крае (местонахождения Большой Терехтюль 2 и Большой Илек) [9, 10]. В данном сообщении приводится информация о другой новой находке докодонта в России – нижнем коренном зубе из местонахождения Большой Кемчуг 3 в Красноярском крае. Данное местонахождение также приурочено к илекской свите и содержит богатый комплекс млекопитающих [11], однако докодонты здесь ранее не были обнаружены.
Описанный материал хранится в коллекции Лаборатории континентальных экосистем мезозоя и кайнозоя (ЛКЭМК) Томского государственного университета (ТГУ) в Томске. Исследованный материал по Sibirotherium из местонахождения Шестаково 1 находится в коллекции Палеонтологического музея Томского государственного университета (ПМ ТГУ).
Левый нижний моляр, экз. ЛКЭМК 005/127 (рис. 1), был найден в 2015 г. в ходе промывки костеносной породы на местонахождении Большой Кемчуг 3 на правом берегу р. Большой Кемчуг в Емельяновском районе Красноярского края (56°31′38″ с.ш., 91°48′49″ в.д.). Разрез нижнемеловых отложений илекской свиты на данном местонахождении состоит из желтоватых и зеленовато-серых плохо литифицированных полимиктовых песчаников с включением аргиллитовых галек. В нижней части разреза встречаются обугленные стволы и ветки растений. В верхней части разреза имеется серия линз мощностью до 2 м с остатками мелких позвоночных. В 2015 г. было промыто 698 кг породы из этих линз и обнаружено несколько сотен определимых остатков позвоночных, включая млекопитающих.
Рис. 1.
Sibirotherium sp., экз. ЛКЭМК 005/127, левый нижний моляр, вид с окклюзиальной стороны (стереопара), сзади, с лингвальной стороны, спереди и с лабиальной стороны; буквы a–d и bb обозначают бугорки; Большой Кемчуг 3, Красноярский край, Россия; илекская свита, нижний мел.
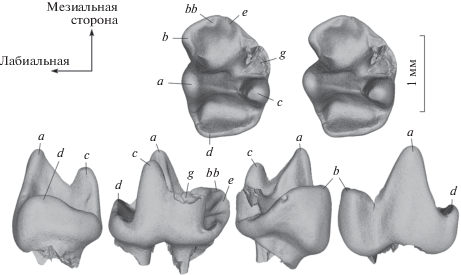
На экз. ЛКЭМК 005/127 главный бугорок a занимает своим основанием больше половины лабиального края коронки. Его передний и задний края в виде приостренных, слегка вогнутых ребер. Два близко расположенных лингвальных гребня соединяют вершину бугорка a с вершинами лингвальных бугорков c и g (последняя обломана). Гребень a-c заметно более острый и высокий по сравнению с гребнем a-g. Наиболее низкая часть гребня a-c расположена существенно выше наиболее низкой части гребня a-g. Бугорок c заметно ниже бугорка a и его переднее и заднее ребра не такие острые. Основание бугорка g немного длиннее мезиодистально и короче лабиолингвально основания бугорка c. Оба бугорка соединены низким гребнем c-g, выраженным только в их основании. Бугорки c и g разделены неглубокой вертикальной бороздкой с лингвальной стороны. Лингвальный цингулид в основании бугорков c и g практически не выражен.
Спереди и сзади от центрального углубления коронки, образованного бугорками a, c и g и гребнями a-c и a-g, расположены два обширных и глубоких бассейна – псевдоталонидный спереди и талонидный сзади. Талонид немного шире псевдоталонида лабиолингвально, но существенно короче мезиодистально. Псевдоталонидный бассейн ограничен высокими бугорком b с лабиальной стороны, плохо выраженным бугорками bb и e спереди и невысоким гребнем g-e с лингвальной стороны. Лингвальнее этого гребня расположена короткая, но глубокая поперечная бороздка, ограниченная отчетливым гребнем спереди. Бугорки bb и e расположены на гребне, полого спускающемся с бугорка b. Бугорок bb обозначен небольшой фасеткой стирания, бугорок e – коротким гребешком, поднимающимся к нему с дистальной стороны. Дно псевдоталонидного бассейна расположено несколько выше дна талонидного бассейна. Наиболее глубокая часть псевдоталонида находится в основании бугорка a, между гребнями a-b и a-g. Талонидный бассейн ограничен гребнем a-d с лабиальной стороны и почти поперечно расположенным гребневидным бугорком d сзади. Между этим гребнем и основанием бугорка c имеется узкая бороздка, оставляющая талонидный бассейн открытым с лингвальной стороны. Наиболее глубокая часть талонида расположена у его лингвального края, в основании бугорка a. Наиболее низкая часть гребня a-b расположена существенно выше, чем наиболее низкая часть гребня a-d.
Коронка зуба мало стерта. Наиболее широкая фасетка стирания наблюдается вдоль гребня a-b.
Оба корня зуба примерно равны по ширине. Передний корень немного короче мезиодистально заднего корня. На мезиальной стороне переднего корня имеется обширная впадина. Корни связаны между собой небольшим продольным гребнем в основании коронки.
Экз. ЛКЭМК 005/127 по длине коронки (1.6 мм) соответствует задним молярам (m4–m5) докодонта Sibirotherium rossicum Maschenko et al., 2003 из местонахождения Шестаково 1 (илекская свита, Кемеровская обл. [6, 7]), однако по ширине коронки (1.2 мм) несколько превосходит их (рис. 2). Как отмечалось ранее [7], экз. ПМ ТГУ 120/9-34 заметно крупнее других экземпляров сибиротерия из Шестаково 1. Также на данном экземпляре m1 более широкий, с лучше развитым талонидом по сравнению с m1 на голотипе S. rossicum. Короткий и широкий моляр из Большого Кемчуга 3 больше соответствует экз. ПМ ТГУ 120/9-34, чем другим молярам сибиротерия из Шестаково 1. Отличия между двумя экземплярами заключаются в практически не развитом лингвальном цингулиде на экз. ЛКЭМК 005/127. Степень развития лингвального цингулида нижних моляров, однако, сильно варьирует у сибиротерия.
Рис. 2.
Длина (L) и ширина (W) коронки нижних моляров (m1, m2, m3, m4, m5) Sibirotherium rossicum из местонахождения Шестаково 1 (Кемеровская область, илекская свита, нижний мел) и Sibirotherium sp. (mx) из местонахождения Большой Кемчуг 3 (Красноярский край, илекская свита, нижний мел).
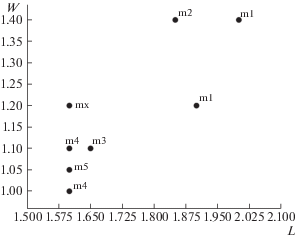
У сибиротерия соотношение бугорков c и g варьирует в зависимости от положения моляра в зубном ряду. Бугорок g заметно меньше, чем бугорок c на m1 и m3, а на m2 эти бугорки одинаковой высоты. На экз. ЛКЭМК 005/127 бугорок g обломан, но, судя по его основанию, он мог быть такой же высоты, как и бугорок c. Если это так, экз. ЛКЭМК 005/127 может быть m2. Кроме того, только у m2 пока известен неполный гребень a-g.
Пока не вполне ясен вопрос о числе видов сибиротерия. Крупные размеры экз. ПМ ТГУ 120/9-34 объяснялись индивидуальной или половой изменчивостью [7]. Однако нельзя исключить существование в одной палеоэкосистеме двух близких видов, различающихся по размерам. Экз. ЛКЭМК 005/127 из Красноярского края ближе по морфологии к крупному экз. ПМ ТГУ 120/9-34 из Шестаково 1 и может быть определен как Sibirotherium sp.
Список литературы
Martin T. // Zool. J. Linnean Soc. 2005. V. 145. № 2. P. 219–248.
Ji Q., Luo Z.-X., Yuan C.-X., Tabrum A.R. // Science. 2006. V. 311. № 5764. P. 1123–1127.
Luo Z.-X., Meng Q.-J., Ji Q. et al. // Science. 2015. V. 347. № 6223. P. 760–764.
Meng Q.-J., Ji Q., Zhang Y.-G. et al. // Science. 2015. V. 347. № 6223. P. 764–768.
Averianov A.O., Lopatin A.V., Krasnolutskii S.A., Ivantsov S.V. // Proceedings of the Zoological Institute of the Russian Academy of Sciences. 2010. V. 314. № 2. P. 121–148.
Maschenko E.N., Lopatin A.V., Voronkevich A.V. // Russ. J. Theriol. 2003. V. 1. № 2. P. 75–81.
Лопатин А.В., Аверьянов А.О., Мащенко Е.Н., Лещинский С.В. // Палеонтол. журн. 2009. № 4. С. 92–100.
Averianov A.O., Martin T., Lopatin A.V. et al. // PLoS ONE. 2018. V. 13. № 7: e0199983.
Averianov A.O., Lopatin A.V. // Cretaceous Research. 2015. V. 54. P. 145–153.
Лопатин А.В., Аверьянов А.О., Иванцов С.В. // ДАН. 2019. 487. № 4. С. 414–417.
Averianov A.O., Skutschas P.P., Lopatin A.V. et al. // Russ. J. Theriol. 2003. V. 4. № 1. P. 1–12.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о Земле