

Доклады Российской академии наук. Науки о Земле, 2023, T. 508, № 1, стр. 88-92
Arginbaatar (Multituberculata, Mammalia) из нового раннемелового местонахождения Шавар-Ово в Монголии
Академик РАН А. В. Лопатин 1, *
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 08.09.2022
После доработки 09.09.2022
Принята к публикации 15.09.2022
- EDN: GESKMX
- DOI: 10.31857/S2686739722601831
Аннотация
Из нового раннемелового местонахождения Шавар-Ово в Монголии (аймак Уверхангай, сомон Гучин-Ус) описан изолированный верхний предкоренной зуб P4 мультитуберкулята, демонстрирующий строение и размеры, характерные для Arginbaatar dmitrievae Trofimov, 1980 (Arginbaataridae). Это первая находка Arginbaatar (и в целом раннемеловых мультитуберкулят Монголии) за пределами урочища Ховур.
В Монголии раннемеловые млекопитающие известны из пяти местонахождений: Ховур, Зун-Ховур, Шалан-Ихэр, Оши (Оши-Нуру) и Хамрын-Ус. Первые четыре расположены в аймаке Уверхангай в центральной части Монголии, а пятое находится в аймаке Дорноговь на юго-востоке страны ([1], рис. 1). Во всех этих местонахождениях найдены эутриконодонты рода Gobiconodon [1–7], тогда как представители других групп млекопитающих, включая мультитуберкулят, указаны только для двух наиболее богатых местонахождений ховурского комплекса – Ховур (Höövör, Khovoor) и Зун-Ховур (Zuun-Höövör, Zun-Khovoor) [8–12]. Они расположены в 1.5 км друг от друга в урочище Ховур Гучинской впадины Северной Гоби.
Описанные ранее [8, 9] материалы по мультитуберкулятам из местонахождения Ховур были недавно нами ревизованы и дополнены [12]. Кроме того, многочисленные зубные и челюстные остатки мультитуберкулят Arginbaatar dmitrievae Trofimov, 1980 (= Monobaatar mimicus Kielan-Jaworowska et al., 1987) и Eobaatar magnus Kielan-Jaworowska et al., 1987 (а также следы зубов мультитуберкулят на кости эутриконодонта Gobiconodon borissiaki Trofimov, 1978) были найдены нами в местонахождении Зун-Ховур [6, 7, 10, 11].
Обнаруженный в 2022 г. Южно-Гобийским отрядом Совместной российско-монгольской палеонтологической экспедиции (ЮГО СРМПЭ) изолированный зуб мультитуберкулята Arginbaatar dmitrievae происходит из нового местонахождения Шавар-Ово (Shavar-Ovoo) в Гучинской впадине (сомон Гучин-Ус, аймак Уверхангай; 45°22′27″ с.ш., 102°35′51″ в.д.) (рис. 1). Это местонахождение было открыто нами (ЮГО СРМПЭ: А.В. Лопатин, А.А. Карху, Палеонтологический институт им. А.А. Борисяка РАН; Д. Бадамгарав, Институт палеонтологии Монгольской академии наук) еще в 2012 г., однако до 2022 г. в нем были найдены только остатки динозавров и неопределимые фрагменты костей млекопитающих.
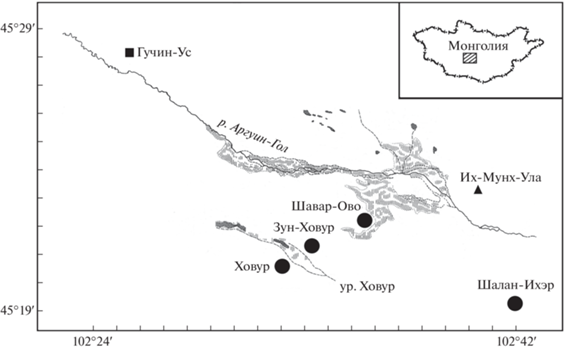
В местонахождении Шавар-Ово, в останце у южного борта долины, разрез нижнемеловой (апт-альбской) хухтыкской свиты представлен следующими отложениями (снизу вверх): толща светло-серых песков, перекрытая слоем коричневато-серых плотных песчаников мощностью до 1 м, содержащих фрагменты костей динозавров; толща косослоистых желтовато-серых глинистых песков (мощность до 6 м; в верхней части встречаются фрагменты панцирей черепах), бронируемая слоем коричневато-серых плотных песчаников мощностью до 0.3 м; пестроцветная толща песчанистых глин (мощность до 5 м). Зуб мультитуберкулята Arginbaatar dmitrievae получен в результате промывки около 1.5 т породы из верхней части толщи косослоистых желтовато-серых глинистых песков.
Описываемый в настоящей статье зуб представляет собой первую находку Arginbaatar (и в целом раннемеловых мультитуберкулят Монголии) за пределами урочища Ховур. Материал хранится в коллекции Палеонтологического института им. А.А. Борисяка РАН (ПИН), Москва. Изображения получены с помощью сканирующего электронного микроскопа Tescan Vega II XMU в ПИН. Номенклатура бугорков зубов мультитуберкулят по [13].
Изолированный левый верхний премоляр P4 (экз. ПИН, № 5875/1) Arginbaatar dmitrievae (Arginbaataridae, Plagiaulacoidea) имеет два длинных корня и сравнительно низкую коронку (рис. 2). Окклюзиальная поверхность субпрямоугольной формы, с небольшим постеролингвальным выступом. Бугорковая формула – 3B:4L (три лабиальных (буккальных) и четыре лингвальных бугорка). Бугорки конические, слегка сжаты поперечно.
Рис. 2.
Arginbaatar dmitrievae Trofimov, 1980, экз. ПИН, № 5875/1, левый P4: а – с окклюзиальной стороны, б – обозначения бугорков, в – с лабиальной стороны, г – с лингвальной стороны, д – положение в зубном ряду (см. [12]), с окклюзиальной стороны; местонахождение Шавар-Ово, Монголия; хухтыкская свита, нижний мел.
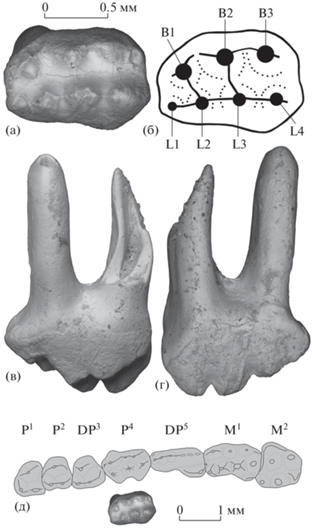
В лабиальном ряду средний бугорок B2 крупнее и выше других бугорков, задний бугорок B3 намного выше и массивнее переднего бугорка B1. У заднего основания бугорка B3 имеется небольшой дистальный выступ. По лабиальному краю окклюзиальной стороны бугорок B2 связан с бугорками B1 и B3 передним и задним продольными гребнями – соответственно, B1–B2 и B2–B3, которые ограничивают поперечные долинки с лабиальной стороны.
В лингвальном ряду бугорки постепенно увеличиваются в заднем направлении по высоте и массивности, наиболее крупный бугорок L4 в основании выступает дистально. Бугорок L1 очень мелкий, расположен в антеролингвальном углу окклюзиальной поверхности, прижат к основанию бугорка L2 и немного удален от основания бугорка B1. Бугорки L2 и L3 имеют по три коротких гребневидных ребра, которые встречаются с супротивными ребрами соседних бугорков в области межбугорковых седловин и попарно с ними формируют гребни L1–L2, B1–L2, L2–L3, B2–L3 и L3–L4, связывающие соответствующие бугорки. Продольные гребни L1–L2, L2–L3 и L3–L4 ограничивают поперечные долинки с лингвальной стороны.
Срединная продольная долинка между лабиальным и лингвальным рядами бугорков мелкая в области бугорков B1/L1–L2 и B2/L3, но более глубокая на уровне бугорков B2/L2–L3 и B3/L3–L4. В двух местах она частично перегорожена низкими косыми гребнями B1–L2 и B2–L3, связывающими супротивные бугорки.
Корни широко расставленные, продольно сжатые.
Размеры коронки экз. ПИН, № 5875/1 (в мм): наибольшая длина – 1.28, наибольшая ширина – 0.88.
Бугорки умеренно стерты апикально, фасетки стирания лабиальных бугорков треугольной формы. Апикальные фасетки стирания лингвальных бугорков звездчатые, при этом фасетки стирания распространяются на лингвальную поверхность коронки и объединяются друг с другом в основании коронки (рис. 2г). На материалах из местонахождения Ховур показано, что при дальнейшем стирании P4 формировалась единая лингвальная фасетка ([12], рис. 25а), которая затем сильно расширялась и уплощалась, и в лингвальной части окклюзиальной поверхности коронки формировался режущий край, а на наиболее поздних стадиях все лингвальные бугорки выравнивались и в итоге исчезали ([12], рис. 18а, 22b, 22c, 22f). Похожим образом происходило стирание постеролингвальной части DP5 ([12], рис. 25k, 25l).
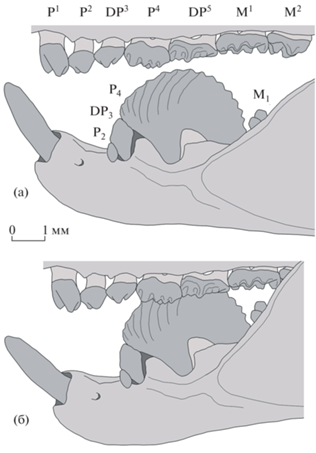
Следует полагать, что у Arginbaatar dmitrievae P4 и DP5 при жевании взаимодействовали с последним нижним премоляром P4 – большим узким зубом с многочисленными эмалевыми гребнями и отчетливыми зубцами на дугообразно выгнутой лезвиеподобной кромке (рис. 3). Видимо, такое строение P4 позволяло этим мультитуберкулятам эффективно разрезать и измельчать плоды и семена растений. В процессе индивидуального развития, чтобы компенсировать стирание зубчатого рельефа, коронка P4 постепенно поднималась на заднем корне и наклонялась вперед, таким образом все больше проворачиваясь задней стороной кверху; это уникальное явление называется “мезиовентральной ротацией” [9, 12]. Благодаря этой специализации в течение почти всей жизни у аргинбаатара на P4 функционировала острая пильчатая кромка, а не сточенная плоская площадка, что обеспечивало более эффективное режущее воздействие на пищевые объекты. При этом окклюдировавшие с P4 задние верхние предкоренные зубы – P4 и DP5 – интенсивно стирались с лингвальной стороны, в результате чего по вентральному краю субвертикальных лингвальных фасеток формировались острые кромки, усиливающие режущую функцию.
Рис. 3.
Arginbaatar dmitrievae Trofimov, 1980, реконструкция верхнего и нижнего зубных рядов, с лабиальной стороны: а – строение (см. [12]), б – окклюзия.
Для представителей семейства Paulchoffatiidae, также имевших увеличенный лезвиеподобный P4, показаны наличие двух боковых фасеток стирания на лингвальной стороне коронки P4 и тенденция к их слиянию при дальнейшей абразии лингвальных бугорков и формировании режущего края в лингвальной части окклюзиальной поверхности [14], ([15], рис. 5). Лабиальная фасетка стирания P4 окклюдировала с постеролингвальной частью P4, а лингвальные склоны лабиальных бугорков P4 – с лабиальными сторонами бугорков лингвального ряда на P4 ([15], рис. 7b). Видимо, у Arginbaatar окклюзия осуществлялась сходным образом, но с учетом отсутствия лабиальных бугорков в основании P4 и в целом более мощного развития этого зуба.
Подобно другим мультитуберкулятам [13–17], Arginbaatar при жевании использовал дистально направленные (палинальные) движения. Большой нижний премоляр P4, взаимодействуя лабиальной частью своего лезвия с лингвальными бугорками P4 и DP5, разрезал пищевые объекты, которые латерально фиксировались лабиальными бугорками этих двух верхних премоляров. Когда при жевании нижняя челюсть сдвигалась назад, лезвие P4 смещалось от P4 к постеролингвальной части DP5.
В коллекциях из местонахождений Ховур и Зун-Ховур Arginbaatar dmitrievae представлен наибольшим числом остатков среди Multituberculata [12]; видимо, этот сравнительно мелкий вид многобугорчатых преобладал среди растительноядных млекопитающих ховурского комплекса.
Список литературы
Лопатин А.В. Раннемеловые млекопитающие из местонахождений Хамрын-Ус и Шалан-Ихэр в Монголии // ДАН. 2017. Т. 477. № 1. С. 113–117.
Трофимов Б.А. Первые триконодонты (Mammalia, Triconodonta) из Монголии // ДАН СССР. 1978. Т. 243. № 1. С. 213–216.
Kielan-Jaworowska Z., Dashzeveg D. Early Cretaceous amphilestid (‘triconodont’) mammals from Mongolia // Acta Palaeontol. Polon. 1998. V. 43. № 3. P. 413–438.
Rougier G.W., Novacek M.J., McKenna M.C., Wible J.R. Gobiconodonts from the Early Cretaceous of Oshih (Ashile), Mongolia // Amer. Museum Novit. 2001. № 3348. P. 1–30.
Lopatin A.V., Averianov A.O. Gobiconodon (Mammalia) from the Early Cretaceous of Mongolia and revision of Gobiconodontidae // J. Mammal. Evol. 2015. V. 22. № 1. P. 17–43.
Лопатин А.В. Следы зубов млекопитающих на кости эутриконодонта Gobiconodon borissiaki (Mammalia, Gobiconodontidae) из нижнего мела Монголии // Докл. РАН. Науки о Земле. 2022. Т. 502. № 1. С. 22–25.
Лопатин А.В. Прямое свидетельство смены моляриформных зубов у Gobiconodon borissiaki (Gobiconodontidae, Mammalia) из раннего мела Монголии // Докл. РАН. Науки о жизни. 2022. Т. 504. С. 235–239.
Трофимов Б.А. Multituberculata и Symmetrodonta из нижнего мела Монголии // ДАН СССР. 1980. Т. 251. № 1. С. 209–212.
Kielan-Jaworowska Z., Dashzeveg D., Trofimov B.A. Early Cretaceous multituberculates from Mongolia and a comparison with Late Jurassic forms // Acta Palaeontol. Polon. 1987. V. 32. № 1–2. P. 3–47.
Лопатин А.В. Новые находки раннемеловых млекопитающих в Монголии // ДАН. 2013. Т. 449. № 4. С. 491–493.
Лопатин А.В., Бадамгарав Д. Новое местонахождение раннемеловых млекопитающих Зун-Ховур (Уверхангай, Монголия) // ДАН. 2013. Т. 453. № 4. С. 413–415.
Lopatin A.V., Averianov A.O. Multituberculata from the Early Cretaceous of Mongolia // Paleontol. J. 2021. V. 55. № 11. P. 1275–1317.
Kielan-Jaworowska Z., Cifelli R.L., Luo Z.-X. Mammals from the age of dinosaurs: origins, evolution, and structure. New York, Columbia University Press, 2004. 630 p.
Hahn G., Hahn R. The dentition of Plagiaulacida (Multituberculata, Late Jurassic to Early Cretaceous) // Geologica et Palaeontologica. 2004. V. 38. P. 119–156.
Lazzari V., Schultz J.A., Tafforeau P., Martin T. Occlusal pattern in paulchoffatiid multituberculates and the evolution of cusp morphology in mammaliamorphs with rodent-like dentitions // J. Mammal. Evol. 2010. V. 17. P. 177–192.
Gingerich P. Patterns of evolution in the mammalian fossil record // Patterns of evolution / Ed. A. Hallam. Amsterdam: Elsevier Scientific Publishing Co., 1977. P. 469–500.
Gambaryan P.P., Kielan-Jaworowska Z. Masticatory musculature of Asian taeniolabidoid multituberculate mammals // Acta Palaeontol. Polon. 1995. V. 40. № 1. P. 45–108.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о Земле