

Доклады Российской академии наук. Науки о жизни, 2020, T. 492, № 1, стр. 251-254
СВЯЗАН ЛИ ЭФФЕКТ ЧИТТИ С ТИПАМИ ПОПУЛЯЦИОННОЙ ДИНАМИКИ?
В. Д. Якушов 1, Б. И. Шефтель 2, *
1 Пензенский государственный университет
Пенза, Россия
2 Институт проблем экологии и эволюции
им. А.Н. Северцова Российской академии наук
Москва, Россия
* E-mail: borissheftel@yahoo.com
Поступила в редакцию 07.02.20
После доработки 20.02.2020
Принята к публикации 20.02.2020
Аннотация
Тридцатилетние исследования популяций восьми видов землероек-бурозубок в Средней Енисейской тайге показали, что в конце XX столетия динамика популяции имела хорошо выраженный циклический характер, а в XXI веке циклические колебания сменились флуктуирующими. Изучение зависимости веса тела зверьков от уровня численности популяции показало, что при циклических колебаниях на пиках численности вес тела зверьков достоверно выше, чем при спадах численности, т.е. соблюдается следствие из гипотезы Читти (эффекта Читти), а при флуктуирующей популяционной динамике подобная закономерность не прослеживается.
В настоящее время описано довольно много различных типов популяционной динамики мелких млекопитающих [1]. Но в данном исследовании нас интересовали только два широко распространенных типа: циклические колебания численности, характерные для стабильных климатических условий, и ациклические, или флуктуации, зависящие от условий среды, в первую очередь климатических. В первом случае лимитирующим фактором выступает переуплотнение популяции при достижении определенного порогового уровня, что ведет к снижению численности. Во втором случае высокая численность соответствует оптимальным условиям среды, а ухудшение условий приводит к снижению численности [2]. При циклических колебаниях на пике численности популяция находится в угнетенном состоянии, а при флуктуациях – наоборот, в оптимальном [3].
Известно, что при резком падении численности популяции или сообщества (группы популяций экологически сходных видов), которое происходит после популяционного пика, уменьшается также вес зверьков. Этот феномен свойственен циклическим популяциям и его часто считают следствием из гипотезы Читти или эффектом Читти [4–6].
Однако в конце XX – начале XXI в. во многих регионах Голарктики циклические колебания численности мелких млекопитающих сменились флуктуационными, ациклическими. Эти изменения связывают в первую очередь с глобальными изменениями климата [7]. Анализ динамики популяций – довольно чувствительный подход, позволяющий оценить последствия глобального изменения климата для биоразнообразия, поскольку в результате таких изменений может произойти как модификация биологии отдельных видов, так и сообщества в целом.
Мы проводили многолетние наблюдения за изменениями численности сообщества землероек-бурозубок на Енисейской экологической станции “Мирное” ИПЭЭ РАН в Средней Енисейской тайге. Сообщество состояло из 8 видов, доминировали обыкновенная бурозубка (Sorex araneus), средняя бурозубка (S. caecutiens), равнозубая бурозубка (S. isodon) и малая бурозубка (S. minutus). Кроме того, отлавливались тундряная бурозубка (S. tundrensis), плоскочерепная бурозубка (S. roboratus), крошечная бурозубка (S. minutissimus) и крупнозубая бурозубка (S. daphaenodon). Исследования велись с 1973 по 1994 гг. и с 2007 по 2019 гг. Землероек отлавливали с помощью ловчих канавок, а там, где вырыть канавки невозможно, ставили заборчики [8, 9]. Для расчетов использованы массы 11117 зверьков в циклический период динамики и 3945 – в флуктуационный период.
Для вычисления периода колебаний численности использовали автокорреляционный анализ [10].
Для оценки влияния стадии цикла на вес особей, входящих в состав сообщества землероек, использовали обобщенные линейные модели (Generalized Linear Models). Это универсальный метод построения регрессионных моделей, позволяющий учитывать взаимодействие между факторами, вид распределения зависимой переменной и предположения о характере регрессионной зависимости.
Анализ проводился средствами компьютерной программы Statistica 10. В качестве зависимой переменной был выбран вес, предикторами являются вид млекопитающего, пол, возраст и стадия цикла. Беременные и кормящие самки не были включены в расчеты. Функция связи – тождественная, к этому выбору мы пришли в ходе анализа диаграмм рассеивания, на которых видна линейная взаимосвязь между переменной отклика и предикторами. Форма распределения ошибок (остатков) стремится к нормальной. Поскольку все предикторы являются категориальными переменными, был выбран план анализа “Главные эффекты” (Main effects ANOVA). Значимость эффектов оценивалась на основе статистики Вальда.
Исследования, проведенные на экологической станции в XX в., показали наличие четырехлетней цикличности в популяционной динамике большинства видов землероек, причем у популяций, обитающих на разных берегах Енисея, наблюдались синхронные циклические колебания [11]. Наблюдения, проведенные в XXI в., обнаружили, что динамика популяций в этот период скорее соответствует флуктуациям, нежели четырехлетней цикличности [12].
Посредством автокорреляционного анализа [10] было обнаружено, что для первого периода наблюдений характерен четкий и статистически значимый период колебаний, равный 4 годам (коэффициент корреляции r = 0.7, p = 0.0009). В XXI в. четкого периода выявить не удалось (рис. 1, 2).
Рис. 1.
Анализ периода колебаний численности сообщества землероек в XX в. “Lag” – величина смещения временных рядов относительно друг друга, “Corr” – коэффициент корреляции, “S.E.” – стандартная ошибка, “р” – уровень значимости.

Рис. 2.
Анализ периода колебаний численности сообщества землероек в XXI в. “Lag” – величина смещения временных рядов относительно друг друга, “Corr” – коэффициент корреляции, “S.E.” – стандартная ошибка, “р” – уровень значимости. В отличие от XX в., четкого периода выявить не удается.
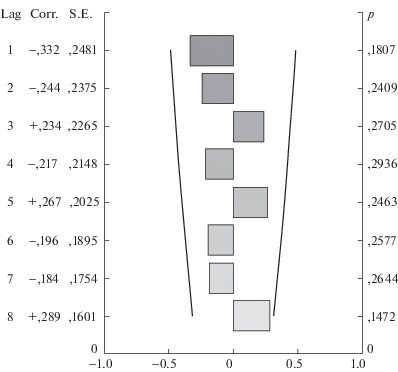
Наибольший коэффициент корреляции соответствует смещению (лагу) в 4 года, что и является искомым значением периода колебаний численности. Более того, прослеживается и восьмилетний период, что говорит о стабильном проявлении четырехлетних колебаний в течение длительного временного промежутка.
При изучении связи между весом зверьков и численностью популяций случайным образом было выбрано 5 пар лет: 2 пары в циклический период динамики и 3 пары лет в период с флуктуирующей динамикой численности. В обеих парах лет от первого ко второму году наблюдалось падение численности во всех популяциях сообщества землероек.
В результате проведенного анализа было выявлено, что в ХХ в. при циклических колебаниях численности вес особей зависел от стадии цикла (коэффициент регрессии равен –0.15 ± 0.002; статистика Вальда = 18; p < 0.01). Зверьки меньше на стадии депрессии в среднем на 0.7 г (7.9 ± 0.03 г в годы высокой численности и 7.2 ± 0.06 г в годы низкой численности) (табл. 1). Стоит заметить, что в рамках использованной модели изменения среднего веса пойманных зверьков оценивались с учетом межвидовых и половозрастных различий.
Таблица 1.
Результаты регрессионного анализа. “Ц” – период, характеризующийся циклическими колебаниями численности, “Фл” – период, характеризующийся нециклическими, флуктуирующими колебаниями
| Коэффициент регрессии | Стандартная ошибка | Статистика Вальда | Р-уровень значимости | |||||
|---|---|---|---|---|---|---|---|---|
| Предиктор | Ц | Фл | Ц | Фл | Ц | Фл | Ц | Фл |
| Вид | 2.44 | 2.5 | 0.06 | 0.2 | 1521 | 198 | <0.01 | <0.01 |
| Пол | 0.03 | 0.29 | 0.04 | 0.29 | 0.57 | 1 | 0.45 | 0.32 |
| Возраст | 1.33 | 1.39 | 0.04 | 0.2 | 1478 | 45.5 | <0.01 | <0.01 |
| Стадия цикла | 0.15 | 0.01 | 0.04 | 0.18 | 18 | 0.005 | <0.01 | 0.32 |
Иная картина складывается в случае нециклических флуктуирующих колебаний численности, характерных для XXI в.: стадия цикла не оказывает значимого влияния на вес зверьков (коэффициент регрессии = 0.01 ± 0.18; статистика Вальда 0.005; р = 0.32) (табл. 1).
Таким образом, мы обнаружили, что вес тела связан с уровнем численности зверьков только при циклических колебаниях численности популяции. Ранее сходные эффекты нами были обнаружены при анализе и других популяционных показателей при сравнении циклических и флуктуирующих популяций. Так, частота асимметричного проявления краниологических признаков (флуктуирующая асимметрия черепа) коррелирует с численностью только в циклических популяциях [3].
Полученные результаты мы интерпретировали следующим образом. Циклические колебания численности обычно происходят при стабильных условиях среды и они регулируются эндогенными факторами. Мы предполагаем, что именно благодаря этому и возникает корреляция популяционных показателей, в данном случае веса тела зверьков с численностью популяции. В случае нециклических колебаний численность регулируется внешними факторами, в первую очередь метеоусловиями, у которых нет обратной связи с популяционными показателями.
Изменение веса зверьков при перепадах численности может использоваться как индикатор, дающий возможность отличить циклические колебания численности от флуктуаций при непродолжительном ряде исследований. А анализ динамики популяций – довольно чувствительный подход, позволяющий оценить последствия глобального изменения климата для биоразнообразия, что является актуальной задачей, так как результатом таких изменений может быть как модификация биологии отдельных видов, так и сообщества в целом.
Список литературы
Hansson L. Cycles and travelling waves in rodent dynamics: A comparison // Acta Theriologica. 2002. V. 47 (Suppl. 1). P. 9–22. https://doi.org/10.1007/bf03192477
Sheftel B.I. Role of different mechanisms in type determination of population dynamics for small mammals from boreal forestry zone. Ch 9. In: Biological diversity and nature conservation: theory and practice for teaching. Ed. I. Spellerberg, J. Slowik, M. Mühlenberg and Y.Y. Dgebuadze. M.: KMK, 2010. P. 130–143. ISBN 978-5-87317-636-6
Zakharov V.M., Pankakoski E., Sheftel B.I., et al. // Amer. Naturalist. 1991. V. 138. № 4. P. 797–810.
Boonstra R., Boag P.T. A test of the Chitty hypothesis. Inheritance of life-history traits in meadow voles Microtus pennsylvanicus // Evolution. 1987. V. 41. P. 929–947.
Chitty D. Mortality among voles (Microtus agrestis) at Lake Vyrnwy, Montgomeryshire in 1936-9 // Phil. Trans. R. Soc. Lond. 1952. V. B36. P. 505–552.
Sundell J., Norrdahl K. Body size-dependent refuges in voles: An alternative explanation of the Chitty effect // Annales Zoologici Fennici. 2002. № 39 (4). P. 325–333.
Ims R.A., Henden J.A., Killengreen S.T. Collapsing population cycles // Trends in Ecology and Evolution. 2008. V. 23. № 2. P. 79–86. https://doi.org/10.1016/j.tree.2007.10.010
Охотина М.В., Костенко В.А. Полиэтиленовая пленка – перспективный материал для изготовления ловчих заборчиков // Фауна и экология наземных позвоночных юга Дальнего Востока СССР. Владивосток. 1974. С. 193–196.
Шефтель Б.И. Методы учета численности мелких млекопитающих. // Russian Journal of Ecosystem Ecology. 2018. V. 3 (3). P. 1–21. https://doi.org/10.21685/2500-0578-2018-3-4
Коросов А.В. Специальные методы биометрии // Петрозаводск: Изд-во ПетрГУ. 2007. 364 c. ISBN 978-5-8021-0615-0
Sheftel B.I. Long-term and seasonal dynamics of shrews in Central Siberia // Annales Zoologici Fennici. Helsinki: Finnish Zoological Publishing Board, 1989. P. 357–369.
Захаров В.М., Шефтель Б.И., Дмитриев С.Г. Изменение климата и популяционная динамика: возможные последствия (на примере мелких млекопитающих в Центральной Сибири) // Успехи современной биологии. 2011. Т. 131. № 5. С. 435–439.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни