

Доклады Российской академии наук. Науки о жизни, 2020, T. 493, № 1, стр. 427-432
ЛИПИДНЫЙ СТАТУС СЕРЫХ КИТОВ Eschrichtius robustus В ПИЛЬТУНСКОМ НАГУЛЬНОМ РАЙОНЕ
М. С. Мамаев 1, С. А. Мурзина 2, С. Н. Пеккоева 2, В. П. Воронин 2, академик РАН В. В. Рожнов 1, *
1 Институт проблем экологии и эволюции
им. А.Н. Северцова Российской академии наук
Москва, Россия
2 Институт биологии КарНЦ РАН, ФИЦ “Карельский научный центр РАН”
Петрозаводск, Россия
* E-mail: rozhnov.v@gmail.com
Поступила в редакцию 20.03.2020
После доработки 22.04.2020
Принята к публикации 22.04.2020
Аннотация
Впервые с использованием методов биохимического анализа показана взаимосвязь липидного и жирнокислотного состава с индивидуальными, половыми и возрастными особенностями у серых китов Eschrichtius robustus в пильтунском нагульном районе.
Западные серые киты (они же западная подпопуляция серых китов [1] или охотско-корейская популяция) населяют северную часть акватории у Тихоокеанского побережья Азии [2]. До момента обнаружения группы особей в районе северо-восточного побережья о. Сахалин в 1983 г. [3] они считались вымершими. В настоящее время западные серые киты занесены в Красную книгу Российской Федерации с категорией 1 “находящиеся под угрозой исчезновения” [4] и Красный список Международного союза охраны природы с категорией “находящиеся под угрозой исчезновения” [1].
Один из основных периодов жизненного цикла серых китов – нагульные миграции. В течение периода размножения и длительного миграционного перехода к местам нагула киты не питаются, используя в качестве источника энергии запасы подкожного жира, и в места нагула в Охотском море, в основном у северо-восточного берега Сахалина, приходят в истощенном состоянии. К Сахалину киты начинают приходить в конце мая–июне, а в июле–августе их численность здесь становится наибольшей. Ключевым местом их кормления является пильтунский нагульный район [5] – мелководная акватория в месте впадения залива Пильтун в Охотское море. Практически весь летний период киты кормятся, причем часто держатся на одних и тех же местах. Приуроченность особей и их группировок к конкретным нагульным пастбищам связана, прежде всего, со стабильно высокой биомассой кормовых объектов китов, среди которых в пильтунском нагульном районе доминирующими являются планктобентические организмы – амфиподы и изоподы [6]. В нагульный непродолжительный период за счет активного питания происходит накопление должного количества жировых запасов в организме китов, которые обеспечивают энергозатратные метаболические реакции и процессы, в том числе в репродуктивный период зимой, на последующий год [7, 8]. В конце сентября начинается миграция китов к местам зимовок.
Известно [9], что состав липидов подкожного жира напрямую отражает качественные и количественные особенности кормовой базы животного. Липидный статус организма, как один из интегральных показателей метаболизма, позволяет охарактеризовать физиологическое состояние как отдельных особей, так и целой популяции. Изучение жирнокислотного профиля кожи и подкожного жира, с особым вниманием к биомаркерным жирным кислотам и их соотношениям, позволяет охарактеризовать трофическое поведение и предпочтения китов, выявить их отдельные группировки, приуроченность к различным нагульным пастбищам, а также охарактеризовать состав и состояние кормовой базы и ее динамику в местах нагула [10].
Изучение липидного статуса западных серых китов ранее не проводилось, тогда как для восточной (чукотско-калифорнийской) популяции серых китов проведен сравнительный анализ липидного профиля кожи и подкожного жира, полученных прижизненной биопсией, с учетом возраста, пола, состояния и репродуктивного статуса особей [11].
В настоящей работе впервые представлены результаты изучения липидного состава в подкожном жире и липидного и жирнокислотного состава в коже половозрелых особей западных серых китов и детенышей (сеголеток) в нагульный период в пильтунском нагульном районе. Отдельно следует отметить прижизненный характер сбора проб из-за природоохранного статуса исследуемых животных.
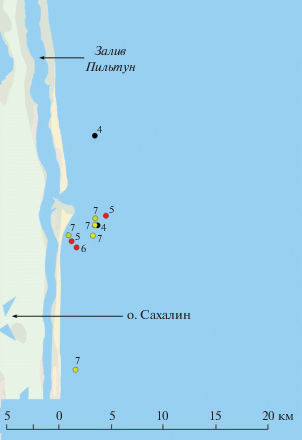
Наблюдения и сбор проб исследованных животных проводили в пильтунском нагульном районе. Биопсию проб осуществляли с помощью арбалета и стрелы с пробоотборником. Собранная проба представляла собой биопсию ткани с фиксацией в 96%-м этиловом спирте. Исследованные образцы были двух типов: первый – представлял собой столбик биопсии кожи, второй – столбик биопсии кожи с подкожным слоем жира (до 1 см). Общее количество проанализированных образцов составило 18, из них семь собраны в пильтунском нагульном районе в летний период (июль–август) 2013 г., девять в этот же период в 2014 г. и один образец в 2018 г. Кроме того, для сравнения использована одна проба, полученная от особи из чукотского нагульного района летом 2017 г.
Возраст серых китов определяли во время полевых наблюдений: животные старше одного года считаются взрослыми и обитают самостоятельно.
Отдельных особей различали с помощью фотоидентификации, которую проводили по методу, описанному в Специальном издании № 12 Международной Китобойной Комиссии [12].
Для определения пола китов использовали генетический анализ собранных образцов. Тотальную ДНК выделяли с использованием набора реагентов Diatom™ DNA Prep 200 (OOO “Лаборатория Изоген”, Москва, Россия). Половую принадлежность особей определяли на основе метода с использованием флуоресцентно меченых праймеров [13]. Анализ ПЦР-продуктов проводили на секвенаторе 3500 Genetic Analyzer (Thermo Fisher Scientific) в присутствии размерного стандарта для контроля соответствия длин получаемых фрагментов ожидаемым.
Оценку липидного статуса исследованных образцов кожи и подкожного жира китов проводили по содержанию общих липидов (ОЛ) и их липидных классов – диацилглицеринов (ДАГ), триацилглицеринов (ТАГ), холестерина (ХС), эфиров холестерина (ЭХС), общих фосфолипидов (ФЛ) и их отдельных классов: в том числе фосфатидилхолина (ФХ), фосфатидилэтаноламина (ФЭА), фосфатидилсерина (ФС), фосфатидилинозитола (ФИ), сфиногомиелина (СФМ), лизофосфатидилхолина (ЛФХ), диацилглицеринов (ДАГ), триацилглицеринов (ТАГ), холестерина (ХС), эфиров холестерина (ЭХС), а также свободных жирных кислот (СЖК) и жирных кислот общих липидов. Использованы индексы соотношений отдельных жирных кислот и их групп: 16:0/18:1(n-9), характеризующего скорость метаболизма липидов, 18:1(n-9)/18:1(n-7), показывающего степень плотоядности у гидробионтов, (n-6)/(n-3) полиненасыщенных жирных кислот (ПНЖК), демонстрирующего количественное соотношение (n-6) и (n-3) семейств ПНЖК. Подготовка материала, методы качественного и количественного анализа липидов (высокоэффективная тонкослойная, высокоэффективная жидкостная и газовая хроматография), а также статистическая обработка результатов описаны ранее [14]. Полученные на данном этапе исследования данные обработаны с помощью описательной статистики. В рамках настоящей статьи приводятся индивидуальные ЖК-профили конкретных особей серых китов, которые при соотнесении таковых с морфологическими и физиологическими характеристиками позволяют оценивать состояние каждой особи.
Полученные результаты, отражающие минимальные, максимальные и средние значения липидных классов в исследованных образцах кожи и кожи с подкожным жиром западных серых китов, приведены в табл. 1.
Таблица 1.
Среднее значение (M), минимум (min), максимум (max) и стандартное отклонение (σ) общих липидов (ОЛ), липидных классов (% сухой массы) и некоторых жирных кислот (ЖК) (% суммы ЖК) в исследованных образцах кожи и кожи с подкожным жиром западных серых китов
| Липидные классы, жирные кислоты | Кожа+жир | Кожа | ||||||
|---|---|---|---|---|---|---|---|---|
| M | min | max | σ | M | min | max | σ | |
| ОЛ | 16.79 | 11.48 | 23.22 | 3.89 | 7.61 | 1.91 | 24.84 | 6.94 |
| ФЛ | 4.35 | 2.39 | 6.09 | 1.33 | 1.96 | 0.36 | 7.30 | 1.99 |
| ФИ | 0.09 | 0.06 | 0.15 | 0.03 | 0.03 | 0.00 | 0.11 | 0.04 |
| ФС | 0.42 | 0.19 | 0.63 | 0.15 | 0.14 | 0.03 | 0.36 | 0.11 |
| ФЭА | 0.35 | 0.09 | 1.06 | 0.34 | 0.35 | 0.07 | 0.93 | 0.24 |
| ФХ | 3.20 | 1.66 | 4.61 | 0.89 | 1.32 | 0.22 | 6.07 | 1.68 |
| ЛФХ | 0.23 | 0.06 | 0.53 | 0.15 | 0.08 | 0.02 | 0.25 | 0.07 |
| СФМ | 0.02 | 0.00 | 0.06 | 0.02 | 0.01 | 0.00 | 0.05 | 0.02 |
| ДАГ | 1.59 | 1.03 | 2.19 | 0.38 | 0.50 | 0.23 | 1.45 | 0.38 |
| ХС | 3.48 | 2.76 | 4.74 | 0.65 | 2.00 | 0.56 | 5.78 | 1.48 |
| СЖК | 0.29 | 0.12 | 0.73 | 0.25 | 0.08 | 0.02 | 0.32 | 0.09 |
| ТАГ | 3.48 | 2.76 | 4.74 | 0.65 | 1.75 | 0.56 | 5.78 | 1.46 |
| ЭХС | 0.65 | 0.38 | 0.87 | 0.17 | 0.70 | 0.13 | 2.05 | 0.70 |
| 16:0 | 10.47 | 8.54 | 11.38 | 0.96 | 10.06 | 7.40 | 16.04 | 2.51 |
| 18:0 | 5.83 | 3.73 | 14.64 | 3.91 | 13.70 | 3.54 | 18.38 | 4.08 |
| 17:1(n-7) | 0.48 | 0.29 | 0.71 | 0.15 | 0.34 | 0.20 | 0.66 | 0.13 |
| 18:1(n-9) | 30.25 | 24.57 | 35.05 | 4.38 | 23.52 | 19.41 | 27.74 | 3.00 |
| 18:1(n-7) | 7.33 | 5.98 | 9.72 | 1.40 | 7.19 | 4.87 | 10.75 | 1.47 |
| 20:1(n-11) | 0.17 | 0.08 | 0.33 | 0.08 | 0.36 | 0.00 | 2.62 | 0.76 |
| 20:1(n-9) | 0.86 | 0.30 | 1.42 | 0.39 | 0.96 | 0.30 | 2.06 | 0.48 |
| 22:1(n-11) | 0.03 | 0.00 | 0.08 | 0.03 | 0.12 | 0.00 | 0.69 | 0.21 |
| 18:2(n-6) | 0.65 | 0.39 | 1.92 | 0.56 | 1.09 | 0.51 | 1.92 | 0.45 |
| 20:4(n-6) | 0.83 | 0.34 | 3.11 | 1.01 | 2.41 | 0.81 | 3.73 | 0.95 |
| 20:5(n-3) | 2.19 | 0.90 | 8.67 | 2.86 | 8.11 | 4.97 | 10.22 | 1.81 |
| Σ(n-6)/Σ(n-3) ПНЖК | 0.42 | 0.32 | 0.48 | 0.06 | 0.33 | 0.14 | 0.50 | 0.13 |
| 16:0/18:1(n-9) | 0.36 | 0.30 | 0.44 | 0.05 | 0.44 | 0.27 | 0.76 | 0.14 |
| 18:1(n-9)/18:1(n-7) | 4.33 | 2.53 | 5.90 | 1.32 | 3.35 | 2.10 | 4.11 | 0.57 |
С помощью биохимического анализа установлена индивидуальная вариабельность содержания липидов у половозрелых особей и детенышей (табл. 2), связанная как с половыми и возрастными различиями (прежде всего по содержанию ОЛ, ФЛ, ФХ, ФЭА, ФИ, ДАГ, СЖК, ТАГ), так и сроками и продолжительностью нагульного сезона, трофическими условиями нагульных пастбищ. Более того, значения индексов 16:0/18:1(n-9) и (n-6)/(n-3) ПНЖК различались и были ниже в образцах кожи самок из пильтунского нагульного района (особи № 8 и № 11), чем у самки, встреченной однократно в этом районе (особь № 18). Для самца, обитающего в чукотском нагульном районе, значение 16:0/18:1(n-9) было выше, а индекса (n-6)/(n-3) ПНЖК ниже, чем у самок из пильтунского нагульного района. Следует отметить, что в образцах кожи и подкожного жира самок и самцов из пильтунского нагульного района значение индекса 16:0/18:1(n-9) различалось незначительно. Полученные результаты позволяют предположить, что различия уровня липогенеза у исследованных особей связаны с трофическими условиями местообитания и мобильной активностью животного.
Таблица 2.
Состав общих липидов (ОЛ) и их липидных классов (% сухой массы), а также некоторых жирных кислот (ЖК) (% суммы ЖК) в образцах тканей западных серых китов
| Половозрелые особи | Детеныши | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ткань | Кожа | Кожа + жир | Кожа | Кожа + жир | ||||||||||||||||
| № особи | 8 | 11 | 17 | 18 | 4 | 5 | 7 | 1 | 2 | 6 | 10 | 12 | 14 | 15 | 3 | 9 | 13 | 16 | ||
| Пол | ♀ | ♀ | ♂ | ♀ | ♂ | ♂ | ♀ | ♀ | ♀ | ♂ | ♀ | ♀ | ♀ | ♂ | ♀ | ♂ | ♀ | ♂ | ||
| Липидные классы | % сухой массы | |||||||||||||||||||
| ОЛ | 4.27 | 3.56 | 9.01 | 24.84 | 14.7 | 16.23 | 11.48 | 8.09 | 5.01 | 4.62 | 1.91 | 3.44 | 3.03 | 15.88 | 23.22 | 14.84 | 20.36 | 16.68 | ||
| ФЛ | 0.83 | 0.87 | 1.97 | 7.3 | 3.88 | 4.04 | 2.39 | 2.82 | 1.47 | 0.96 | 0.36 | 0.88 | 0.81 | 3.29 | 6.08 | 3.69 | 6.09 | 4.27 | ||
| ФИ | 0.01 | 0 | 0.02 | 0.05 | 0.07 | 0.06 | 0.09 | 0.02 | 0.08 | 0 | 0.02 | 0 | 0 | 0.11 | 0.15 | 0.09 | 0.07 | 0.07 | ||
| ФС | 0.04 | 0.09 | 0.2 | 0.14 | 0.4 | 0.34 | 0.3 | 0.36 | 0.24 | 0.06 | 0.03 | 0.07 | 0.05 | 0.27 | 0.55 | 0.19 | 0.63 | 0.51 | ||
| ФЭА | 0.18 | 0.26 | 0.3 | 0.48 | 0.12 | 0.09 | 0.11 | 0.93 | 0.47 | 0.17 | 0.07 | 0.26 | 0.22 | 0.47 | 0.44 | 0.29 | 1.06 | 0.37 | ||
| ФХ | 0.55 | 0.48 | 1.32 | 6.07 | 3.09 | 3.25 | 1.66 | 1.37 | 0.61 | 0.63 | 0.22 | 0.48 | 0.48 | 2.27 | 4.61 | 2.89 | 3.72 | 3.18 | ||
| ЛФХ | 0.03 | 0.03 | 0.08 | 0.25 | 0.16 | 0.21 | 0.2 | 0.11 | 0.06 | 0.07 | 0.02 | 0.04 | 0.04 | 0.14 | 0.25 | 0.17 | 0.53 | 0.06 | ||
| СФМ | 0 | 0 | 0 | 0.05 | 0.02 | 0.03 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.01 | 0.06 | 0.01 | 0.01 | 0.01 | ||
| ДАГ | 0.25 | 0.39 | 0.62 | 1.45 | 1.39 | 1.57 | 1.03 | 0.43 | 0.34 | 0.32 | 0.28 | 0.27 | 0.23 | 0.96 | 2.19 | 1.56 | 1.97 | 1.43 | ||
| ХС | 1.28 | 1.27 | 1.87 | 5.78 | 3.26 | 3.82 | 2.76 | 3.33 | 1.89 | 1.44 | 0.56 | 1.18 | 0.83 | 2.58 | 4.74 | 3.11 | 3.59 | 3.11 | ||
| СЖК | 0.03 | 0.03 | 0.12 | 0.32 | 0.12 | 0.21 | 0.15 | 0.08 | 0.04 | 0.1 | 0.03 | 0.02 | 0.02 | 0.12 | 0.73 | 0.12 | 0.55 | 0.14 | ||
| ТАГ | 1.28 | 1.27 | 1.87 | 5.78 | 3.26 | 3.82 | 2.76 | 0.62 | 1.89 | 1.44 | 0.56 | 1.18 | 0.83 | 2.58 | 4.74 | 3.11 | 3.59 | 3.11 | ||
| ЭХС | 0.4 | 0.3 | 2.05 | 2 | 0.64 | 0.55 | 0.38 | 0.82 | 0.73 | 0.23 | 0.13 | 0.17 | 0.18 | 0.71 | 0.87 | 0.58 | 0.66 | 0.86 | ||
| 16:0 | 8.1 | 8.75 | 16.04 | 12.09 | 11.24 | 10.02 | 8.54 | 9.46 | 10.93 | 8.54 | 11.84 | 9.32 | 8.21 | 7.4 | 10.61 | 10.59 | 10.89 | 11.38 | ||
| 18:0 | 13.97 | 18.38 | 12.67 | 11.12 | 4.86 | 3.73 | 14.64 | 15.87 | 16.51 | 14.64 | 11.37 | 17.04 | 15.61 | 3.54 | 3.93 | 4.42 | 4.72 | 4.51 | ||
| 17:1(n-7) | 0.39 | 0.66 | 0.39 | 0.34 | 0.71 | 0.62 | 0.38 | 0.2 | 0.21 | 0.38 | 0.32 | 0.38 | 0.29 | 0.23 | 0.45 | 0.29 | 0.35 | 0.55 | ||
| 18:1(n-9) | 27.74 | 22.31 | 21.06 | 26.22 | 34.52 | 35.05 | 26.27 | 19.41 | 20.04 | 26.27 | 21.17 | 24.8 | 22.59 | 27.09 | 30.75 | 26.74 | 24.57 | 33.86 | ||
| 18:1(n-7) | 7.04 | 6.16 | 6.5 | 6.92 | 6.12 | 5.98 | 7.46 | 7.06 | 4.87 | 7.46 | 6.7 | 7.13 | 10.75 | 8.5 | 6.99 | 8.66 | 9.72 | 6.36 | ||
| 20:1(n-11) | 2.62 | 0 | 0 | 0.08 | 0.16 | 0.13 | 0.24 | 0.13 | 0 | 0.24 | 0.25 | 0.11 | 0.06 | 0.46 | 0.33 | 0.13 | 0.14 | 0.08 | ||
| 20:1(n-9) | 0.66 | 0.89 | 2.06 | 1.25 | 1.42 | 1.12 | 0.3 | 0.53 | 0.92 | 0.3 | 1.36 | 1.07 | 0.83 | 0.66 | 0.65 | 0.94 | 1.08 | 0.51 | ||
| 22:1(n-11) | 0.69 | 0 | 0 | 0.05 | 0.08 | 0.03 | 0.04 | 0 | 0 | 0.04 | 0.31 | 0.11 | 0.04 | 0.03 | 0 | 0 | 0.04 | 0 | ||
| 18:2(n-6) | 1.28 | 1.13 | 0.85 | 0.66 | 0.4 | 0.54 | 1.92 | 1.03 | 1.89 | 1.92 | 0.87 | 0.95 | 0.87 | 0.51 | 0.39 | 0.47 | 0.4 | 0.4 | ||
| 20:4(n-6) | 3.73 | 2.87 | 1.01 | 2.46 | 0.62 | 0.47 | 3.11 | 2.71 | 2.22 | 3.11 | 1.43 | 2.96 | 3.24 | 0.81 | 0.4 | 0.44 | 0.34 | 0.42 | ||
| 20:5(n-3) | 8.04 | 9.83 | 5.83 | 4.97 | 1.19 | 1.16 | 8.67 | 10.22 | 6.79 | 8.67 | 9.2 | 9.56 | 9.61 | 6.52 | 0.9 | 1.36 | 1.05 | 0.99 | ||
| Σ(n-6)/Σ(n-3) ПНЖК | 0.44 | 0.34 | 0.16 | 0.5 | 0.48 | 0.42 | 0.48 | 0.28 | 0.43 | 0.48 | 0.21 | 0.34 | 0.36 | 0.14 | 0.44 | 0.32 | 0.39 | 0.4 | ||
| 16:0/18:1(n-9) | 0.29 | 0.39 | 0.76 | 0.46 | 0.33 | 0.3 | 0.33 | 0.49 | 0.55 | 0.33 | 0.56 | 0.38 | 0.36 | 0.27 | 0.35 | 0.4 | 0.44 | 0.34 | ||
| 18:1(n-9)/18:1(n-7) | 3.9 | 3.6 | 3.2 | 3.8 | 5.6 | 5.9 | 3.5 | 2.75 | 4.11 | 3.52 | 3.16 | 3.48 | 2.1 | 3.19 | 4.4 | 3.09 | 2.53 | 5.32 | ||
Индекс 18:1(n-9)/18:1(n-7) в образцах кожи и подкожного жира у самцов и самки из одного и того же нагульного района различался и составил 5.6 и 5.9 для самцов и 3.5 для самок (среднее значение 4.33 ± 1.32), что указывает на половые различия в пищевых спектрах китов.
Для одной особи самца пильтунского нагульного района выявлен схожий липидный спектр кожи по сравнению с самками, при этом уровень насыщенных жирных кислот (НЖК) был равнозначен таковому для мононенасыщенных жирных кислот (МНЖК). Присутствие НЖК и МНЖК в организмах любых таксономических групп свидетельствует об их фундаментальной роли в поддержании общего хода обмена веществ в живых системах.
Для западных серых китов пильтунского нагульного района в пробах кожи с подкожным жиром (взрослые особи № 4, № 5, № 7) установлены наибольшие различия по количеству для таких доминирующих жирных кислот как МНЖК – 18:1(n-9), 18:1(n-7), НЖК – 16:0, 18:0 и ПНЖК – 18:2(n-6), 20:4(n-6), 20:5(n-3), а также минорных жирных кислот 17:1(n-7), группы жирных кислот (n-4) семейства, имеющих бактериальное происхождение, и 20:1(n-11), 20:1(n-9), 22:1(n-11), поступающих, как правило, за счет питания зоопланктоном. Полученные результаты по ключевым жирным кислотам и их соотношениям связаны с неоднородностью распределения кормового планктобентоса в этой акватории [6], и кормлением китов в разных местах акватории. Так, особь № 5 встречалась только рядом с устьем залива Пильтун, особь № 4 находилась рядом с устьем и севернее, а особь № 7 пришла к заливу с юга. Наибольшее содержание ОЛ, ФЛ, ФИ, ФХ, СФМ, ДАГ, ХС, ТАГ показано для особи № 5, в меньших количествах эти липиды содержались в пробах № 4 и 7.
Скорость липидного обмена и его особенности, в частности, включение пищевых жирных кислот в состав липидов жировой ткани серых китов, до сих пор изучены недостаточно. Однако согласно данным литературы, у тюленей и гренландских китов запасание пищевых жирных кислот в структуре энергетических липидов жировой ткани может происходить в период от 12 ч до нескольких дней, наибольшее – месяца [15].
Причины количественных различий липидов, в том числе жирных кислот у исследованных западных серых китов пильтунского нагульного района, могут быть связаны с разными условиями питания (видовым составом, массовостью, доступностью корма), неодинаковой тратой энергии при разных скоростях течения в различных участках акватории, плотностью особей, их трофическим поведением на участках.
Нами выявлены различия в содержании липидных классов в пробах кожи и жира взрослых китов и детенышей. При этом установлено, что два детеныша (особи № 9 и № 16) характеризуются липидными профилями, схожими с таковыми у взрослых животных, а для особей № 3 и № 13 показано повышенное содержание ОЛ, ФЛ, ДАГ, СЖК, ЭХС. Полученные результаты могут быть объяснены тем, что особи № 9 и № 16 уже перешли на самостоятельное питание, в то время как особи № 3 и № 13 продолжают кормиться молоком матери.
Интересным результатом сравнения жирных кислот у взрослых китов и детенышей является обнаружение высокой степени сходства по содержанию исследованных липидных классов в образцах кожи в паре “самка–детеныш” у особей № 11 и № 12 соответственно.
Четыре липидных класса СФМ, ФХ, ФЭА, ФИ имеют одинаковое значение у обеих особей, а уровни ОЛ, ФЛ и ФС отличаются на 0.12; 0.01 и 0.02%.
На основе полученных данных сформирована рабочая гипотеза о «метаболическом воспроизведении» профиля липидов кожи, особенно классов ФЛ, у самки и ее детеныша в определенный репродуктивный период. Эта гипотеза требует более детальных исследований, которые предполагается продолжить коллективом авторов. Подчеркнем, что липидный состав кожи детеныша № 12 существенно отличался от такового других исследованных детенышей.
Таким образом, нами впервые получены и обсуждаются параметры липидного и жирнокислотного состава западных серых китов, которые показывают связь с индивидуальными, половыми и возрастными особенностями животных. Анализ жирных кислот, биомаркерных для некоторых видов кормовых объектов исследованных млекопитающих, позволяет обсуждать специфику их аккумуляции в жировых депо китов. Сформирована рабочая гипотеза о “метаболическом воспроизведении” на основе анализа отдельных показателей липидного обмена для пары “самка–детеныш”.
Список литературы
Cooke J.G., Taylor B.L., Reeves R., Brownell Jr., R.L. Eschrichtius robustus (western subpopulation). The IUCN Red List of Threatened Species 2018: e.T8099A50345475.
Бурдин А.М., Филатова О.А., Хойт Э. Морские млекопитающие России: справочник определитель. Киров: Волго-Вятское книжное изд-во, 2009. 210 с.
Blokhin S.A., Maminov M.K., Kosygin G.M. // Report of the International Whaling Commission. 1985. V. 35. P. 375–376.
Красная книга Российской Федерации (животные). М.: АСТ-Астрель, 2001. 862 с.
Бурдин А.М. 20 лет исследований серых китов (Eschrichtius robustus) у северо-восточного Сахалина, Россия, итоги (1997–2017) // Сборник тезисов Х Международной конференции “Морские млекопитающие Голарктики”. Архангельск, 29 октября–02 ноября 2018 г. С. 27.
Фадеев В.И. Состояние бентоса в районах нагула серых китов в 2013 году. Отчет о НИР. WGWAP-14\7. Владивосток, 2014. 60 с.
Томилин А.Г. Звери СССР и прилежащих стран. Т. IX. Китообразные. М., 1957. 757 с.
Waugh C.A., Nichols P.D., Noad M.C., Nash S.B. // Marine ecology progress series. 2012. V. 471. P. 271–281.
Budge S.M., Iverson S.J., Koopman H.N. // Marine Mammal Science. 2006. V. 22. № 4. P. 759–801.
Ruchnonnet D., Boutoute M., Guinet C., Mayzaud P. // Mar Ecol Prog Ser. 2006. V. 311. P. : 165–174.
Krahn M.M., Ylitalo G.M., Burrows D.G., Calambokidis J., et al. // J. Cetacean Res. Manage. 2001. V. 3. № 1. P. 19–29.
Hammond P.S., Mizroch S.A., Donovan G.P. // Report of the Int. Whal. Comm. Special Issue 12. 1990. 440 p.
Jayasankar P., Anoop B., Rajagopalan M. // Current Sci. 2008. V. 94. № 11. P. 1513–1516.
Мурзина С.А., Нефедова З.А., Руоколайнен Т.Р., Васильева О.Б., Немова Н.Н. // Онтогенез. 2009. Т. 40. № 3. С. 208–214.
Budge S.M., Cooper M.H., Iverson S.J. // Physiological and Biochemical Zoology. 2004. V. 77. P. 682–687.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни