

Доклады Российской академии наук. Науки о жизни, 2020, T. 494, № 1, стр. 437-443
Стратегии дележа ограниченными ресурсами у детей и подростков в трех традиционных обществах Восточной Африки: социокультурные и генетические факторы
М. Л. Бутовская 1, *, Д. В. Карелин 2, Д. А. Дронова 1, В. О. Филатова 1, П. Р. Бутовская 3, академик РАН В. А. Тишков 1
1 Институт этнологии и антропологии
им. Н.Н. Миклухо-Маклая Российской академии наук
Москва, Россия
2 Институт географии Российской академии наук
Москва, Россия
3 Институт общей генетики им. Н.И. Вавилова Российской академии наук
Москва, Россия
* E-mail: marina.butovskaya@gmail.com
Поступила в редакцию 13.04.2020
После доработки 19.04.2020
Принята к публикации 22.04.2020
Аннотация
Эгалитарность, стремление к равенству и альтруизм являются важнейшими эволюционно-стабильными стратегиями в человеческом обществе. В рамках данного исследования представлены данные по результатам экономических игр на дележ с потенциальным другом или незнакомым сверстником в трех этнических группах Восточной Африки, хадза, ирак и меру. Общая выборка составила 583 ребенка и подростка, средний возраст 13.5 ± 3.1 года. Помимо этого, для 162 меру был проведен анализ ДНК и прогенотипирован однонуклеотидный полиморфизм гена OXTR rs53576. Установлено наличие выраженной индивидуальной вариабельности по принятию решений о дележе с потенциальным партнером. Достоверно чаще дети и подростки вели себя альтруистично в направлении друзей по сравнению с незнакомцами. Альтруизм достоверно чаще демонстрировали носители варианта GG OXTR rs53576 как к друзьям (U = 3376.500, p = 0.047; OR = 3.075, p = 0.032), так и к незнакомцам (U = 3478.000, p = 0.025; OR = 3.133, p = 0.007). Показаны также достоверные межгрупповые различия по эгалитарианизму, эгоизму и альтруизму. Полученные данные могут свидетельствовать о наличие положительного группового отбора в направлении альтруистов.
Эгалитарность, стремление к равенству и альтруизм являются важнейшими эволюционно-стабильными стратегиями в человеческом обществе [1, 17]. По уровню их развития Homo sapiens существенно превосходит все другие виды животных, включая современных шимпанзе и бонобо [13]. Особое внимание антропологи уделяют беспрецедентному развитию альтруизма, появляющегося, в первую очередь, в дележе пищей и другими ограниченными ресурсами во всех человеческих обществах [1, 8, 9]. Ведущим методом количественной оценки склонности к про-социальному поведению у человека служат экономические эксперименты, принцип которых был заимствован из теории игр. Показано, что общества с разным культурно-хозяйственным типом варьируют по уровню кооперации и практик дележа ресурсами. Различия касаются также и жесткости наказания в отношении обманщиков (лиц, не желающих делиться своими ресурсами с другими и при этом получающих блага от других в одностороннем порядке). Готовность делиться ресурсами варьирует также на индивидуальном уровне, служит одним из проявлений эмпатии и может быть связана с генетической компонентой, в частности с геном рецепторов окситоцина OXTR [14].
Цель исследования – выявить общие закономерности и культурно-специфические особенности стратегий дележа ограниченными ресурсами у детей и подростков из трех традиционных обществ Восточной Африки; оценить генетическую предрасположенность к дележу на примере ограниченной выборки детей одного из изученных обществ (в качестве гена-кандидата выбран ген OXTR, однонуклеотидный полиморфизм rs53576). Исследования проводились в 2015 и 2019 гг. Объекты исследования: дети и подростки из трех традиционных обществ Северной Танзании: хадза (бродячие охотники-собиратели), ирак (агро-пасторалисты), меру (фермеры). Выборка составила 583 человека, включая 130 хадза, 164 ирак и 289 меру. Сведения по экологии, социальной организации и хозяйственному типу этих обществ представлены в наших ранних работах и публикациях других авторов [2, 3, 15, 16]. После сбора демографических данных (пол, возраст, этническая принадлежность, состав семьи), а также краткого опросника на установки к кооперации, детям и подросткам предлагали по три игры на распределение ограниченных ресурсов в отношении друга и незнакомого сверстника (то есть в общей сложности каждый участник играет 6 раз). Схема игры была ранее разработана Е. Фером с соавторами и использована для анализа возрастной динамики формирования установок на эгалитарность, альтруизм и эгоизм у детей в современном западном обществе [10].
Для ограниченной выборки детей и подростков меру (162 человека) провели сбор буккальных проб, из которых была выделена ДНК с помощью Diatom DNA Prep 200 (Лаборатория “Изоген”, Москва, Россия). Качество и концентрация ДНК определялась на приборе Nanodrop. Однонуктеотидный полиморфизм гена OXTR (rs53576) был прогенотипирован с помощью Taqman 5′ exonuclease assay (Applied Biosystems). Реакцию амплификации проводили в 5 мкл смеси ПЦР, реакционная смесь содержала 5 нг геномной ДНК, 2.5 μl Taqman Мастер Микса и 0.25 μl 40 кратного флуоресцентного зонда, соответствующего анализируемому полиморфизму [4]. Каталожный номер использованной флуоресцентной пробы 43511379 (https://www.thermofisher.com) для анализа TaqMan генотипирование SNP (идентификационный номер: C___3290335_10). Детекция флуоресценции проводилась по каналам FAM и VIC. Для анализа результатов генотипирования по конечной точке использовали прибор ABI PRISM 7900HT, а для интерпретации результатов программное обеспечение SDS v2.3 (Applied Biosystems).
Роль факторов пола и групповой принадлежности в выборе решений по каждой из игр (в отношении другу и незнакомого сверстника соответственно) оценивали с помощью двухфакторного дисперсионного анализа с независимыми переменными “пол” и “этническая группа”, а также взаимодействия этих факторов (табл. 1).
Таблица 1.
Двухфакторный дисперсионный анализ по шести вариантам зависимых переменных при анализе социального поведения детей трех этнических групп (Танзания), где в качестве независимых переменных использовались пол, этническая группа и взаимодействие факторов пол*этническая группа
| Зависимая переменная | Показатели полученной модели | Объясненная сумма квадратов отклонений | Число степе-ней свободы | Средний квадрат отклонений | F | Р | Эффективный размер (partial eta squared) |
|---|---|---|---|---|---|---|---|
| Просоциальное с другом | Пол | 0.122 | 1 | 0.122 | 1.401 | 0.237 | 0.002 |
| Этническая группа | 1.961 | 2 | 0.980 | 11.246 | 0.000 | 0.038 | |
| Пол * Этническая группа | 0.170 | 2 | 0.085 | 0.972 | 0.379 | 0.003 | |
| Ошибка | 50.125 | 575 | 0.087 | ||||
| Сумма | 2150 | 581 | |||||
| Зависть к другу | Пол | 0.160 | 1 | 0.160 | 0.440 | 0.508 | 0.001 |
| Этническая группа | 6.258 | 2 | 3.129 | 8.577 | 0.000 | 0.029 | |
| Пол * Этническая группа | 2.986 | 2 | 1.493 | 4.093 | 0.017 | 0.014 | |
| Ошибка | 209.396 | 574 | 0.365 | ||||
| Сумма | 3309 | 580 | |||||
| Дележ с другом | Пол | 0.027 | 1 | 0.027 | 0.103 | 0.749 | 0.000 |
| Этническая группа | 11.248 | 2 | 5.624 | 21.394 | 0.000 | 0.070 | |
| Пол * Этническая группа | 1.334 | 2 | 0.667 | 2.537 | 0.080 | 0.009 | |
| Ошибка | 148.794 | 566 | 0.263 | ||||
| Сумма | 2164 | 572 | |||||
| Просоциальное с незнакомцем | Пол | 0.015 | 1 | 0.015 | 0.107 | 0.744 | 0.000 |
| Этническая группа | 3.648 | 2 | 1.824 | 12.960 | 0.000 | 0.043 | |
| Пол * Этническая группа | 0.377 | 2 | 0.188 | 1.339 | 0.263 | 0.005 | |
| Ошибка | 80.778 | 574 | 0.141 | ||||
| Сумма | 2011 | 580 | |||||
| Зависть к незнакомцу | Пол | 0.017 | 1 | 0.017 | 0.042 | 0.838 | 0.000 |
| Этническая группа | 11.840 | 2 | 5.920 | 14.864 | 0.000 | 0.049 | |
| Пол * Этническая группа | 5.602 | 2 | 2.801 | 7.033 | 0.001 | 0.024 | |
| Ошибка | 228.613 | 574 | 0.398 | ||||
| Сумма | 3013 | 580 | |||||
| Дележ с незнакомцем | Пол | 0.044 | 1 | 0.044 | 0.168 | 0.682 | 0.000 |
| Этническая группа | 20.969 | 2 | 10.484 | 39.811 | 0.000 | 0.123 | |
| Пол * Этническая группа | 1.828 | 2 | 0.914 | 3.470 | 0.032 | 0.012 | |
| Ошибка | 149.849 | 569 | 0.263 | ||||
| Сумма | 2040 | 575 |
Хадза в целом были менее склонны принимать решения по дележу в пользу друзей по сравнению с меру (тест Bonferroni: p = 0.001). В отношении незнакомых сверстников хадза также были менее альтруистичны по сравнению с ирак (тест Bonferroni: p = 0.042) и меру (тест Bonferroni: p < 0.0001). Ирак были менее склонны делиться с незнакомцами по сравнению с меру (тест Bonferroni: p = = 0.002).
В табл. 2 представлено абсолютное и процентное соотношение детей и подростков, предпочитавших следовать одной из стратегий (эгалитарная, альтруистическая, эгоистическая и смешанная) в отношении друзей и незнакомых сверстников в каждой группе. T-тест для парных выборок показал, что в целом участники были более позитивно настроены в отношении друзей.
Таблица 2.
Обобщенные варианты основных стратегий поведения в отношении друзей и незнакомых сверстников при распределении ограниченных ресурсов у хадза, ирак и меру (Танзания)
| Стратегия | Варианты стратегий | Хадза | Ирак | Меру | |||
|---|---|---|---|---|---|---|---|
| Друзья | Незнакомые | Друзья | Незнакомые | Друзья | Незнакомые | ||
| Эгалитаризм | Выраженный | 31 (23.9) | 27 (20.8) | 51 (31.1) | 49 (29.9) | 132 (45.7) | 134 (46.4) |
| Слабый | 15 (11.5) | 12 (9.2) | 13 (7.9) | 22 (13.4) | 15 (5.2) | 12 (4.2) | |
| Сумма | 46 (35.4) | 39 (30.0) | 64 (39.0) | 71 (43.3) | 147 (50.9) | 146 (50.6) | |
| Альтруизм | Выраженный | 7 (5.4) | 3 (2.3) | 0 | 2 (1.2) | 13 (4.5) | 16 (5.5) |
| Умеренный | 25 (19.2) | 20 (15.4) | 64 (39.0) | 48 (29.2) | 66 (22.8) | 48 (16.6) | |
| Слабый | 14 (10.8) | 12 (9.2) | 12 (7.3) | 11 (6.7) | 12 (4.2) | 10 (3.5) | |
| Сумма | 46 (35.4) | 35 (26.9) | 76 (46.3) | 61 (37.1) | 91 (31.5) | 74 (25.6) | |
| Эгоизм | Выраженный | 15 (11.5) | 30 (23.1) | 3 (1.8) | 2 (1.2) | 3 (1.0) | 3 (1.0) |
| Слабый | 2 (1.5) | 3 (2.3) | 0 | 0 | 2 (0.7) | 6 (2.1) | |
| Сумма | 17 (13.0) | 33 (25.4) | 3 (1.8) | 2 (1.2) | 5 (1.7) | 9 (3.1) | |
| Неопределенная | 21 (16.2) | 23 (17.7) | 21 (12.8) | 30 (18.3) | 46 (15.9) | 60 (20.8) | |
| Число респондентов | 130 | 130 | 164 | 164 | 289 | 289 | |
Достоверные различия получены для двух стратегий: умеренного альтруизма (чаще принимались решения в пользу друзей: t = 3.363, df = 575, p = 0.002) и выраженного эгоизма (чаще принимались негативные решения в отношении незнакомцев: t = –3.155, df = 579, p = 0.002). Обобщенные суммарные значения по всем играм, условно отражающие доброту и отзывчивость в целом по отношению к друзьям (6.08 ± 0.04) и незнакомцам (5.81 ± 0.05), в тесте для попарных выборок также указывают на альтруизм в пользу друзей (t = 6.309, df = 566, p < 0.00001).
Исследованная выборка меру находилась в равновесии Харди-Вайнберга (χ2 = 0.146, степень df = 1, p = 0.703). Частоты аллелей и генотипов составили (A = 0.19, G = 0.81, AA = 0.03, AG = 0.32, GG = 0.65). Различия ожидаемой и наблюдаемой гетерозиготности практически отсутствовали (Hobs = 0.315, Hexp = 0.306, коэффициент Райта = = –0.030).
Для ограниченной выборки детей меру (n = 162) проанализировали связь между выбором стратегий игрового поведения и полиморфизмом гена OXTR rs53576. С этими целями использовали таблицы сопряженности и вычислили отношения шансов Мантеля–Гензеля (табл. 3). Для данного анализа результаты игры перекодировали для всех случаев как: 1 – отказ от дележа; 2 – дележ с оппонентом (независимо от того, отдавалась ли ему одна конфета или две). Тест Манна–Уитни так же подтвердил связь между генотипами по гену OXTR и игрой на дележ (в этом случае использовали исходные коды: 1 – отказ от дележа; 2 – равное распределение конфет с оппонентом; 3 – полный отказ от конфет в пользу оппонента). Носители генотипа GG достоверно чаще оказывались альтруистами как в отношении друга (U = = 3376.500, p = 0.047), так и в отношении незнакомого сверстника (U = 3478.000, p = 0.025) (табл. 3, рис. 1, 2).
Таблица 3.
Таблица сопряженности выбора игровых стратегий с генотипами OXTR. Приведены оценки отношения шансов Мантеля–Гензеля
| Стратегия | OXTR | χ2 (P) | OR (P) | |||
|---|---|---|---|---|---|---|
| 1 = AA, AG | 2 = GG | |||||
| Просоциальное с другом | 1 | реальное | 2 | 5 | 0.116 (0.733) | 0.748 (0.734) |
| 2 | ожидаемое | 2.4 | 4.6 | |||
| 1 | реальное | 54 | 101 | |||
| 2 | ожидаемое | 53.6 | 101.4 | |||
| Просоциальное с незнакомцем | 1 | реальное | 11 | 11 | 2.680 (0.102) | 2.111 (0.107) |
| 2 | ожидаемое | 7.6 | 14.4 | |||
| 1 | реальное | 45 | 95 | |||
| 2 | ожидаемое | 48.4 | 91.6 | |||
| Зависть к другу | 1 | реальное | 2 | 5 | 0.116 (0.733) | 0.748 (0.734) |
| 2 | ожидаемое | 2.4 | 4.6 | |||
| 1 | реальное | 54 | 101 | |||
| 2 | ожидаемое | 53.6 | 101.4 | |||
| Зависть к незнакомцу | 1 | реальное | 5 | 9 | 0.662 (0.416) | 1.540 (0.419) |
| 2 | ожидаемое | 5.5 | 10.5 | |||
| 1 | реальное | 49 | 97 | |||
| 2 | ожидаемое | 50.5 | 95.5 | |||
| Дележ с другом | 1 | реальное | 10 | 7 | 4.940 (0.026) | 3.075 (0.032) |
| 2 | ожидаемое | 5.9 | 11.1 | |||
| 1 | реальное | 46 | 99 | |||
| 2 | ожидаемое | 51.1 | 94.9 | |||
| Дележ с незнакомцем | 1 | реальное | 16 | 12 | 7.697 (0.006) | 3.133 (0.007) |
| 2 | ожидаемое | 9.7 | 18.3 | |||
| 1 | реальное | 40 | 94 | |||
| 2 | ожидаемое | 46.3 | 87.7 | |||
Рис. 1.
Ассоциация генотипов OXTR rs53576 с альтруистической стратегий дележа с друзьями у детей меру. Условные обозначения по горизонтальной оси: 1 – генотипы AA и AG; 2 – генотип GG; по вертикальной оси: средние баллы по дележу со стандартными ошибками.
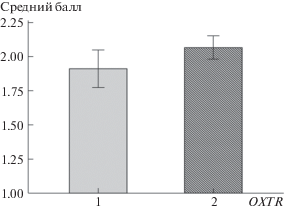
Рис. 2.
Ассоциация генотипов OXTR rs53576 с альтруистической стратегий дележа с незнакомыми сверстниками у детей меру. Условные обозначения по горизонтальной оси: 1 – генотипы AA и AG; 2 – генотип GG; по вертикальной оси: средние баллы по дележу со стандартными ошибками.

Таким образом, в целом, дети и подростки в традиционных обществах ориентированы на дележ и кооперацию (альтруизм) и следуют нормам просоциального поведения и справедливости при распределении ресурсов. Выявлена выраженная индивидуальная вариабельность по готовности жертвовать ценные ресурсы. Существенно значимым оказался фактор групповой принадлежности. Культурные нормы и предписанные правила поведения разнились по отношению к “своим” и “чужим”. Во всех группах прослежен парохиальный альтруизм [5, 9, 11]. Доля детей, демонстрирующих выраженный и умеренный альтруизм, варьировала. У меру и ирак выявлено больше альтруистов (в отношении друзей) по сравнению с хадза.
В выборке меру практически отсутствовали генотипы AA, ассоциированные другими авторами с аутизмом, низкой эмпатией [14]. Данные по ассоциации генотипов с игровыми стратегиями на дележ ограниченными ресурсами у детей меру свидетельствуют о том, что носители GG генотипа ведут себя достоверно более альтруистично в наиболее “дорогостоящей” для игрока стратегии, требующей реальной жертвы собственными ценными ресурсами по сравнению с генотипами АА и АG. К сожалению, мы не располагаем данными по генотипам детей хадза, участвующих в эксперименте. Поэтому, заметим лишь, что в игре на дележ с друзьями в процентном соотношении 41.9% детей вели себя как эгоисты и 58.1% как альтруисты, тогда как для меру и ирак это соотношение было достоверно в пользу преобладания альтруистов (12.5% к 87.5% и 21.7% к 78.3% соответственно). Соотношение носителей А аллеля к GG гомозиготам для меру составило 0.35 к 0.65, тогда как для выборки хадза оно составило 0.70 к 0.30 (эти данные получены нами ранее по взрослым из той же популяции, что и дети) [4].
Естественный отбор формирует психологические установки на потребность в общении с другими людьми и мотивацию к заботе и кооперации с окружающими [12]. Они усваиваются детьми и подростками в процессе интеграции в социальные сети конкретного общества. Неолитическая революция и потребности производящего хозяйства привели не только к росту межгрупповой конкуренции, но также стимулировали усиление внутригрупповой сплоченности и кооперации внутри коллектива. Предрасположенность к альтруизму могла фиксироваться на генетическом уровне, и группы, в состав которых входило больше альтруистов, оказывались более конкурентоспособными [6, 7]. Мы предполагаем, что полученные нами данные могут свидетельствовать в пользу более выраженного отбора на альтруистов в обществах агро-пасторалистов и земледельцев по сравнению с охотниками-собирателями. Достоверные различия в стратегиях дележа у детей и подростков из обществ с различными системами жизнеобеспечения указывают на роль социо-культурных факторов, формирующих общие нормы поведения при распределении ограниченных ресурсов и подтверждают идею о генно-культурной ко-эволюции.
Список литературы
Apicella C.L., Silk J.B. The evolution of human cooperation // Current Biology. 2019. V. 29. № 11. P. 447–450. https://doi.org/10.1016/j.cub.2019.03.036
Butovskaya M.L., Vasilyev V.A., Lazebny O.E., et al. Aggression, digit ratio, and variation in the androgen receptor, serotonin transporter, and dopamine D4 receptor genes in African foragers: the Hadza // Behavior genetics. 2012. V. 42. № 4. P. 647–662. https://doi.org/10.1007/s10519-012-9533-2
Butovskaya M.L., Burkova V.N., Karelin D.V. The Wameru of Tanzania: Historical Origin and their role in the process of National Integration // Social Evolution and History. 2016. V. 15. № 2. P. 160–180.
Butovskaya P.R., Lazebny O.E., Sukhodolskaya E.M., et al. Polymorphisms of two loci at the oxytocin receptor gene in populations of Africa, Asia and South Europe // BMC genetics. 2016. V. 17. № 1. P. 17. https://doi.org/10.1186/s12863-015-0323-8
Bernhard H., Fischbacher U., Fehr E. Parochial altruism in humans // Nature. 2006. V. 442. № 7105. P. 912–915. https://doi.org/10.1038/nature04981
Czárán T., Aanen D.K. The early evolution of cooperation in humans. On cheating, group identity and group size // Behaviour. 2016. V. 153. № 9–11. P. 1247–1266. https://doi.org/10.1163/1568539X-00003337
Gintis H., van Schaik C., Boehm C. Zoon politikon: The evolutionary origins of human socio-political systems // Behavioural processes. 2019. V. 161. P. 17–30. https://doi.org/10.1016/j.beproc.2018.01.007
Gurven M. To give and to give not: the behavioral ecology of human food transfers // Behavioral and Brain Sciences. 2004. V. 27. № 4. P. 543–559. https://doi.org/10.1017/S0140525X04000123
Gurven M., Stieglitz J., Hooper P.L., et al. From the womb to the tomb: the role of transfers in shaping the evolved human life history // Experimental gerontology. 2012. V. 47. № 10. P. 807–813. https://doi.org/10.1016/j.exger.2012.05.006
Fehr E., Bernhard H., Rockenbach B. Egalitarianism in young children // Nature. 2008. V. 454. № 7208. P. 1079. https://doi.org/10.1038/nature07155
Henrich J., Ensminger J., McElreath R., et al. Markets, religion, community size, and the evolution of fairness and punishment // Science. 2010. V. 327. P. 1480–1484. https://doi.org/10.1126/science.1182238
Henrich J., Muthukrishna M. The Origins and Psychology of Human Cooperation // Annual Review of Psychology. 2020. https://henrich.fas.harvard.edu/files/henrich/files/henrich_and_muthukrishna_the_origins_and_psychology_of_human_cooperation_final.pdf
Hamann K., Warneken F., Greenberg J.R., et al. Collaboration encourages equal sharing in children but not in chimpanzees // Nature. 2011. V. 476. № 7360. P. 328–331. https://doi.org/10.1038/nature10278
Luo S., Zhang T., Li W., et al. Interactions between oxytocin receptor gene and intergroup relationship on empathic neural responses to others’ pain // Social cognitive and affective neuroscience. 2019. V. 14. № 5. P. 505–517. https://doi.org/10.1093/scan/nsz029
Marlowe F.W. Hadza cooperation // Human Nature. 2009. V. 20. № 4. P. 417–430. https://doi.org/10.1007/s12110-009-9072-6
Rekdal O. Money, milk and sorghum beer: Change and continuity among the Iraqw of Tanzania // Africa. 1996. V. 66. № 3. P. 367–385. https://doi.org/10.2307/1160958
Silk J.B., House B.R. Evolutionary foundations of human prosocial sentiments // Proceedings of the National Academy of Sciences. 2011. V. 108 (Supplement 2). P. 10910–10917. https://doi.org/10.1073/pnas.1100305108
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни