

Доклады Российской академии наук. Науки о жизни, 2020, T. 494, № 1, стр. 481-485
Похож ли жаберный скелет кишечнодышащих (Enteropneusta) на жаберный скелет ланцетника (Cephalochordata)?
О. В. Ежова 1, *, академик РАН В. В. Малахов 1
1 Московский государственный университет
имени М.В. Ломоносова
Москва, Россия
* E-mail: olga.ejova@gmail.com
Поступила в редакцию 23.04.2020
После доработки 30.04.2020
Принята к публикации 30.04.2020
Аннотация
Жаберный скелет кишечнодышащего Saccoglossus mereschkowskii состоит из серии трезубцев. Центральный зубец каждого трезубца оканчивается вилочкой. Самый передний элемент жаберного скелета имеет простую подковообразную форму. В строении жаберного аппарата Cephalochordata найдены гомологи элементов жаберного аппарата Enteropneusta. Строение жаберного скелета Enteropneusta и Cephalochordata может быть выведено из метамерных подковообразных элементов. Сходство в строении жаберного скелета Enteropneusta и Cephalochordata противоречит распространенной гипотезе о перевернутости хордовых.
Жаберные щели представляют собой уникальные органы, свойственные только двум группам животного царства – полухордовым и хордовым. Особенности строения и развития скелета полухордовых имеют большое значение для понимания происхождения хордовых. Однако до сих пор не было проведено сравнение строения жаберного скелета Hemichordata и Chordata. В обзоре Э. Рупперта [1] приведено очень подробное сравнение организации жаберного аппарата полухордовых и низших хордовых животных, затрагивающее форму жаберных щелей и жаберных пор, характер их развития в онтогенезе, поддержку жаберных щелей коллагеновыми скелетными элементами, наличие синаптикул, присутствие целомических полостей в жаберных перегородках, моно- и мультицилиарность выстилающего жаберные щели гастродермиса и расположение в нем железистых клеток, направление движения крови в жаберном аппарате и наличие в последнем подоцитов и нервных элементов. Однако в цитируемом обзоре нет сравнения как таковой формы самих скелетных элементов жаберного аппарата полухордовых и низших хордовых.
Задача настоящей работы состояла в том, чтобы реконструировать организацию жаберного скелета представителя полухордовых – кишечнодышащего Saccoglossus mereschkowskii и попытаться сравнить строение жаберного скелета полухордовых и хордовых.
Взрослые экземпляры S. mereschkowskii были собраны в илистом грунте на глубине 4–10 м в окрестностях Беломорской биологической станции МГУ в Кандалакшском заливе Белого моря. Фрагменты животных были зафиксированы в жидкости Буэна, затем дегидратированы в спиртах восходящей концентрации и заключены в парапластовые блоки. Для последующей 3D-реконструкции была изготовлена серия фронтальных срезов S. mereschkowskii толщиной 7 мкм. Срезы были окрашены гематоксилином и отсняты с помощью фотомикроскопа Zeiss Axioplan 2. Упрощенные схематические рисунки, представленные в работе, сделаны на основе 3D-реконструкции, осуществленной в программе Amira, версия 6.4.0.
Важно отметить во избежание путаницы, что в работе мы употребляем сравнение “подковообразный” только по отношению к скелетным элементам, а “U-образный” – только по отношению к жаберным щелям.
Скелетные элементы, поддерживающие форму жаберных щелей в глотке Saccoglossus mereschkowskii, представляют собой утолщения экстра-клеточного матрикса в складках глоточного гастродермиса. Каждый скелетный элемент, кроме самой перво й, передней пары скелетных элементов, имеет форму трезубца с направленными вниз, к вентральной стороне, зубцами (рис. 1). Первая пара скелетных элементов имеет подковообразную (“двузубую”) форму, причем зубцы имеют различную длину (рис. 2а). На реконструкции хорошо видно, что форма трезубца у всех остальных жаберных скелетных элементов достигается за счет срастания двух таких подковообразных элементов. В результате такого срастания центральный зубец получается двойным, более мощным, длинным и раздвоенным на конце. Боковые, или краевые (маргинальные), зубцы – простые и более короткие (рис. 2а).
Рис. 1.
Реконструированная модель жаберного аппарата кишечнодышащего Saccoglossus mereschkowskii, демонстрирующая организацию жаберного скелета: вжп – вторичная жаберная перегородка; гл – гастродермис глотки; жм – жаберный мешок; жп – жаберная пора; жщ – жаберная щель; пжп – первичная жаберная перегородка; ск – скелетные элементы.

Рис. 2.
Схема строения жаберного скелета Saccoglossus mereschkowskii (а) (наши данные) и Branchiostoma sp. (б) (по [6], синаптикулы не показаны).

Жаберные щели в гастродермисе глотки S. mereschkowskii имеют U-образную форму и отделяются друг от друга так называемыми первичными жаберными перегородками, или “септами” (рис. 1). Их U-образная форма достигается за счет неполных вторичных жаберных перегородок (или язычковых перегородок) (рис. 1). И первичные, и вторичные жаберные перегородки укреплены скелетными элементами.
Центральные зубцы жаберных скелетных элементов (с вилочкой на конце) проходят именно в первичных жаберных перегородках. Краевые же зубцы укрепляют вторичные жаберные перегородки, причем в каждую такую перегородку входит один краевой зубец от предыдущего скелетного элемента и один краевой зубец – от последующего скелетного элемента. Таким образом, каждый скелетный трезубец поддерживает заднюю половинку одной жаберной щели и переднюю половинку соседней жаберной щели (рис. 1).
Простые, несросшиеся подковообразные скелетные элементы первой пары своим задним, более коротким зубцом поддерживают вторичную перегородку первой жаберной щели, а их передний, более длинный зубец просто “заякорен” в складке гастродермиса глотки.
Форма трезубца для жаберных скелетных элементов свойственна всем кишечнодышащим. У представителей семейства Harrimaniidae и у некоторых видов семейства Spengelidae вторичные жаберные перегородки свободно свисают в жаберную щель, а жаберные скелетные элементы имеют форму простого трезубца. У представителей семейства Ptychoderidae отверстия жаберных щелей пересечены поперечными синаптикулами, соединяющими первичные и вторичные жаберные перегородки [2].
У представителей другого класса полухордовых – Pterobranchia – жаберный аппарат очень упрощен по сравнению с кишечнодышащими. У Cephalodiscus имеется единственная пара овальных (лишенных вторичных перегородок и, следовательно, не U-образных) жаберных щелей [3, 4]; у Rhabdopleura жаберные поры вообще отсутствуют [5]. Каких-либо скелетных структур, поддерживающих жаберные щели, у Cephalodiscus нет. Только сами жаберные канальцы, соединяющие полость глотки с внешней средой, выстланы вакуолизированными клетками [3], которые, вероятно, обеспечивают необходимую жесткость стенок овальных жаберных щелей.
Жаберный скелет кишечнодышащих обладает очевидным сходством с жаберным скелетом низших хордовых, а именно Cephalochordata (рис. 2). По своему гистологическому строению и химическому составу жаберные скелетные элементы кишечнодышащих и ланцетника также схожи – в обоих случаях это лишенные клеток утолщения экстраклеточного вещества, состоящие из коллагена и протеогликанов [7, 8]. Жаберный скелет и у кишечнодышащих, и у Cephalochordata образован метамерными подковообразными элементами, срастающимися между собой (рис. 2б). Концы подковообразных элементов при этом в обеих группах направлены к вентральной стороне. У кишечнодышащих подковообразные элементы срастаются попарно, образуя трезубцы. В результате такого срастания имеет место чередование центральных более длинных зубцов с вилочкой на конце и более коротких несросшихся краевых зубцов (рис. 2а). У ланцетника все жаберные скелетные элементы срастаются между собой, образуя общую решетку на протяжении глотки. Однако и у ланцетника имеет место чередование зубцов с вилочкой на конце и простых зубцов без вилочки (рис. 2б). Это дает возможность предположить, что жаберная решетка ланцетника сформировалась из того же варианта, который сохранился у современных кишечнодышащих. Центральные зубцы трезубца кишечнодышащих (с вилочкой на конце) соответствуют зубцам решетки ланцетника с такой же вилочкой на конце. Чередующиеся с ними простые зубцы решетки ланцетника соответствуют несросшимся краевым зубцам жаберного скелета кишечнодышащих (рис. 3). Интересно, что в жаберном скелете кишечнодышащих самый первый элемент сохраняет подковообразную форму. Этот первый подковообразный элемент легко вычленяется и в жаберном скелете ланцетника (рис. 2).
Рис. 3.
Гипотетическая схема эволюции жаберного скелета Enteropneusta и Cephalochordata. Пунктиром показаны жаберные щели.
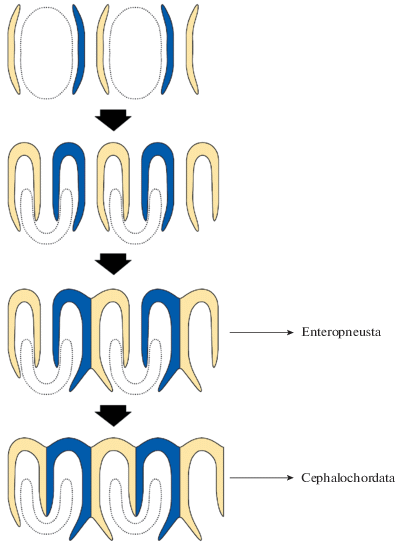
На рис. 3 представлена гипотетическая схема эволюции жаберного скелета Enteropneusta и Cephalochordata.
Жаберные щели кишечнодышащих закладываются как овальные отверстия (у Pterobranchia они такими и остаются). В процессе развития жаберные щели принимают U-образную форму за счет того, что с дорсальной стороны в каждую щель врастает язычок, состоящий из несросшихся краевых зубцов жаберного скелета [9]. У ланцетника жаберные щели также закладываются как овальные отверстия [10]. В процессе развития жаберные щели становятся U-образными за счет того, что с дорсальной стороны в них врастают лишенные вилочки зубцы жаберного скелета. У ланцетника эти зубцы продолжают рост и полностью разделяют U-образную жаберную щель на две отдельных щели.
Приведенные выше соображения представляют интерес в связи с гипотезой перевернутости хордовых. Современный вариант этой гипотезы был предложен одним из авторов этой статьи [11]. Позднее эта гипотеза нашла подкрепление в данных по экспрессии гомеобоксных генов [12]. В настоящее время гипотеза перевернутости хордовых рассматривается как одна из парадигм современной зоологии [13].
Если считать, что хордовые – это перевернутые кишечнодышащие, то свободные концы подковообразных элементов жаберного скелета должны быть направлены в разные стороны. Если у кишечнодышащих они направлены вентрально, то у перевернутых хордовых они должны быть направлены дорсально. То же самое относится и к жаберным щелям. У кишечнодышащих свободные концы U-образных жаберных щелей направлены дорсально. У перевернутых хордовых свободные концы U-образных жаберных щелей должны быть направлены вентрально. Тем не менее, в обеих группах свободные концы подковообразных скелетных элементов направлены вентрально, а свободные концы U-образных жаберных щелей в обеих группах направлены дорсально.
Эти факты противоречат гипотезе перевернутости хордовых. Если пытаться сохранить трактовку хордовых как перевернутых кишечнодышащих, поскольку эта гипотеза хорошо подкреплена современными данными по биологии развития, то придется признать, что эволюционная судьба кишечнодышащих и хордовых разошлась на этапе, когда они имели простые овальные жаберные щели, и формирование жаберного скелета происходило в обеих группах независимо. К этому же выводу приходит в своем обзоре и Э. Рупперт на основании сравнения других деталей строения жаберного аппарата низших вторичноротых [1].
Список литературы
Ruppert E.E. Key characters uniting hemichordates and chordates: homologies or homoplasies? // Canadian Journal of Zoology. 2005. V. 83. № 1. P. 8–23.
Spengel J.W. Die Enteropneusten des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna und Flora des Golfes von Neapel. Herausgegeben von der Zoologischen Station zu Neapel 18. Monograph. Berlin: Verlag von R. Friedländer and Sohn. 1893.
Ridewood W. Pterobranchia: Cephalodiscus. National Antarctic Expedition Reports, Natural History. London: British Museum. 1907.
Schepotieff A. Die Pterobranchier. Die Anatomie von Cephalodiscus dodecalophus // Zool. Jahrb. Abt. Anat. Ontog. Tiere. 1907a. № 24. S. 193–238.
Schepotieff A. Die Pterobranchier. Die Anatomie von Rhabdopleura // Zool. Jahrb. Abt. Anat. Ontog. Tiere. 1907b. № 23. S. 463–534.
Franz V. Morphologie der Akranier // Ergebn. Anat. und Entwickl. Gesch. 1927. V. 27. P. 464–568.
Pardos F., Benito J. Blood vessels and related structures in the gill bars of Glossobalanus minutus (Enteropneusta) // Acta Zool. 1988. V. 69. P. 87–94. https://doi.org/10.1111/j.1463-6395.1988.tb00905.x
Rychel A.L., Smith S.E., Shimamoto H.T., et al. Evolution and Development of the Chordates: Collagen and Pharyngeal Cartilage // Mol. Biol. Evol. 2006. V. 23. № 3. P. 541–549. https://doi.org/10.1093/molbev/msj055
Bateson W. Continued account of the later stages in the development of Balanoglossus kowalevskii, and of the morphology of the Enteropneusta // Quart. Jour. Microsc. Sci. 1886. V. 26. P. 511–533.
Willey A. The later larval development of Amphioxus // Quart. Jour. Microsc. Sci. 1891. V. 32. № 126. P. 183–230.
Малахов В.В. Проблема основного плана строения в различных группах вторичноротых животных // Журнал общей биологии. 1977. Т. 38. № 4. С. 485–499.
Slack J., Holland P., Graham C. The zootype and the phylotypic stage // Nature. 1993. V. 361. P. 490–492. https://doi.org/10.1038/361490a0
Ruppert E.E., Barnes R.D., Fox R.S. Invertebrate Zoology. Belmont: Thomson Brooks/Cole. 2004. V. 27. Hemichordata.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни