

Доклады Российской академии наук. Науки о жизни, 2021, T. 496, № 1, стр. 28-31
КОНЦЕНТРИЧЕСКИЕ ВКЛЮЧЕНИЯ В КИШЕЧНОМ ЭПИТЕЛИИ ЭХИУРИДЫ BONELLIA VIRIDIS: СТРОЕНИЕ И ВОЗМОЖНАЯ ФУНКЦИЯ
П. А. Кузнецов 1, Е. Н. Темерева 1, *
1 Московский государственный университет
имени М.В. Ломоносова
Москва, Россия
* E-mail: temereva@mail.ru
Поступила в редакцию 23.09.2020
После доработки 29.09.2020
Принята к публикации 30.09.2020
Аннотация
Изучение анатомии и тонкого строения эхиурид имеет большое значение для понимания биологии этих животных, ведущих скрытный образ жизни и доминирующих в различных бентосных сообществах. Методами сканирующей и трансмиссионной электронной микроскопии были получены первые данные об организации сифонального отдела средней кишки самок Bonellia viridis. Впервые в клетках средней кишки обнаружены необычные концентрические включения, сходные по ультраструктуре с включениями, описанными у других животных, например, в кишечнике многих видов нематод и в тегументе некоторых цестод. Для этих животных известно, что концентрические включения играют важную роль в связывании химических агентов, присущих окислительно-восстановительным средам. Интересно, что исследованные особи B. viridis были обнаружены на поверхности субстрата, лишенного признаков окислительно-восстановительной среды. Новые результаты свидетельствует о наличии у B. viridis и, возможно, у всех эхиурид, преадаптаций к жизни в окислительно-восстановительных средах. Новые данные о строении и составе концентрических включений прольют свет на их происхождение и функции.
Эхиуриды – это небольшая группа несегментированных морских бентосных червей, которая насчитывает около 165–195 видов [1]. Согласно современным данным молекулярной филогенетики эхиуриды представляют собой одну из групп кольчатых червей (Annelida). Эхиуриды живут скрытно, их тело целиком скрыто в норе, из которой они высовывают только хобот. Такие особенности биологии делают эхиурид крайне труднодоступными для изучения, из-за чего многие системы органов эхиурид остаются практически не исследованными современными методами [2]. Это в полной мере касается и организации пищеварительного тракта, который, как оказалось, демонстрирует существенные вариации анатомической организации у разных видов [3]. Детальное изучение тонкого строения средней кишки позволит лучше понять особенности функционирования этого особого отдела кишечника и прольет свет на биологию этих загадочных животных.
Целью настоящей работы было изучение тонкой организации эпителия сифонального отдела средней кишки самок Bonellia viridis.
Материалом для работы послужили три самки B. viridis Rolando, 1821, собранные на каменистом дне в Марсельском заливе у острова План (Plane) 43°11'09.99'' N – 5°23'29.52'' E, на глубине 25–27 м. Животных собирали на поверхности каменистого субстрата в ходе погружений с легководолазным снаряжением. Животных вскрывали, участки средней кишки фиксировали в 2.5% растворе глютарового альдегида на 0.2M фосфатном буфере. Для изучения методом сканирующей электронной микроскопии (СЭМ) участки средней кишки дегидратировали в этаноле и ацетоне, высушивали в критической точке, наклеивали на столики и напыляли смесью платины и палладия. Тонкая морфология сифонального отдела средней кишки была изучена в сканирующем электронном микроскопе JEOL JSM-6380LA. Для изучения ультратонкой организации кишечного эпителия методом трансмиссионной электронной микроскопии (ТЭМ) участки кишечника были отмыты в буфере, постфиксированы в 1% растворе тетроксида осмия, обезвожены и заключены в смолу Embed-812. Подготовленные блоки были ультратомированы на полутонкие и ультратонкие срезы. Полутонкие срезы были окрашены метиленовым синим, изучены в световой микроскоп Zeiss Axioplan 2 и сфотографированы цифровой камерой. Ультратонкие срезы контрастировали уранил ацетатом и цитратом свинца и изучали на микроскопах JEOL JEM-2100 и JEOL JEM-1011.
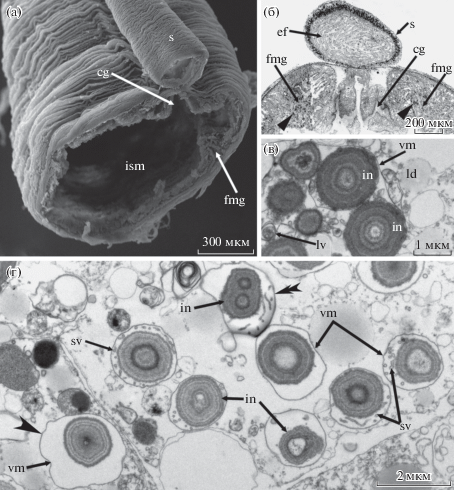
Сифональный отдел средней кишки самок B. viridis характеризуется наличием протяженной коллатеральной трубки – сифона (рис. 1а, 1б). Диаметр сифона не превышает 300 мкм, тогда как диаметр сифонального отдела средней кишки может достигать 4 мм (рис. 1а, 1б). Просвет сифона заполнен тонкими длинными складками эпителия (рис. 1б). Сифональный отдел средней кишки выстлан высоким столбчатым однослойным эпителием, высота клеток которого составляет 60–70 мкм (рис. 1б). Эпителий формирует крупные продольные складки, которые вдаются в просвет кишки. Эпителий сифонального отдела средней кишки не однороден по гистологическому строению. Так, ресничный желобок, проходящий в месте прилежания сифона, образован ресничными клетками, лишенными включений, тогда как клетки остального эпителия сифонального отдела средней кишки несут многочисленные включения (рис. 1б). Как показали ультраструктурные исследования, включения, как правило, занимают апикальную и среднюю части клеток. Каждое включение располагается внутри вакуоли и окружено тонкой мембраной (рис. 1г). Пространство вакуоли между включением и мембраной имеет различный объем, заполнено электронно-светлым материалом и содержит мелкие мембранные пузырьки диаметром около 0.1 мкм (рис. 1г). Мембранных пузырьков может быть довольно много, либо же они могут вовсе отсутствовать. В некоторых включениях в пространстве под мембраной обнаруживаются крупные везикулы, а также небольшие включение и мультивезикулярные тельца. Сами концентрические включения имеют разные размеры: их диаметр колеблется от 1 до 2 мкм. Включения имеют очень характерную правильную концентрическую структуру: в них закономерно чередуются электронно-плотные и электронно-светлые зоны, которые образуют кольца по периметру включения (рис. 1в, г). Число концентрических зон может быть различно: от 2 до 10. Центр включения может быть светлым или же заполнен электронно-плотным содержимым. В некоторых включениях обнаруживаются две ацентрические области (рис. 1г). Как правило, одно концентрическое включение залегает внутри собственного пространства, окруженного мембраной. Однако зачастую можно видеть, что несколько включений формируют крупные скопления, окруженные общей мембраной. При этом индивидуальная мембрана может сохраняться или отсутствовать (рис. 1в). Диаметр таких скоплений может достигать 3–4 мкм.
Рис. 1.
Строение сифонального отдела средней кишки самки Bonellia viridis. (а) – Участок сифонального отдела средней кишки на срезе (СЭМ). (б) – Участок средней кишки с сифоном на поперечном полутонком срезе. Концентрические включения в клетках указаны прямыми наконечниками. (в) – Крупное скопление концентрических включений, окруженных общей мембраной (ТЭМ). (г) – Разные типы концентрических включений: с двумя ацентрическими областями (двойной наконечник), с мембранным пространством, лишенным везикул (вогнутый наконечник) (ТЭМ). Обозначения: (cg) – ресничный желобок, (ef) – складки эпителия сифона, (fmg) – складка эпителия средней кишки, (in) – концентрические включения, (ld) – липидные капли, (lsm) – просвет сифонального отдела средней кишки, (lv) – крупные везикулы, (s) – сифон, (sv) – мелкие мембранные пузырьки, (vm) – мембрана вакуоли.
Анатомия пищеварительного тракта сходна в пределах группы Echiura. За исключением карликовых самцов бонеллид пищеварительная система эхиурид состоит из 3 отделов: передней, средней и задней кишки [2]. Уникальной особенностью строения средней кишки эхиурид является наличие сифона. Его функция до конца непонятна.
Необычной ультраструктурной особенностью клеток средней кишки исследованных нами экземпляров B. viridis является наличие в их цитоплазме специфических концентрических включений. В эпителии кишечника эхиуриды Urechis caupo были описаны клетки, содержащие сульфидокисляющие гранулы, такие же, как и в эпителии стенки тела у этого вида [4]. По мнению авторов работы, сульфидокисляющие гранулы участвуют в детоксификации сульфидов, которые накапливаются в норе животного в бескислородных условиях. Микрорентгеновский анализ этих включений показал, что они состоят из алюминия, хлора, железа, меди и фосфора. Эти вещества представлены в разных пропорциях на протяжении всего пищеварительного тракта. Хлор составляет большую долю, и его концентрация наибольшая в гранулах клеток средней кишки. Железо и алюминий занимают второе место по обилию в составе гранул. Наибольшая концентрация обоих этих элементов отмечена в клетках жевательного желудка [4]. Важно отметить, однако, что на электронограммах, представленных в работах цитированных авторов [4], сульфид-окисляющие гранулы не имеют характерной концентрической структуры.
Ультратонкое строение включений, обнаруженных в средней кишке B. viridis, довольно сильно отличается от такового сульфидокисляющих гранул, описанных Меноном и Арпом у U. caupo [4], однако имеет настолько характерные черты, что сравнимо с подобными включениями, обнаруженными в кишечнике других беспозвоночных. Специфические концентрические включения были найдены в кишечнике у нематод, чей пищеварительный тракт был изучен на ультраструктурном уровне [5]. Рентгеновский анализ концентрических включений нематод Trichuris suis и T. murk показал, что они имеют высокое содержание кальция, а дополнительное присутствие фосфора позволяет предположить, что основным компонентом включения является фосфат кальция. Кроме того, во включениях были найдены следы магния, железа и калия [5]. Функция этих включений неизвестна. Большинство авторов склоняются к мысли, что это экскреты: связанные в физиологически и химически неактивные соединения – продукты метаболизма. Есть, однако, и другое мнение о природе этих необычных включений – фон Бранд и Вайнбах [6] показали, что концентрические включения тегумента ленточных червей способны быстро поглощать и высвобождать фосфат [6]. Фосфат кальция, содержащийся в концентрических включениях, мог бы стать эффективным средством связывания и, в конечном счете, устранения некоторых отходов метаболизма [6].
Возможно, специфические концентрические включения, обнаруженные в клетках средней кишки у B. viridis, выполняют функцию депо ионов для связывания агрессивных химических агентов, которые в избытке присутствуют в глубоких анаэробных слоях ила. Поскольку эхиуриды способны строить довольно глубокие норы, им приходится иметь дело со средами с высокими концентрациями сероводорода, сульфидов, сульфатов и фосфатов. Интересно, что исследованные нами экземпляры B. viridis были собраны на каменистом грунте, лишенном признаков анаэробной среды. Если предположение о функции концентрических включений как депо химических веществ для связывания агрессивных агентов из окружающей среды верно, то B. viridis имеют преадаптации к жизни в окислительно-восстановительных средах и способны использовать включения при попадании в такие условия. То есть при необходимости ионы, содержащиеся в концентрических включениях, поступают в просвет кишки, связываются с агрессивными химическими веществами из окружающей среды, деактивируют их и в форме химически неактивных соединений выводятся из организма.
С другой стороны, сами эти включения могут являться продуктами связывания агрессивных химических агентов и формой их инактивации в клетках кишечного эпителия.
Достоверность обоих этих допущений может быть проверена лишь при наличии дополнительных биохимических, ультраструктурных данных и результатов рентгеновского элементарного анализа. Новые данные по строению и составу этих загадочных включений прольют свет на их происхождение и функции.
Список литературы
Goto R. // Mol. Phylogenet. Evol. 2016. V. 99. P. 247–260.
Pilger J.F. Echiura. Microscopic Anatomy of Invertebrates. Wiley-Liss Inc., New York, 1993. V. 12. P. 185–236.
Temereva E.N., Kuznetsov P.A., Kiseleva E.A. // Rus. J. Mar. Biol. 2017. V. 43. № 6. P. 453–470.
Menon J.G., Arp A.J. // Invertebr. Biol. 1998. V. 117. № 4. P. 307–317.
Jenkins, T., Erasmus, D.A., & Davies, T.W. // Exp. Par. 1977. V. 41. №. 2. P. 464–471.
Von Bhand T., Weinbach E.C. // Zeitschrift fiir Parasitenkunde. 1975. V. 48 P. 53–63.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни