

Доклады Российской академии наук. Науки о жизни, 2021, T. 497, № 1, стр. 148-151
ПЕРИОДИЧЕСКАЯ “СПОНТАННАЯ” АКТИВНОСТЬ ЖИВОТНЫХ ОПРЕДЕЛЯЕТСЯ КВАЗИРИТМИЧЕСКИМ ФАКТОРОМ ВНЕШНЕЙ СРЕДЫ?
М. Е. Диатроптов 1, *, член-корреспондент РАН А. В. Суров 1
1 ФГБУН “Институт проблем экологии и эволюции
им. А.Н. Северцова РАН”
Москва, Россия
* E-mail: diatrom@inbox.ru
Поступила в редакцию 09.11.2020
После доработки 23.11.2020
Принята к публикации 25.11.2020
Аннотация
Проведено исследование динамики пения садовой овсянки (Emberiza hortulana) и разлета на ночлег стаи скворцов (Sturnus vulgaris). Полученные данные были сопоставлены с измеренными параллельно колебаниями температуры тела группы лабораторных мышей. Оправданность таких сопоставлений определялась наличием устойчивой корреляции между показателями ежеминутных изменений температуры тела у лабораторных мышей и зеленушек (Chloris chloris). Было показано, что одновременно с повышением температуры тела у мышей наблюдается увеличение частоты пения садовой овсянки. Более того, моменты устройства на ночлег скворцов совпадают с максимумами в динамике ежеминутных изменений температуры тела мышей. Представленные факты указывают на возможное наличие внешнего синхронизатора “спонтанных” колебаний активности и связанных с ними изменений температуры тела, определяющихся, вероятно, тонусом симпатической нервной системы.
Периодические изменения активности животных разных классов известны и изучаются уже давно [1]. В частности, в динамике интенсивности пения дроздовидной камышевки (Acrocephalus arundinaceus) выявляется периодическое (в диапазоне 4–20 мин) увеличение продолжительности песни с укорочением времени между ними, чередующееся с периодом, когда песни короче, а временные интервалы между ними длиннее [2]. То есть у этого вида птиц наблюдаются колебания мотивационного состояния к пению, вероятно, определяющиеся уровнем возбуждения птицы. В настоящее время нет ответа на вопрос: являются ли такие колебания в активности животных эндогенными или они вызваны действием факторов внешней среды?
Наличие таких ритмов у таксономически далеких организмов указывает на фундаментальные свойства активности ЦНС. Периодическое изменение ЭЭГ реакции на повторяющиеся одинаковые раздражители объясняются изменением спонтанной активности нервной системы [3, 4]. Также принято считать, что незначительные колебания температуры тела, отражающие колебания активности симпатического и парасимпатического отдела вегетативной нервной системы, имеют эндогенную природу [5, 6].
Результаты наших исследований показали, что прием пищи скворцов, находящихся на расстоянии до 100 км друг от друга, происходит преимущественно одновременно, что указывает на синхронность мотивационного поведения к приему пищи [7]. Также было показано, что в суммарных спектрах температуры тела у нескольких видов млекопитающих и птиц, различающихся уровнем обмена веществ, в диапазоне 10–120 мин выявляется одинаковый набор основных гармоник, которые проявляются синхронно у разных особей. Следовательно, период колебаний температуры тела не определяется внутренними особенностями организма и, предположительно, отражает влияние внешнего биотропного фактора среды [8].
Целью данной работы является попытка выявить периодические изменения в динамике пения садовой овсянки и разлета на ночлег стаи скворцов и сопоставить их с измеренными параллельно изменениями температуры тела у лабораторных мышей.
В качестве удобного объекта для наблюдения динамики пения была выбрана садовая овсянка, песня у которой представлена отдельными короткими трелями. Всего было проанализировано 7 ч пения, полученных от трех разных самцов. Из этого времени 3 ч приходилось на вечер, а 4 ч – на утренние часы. Наблюдения были проведены с 9 по 12 июня 2020 г., когда большинство самцов активно не поют и выкармливают птенцов. Проведение наблюдений в такое время минимизировало возможность возбуждения птицы пением самцов конспецификов.
В качестве параметра для оценки ритмичности “принятия решения” была проанализирована динамика устройства на ночлег стаи обыкновенных скворцов. Наблюдения проводили с 1 по 13 мая 2020 г. в период откладки и насиживания яиц, когда самки ночуют в скворечниках. Всего на участке в 30 соток было развешено и заселено 29 скворечников. Вечером в зависимости от погоды за 30–60 мин до устройства на ночлег скворцы усаживались стаей на ближайших самых высоких деревьях, и оттуда, по несколько особей, периодически, на протяжении 20–40 мин залетали в скворечники. Преимуществом такой регистрации являлось то, что “принятие решения” о залете принималось на фоне спокойного вечернего отдыха птицы. Время залета регистрировали с точностью 1 мин. Наиболее длинные и информативные ряды были получены при ясной безветренной погоде, когда продолжительность вечернего отдыха и интервал между залетом на ночлег первых и последних особей максимальны. Вышеперечисленные исследования проводились в Спасском районе Рязанской области.
Параллельная регистрация динамики температуры тела у лабораторных мышей и обыкновенных зеленушек (Chloris chloris) была проведена в период марта-апреля 2020 г. Динамику внутрибрюшинной температуры тела регистрировали с частотой 1 раз в минуту при помощи имплантированных внутрибрюшинно датчиков ДТН3-28 (“ЭМБИ РЕСЕРЧ”, Новосибирск). Внутрибрюшинное внедрение датчиков проводили под золетиловым наркозом (“Virbac Sante Animale”, Франция), вводимым внутримышечно в дозе 7–10 мг/кг. Для лабораторных мышей (n = 7) были созданы условия искусственного постоянного не яркого освещения интенсивностью около 600 лк., а птицы (n = 6) находились при естественном световом режиме. Процедура эксперимента одобрена комиссией по биоэтике ИПЭЭ РАН протокол № 14 от 15.01.2018.
Несмотря на то что анализ индивидуальных ритмов очень важен, для целей настоящей работы мы использовали показатель медианы ежеминутных изменений температуры тела по группе животных, отражающую интегративную реакцию разных особей. Между медианами этих значений по группе мышей и зеленушек показатели корреляции Спирмена в зависимости от календарной даты составляли от 0.08 (р = 0.01) до 0.24 (р = = 0.004). Анализ совпадения экстремумов медианы ежеминутных изменений температуры тела между мышами и зеленушками в диапазоне ритмов с периодом 4–16 мин показал их совпадение с точностью до ±1 мин в 78% случаев. Таким образом, ритмические изменения температуры тела у мышей и птиц наблюдаются синхронно и, вероятно, определяются каким-то квазиритмическим фактором внешней среды.
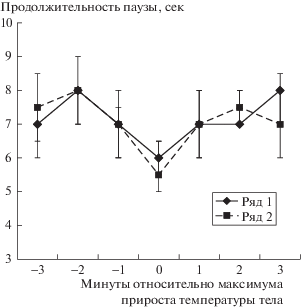
На рис. 1 приведен пример динамики изменения интенсивности пения садовой овсянки и температуры тела в группе мышей: в зависимости от даты исследования преобладали либо 4-минутные изменения, либо 6–12-минутные. Коэффициент корреляции Спирмена между исследуемыми параметрами был статистически значимым. Также сопоставление интенсивности пения и изменений температуры тела было проведено методом наложенных эпох. Было построено распределение показателя продолжительности интервалов между отдельными трелями овсянки относительно экстремумов ежеминутных изменений температуры тела в группе мышей (рис. 2). Оказалось, что в период экстремума температуры тела мышей продолжительность интервалов между отдельными трелями равна 5–6 сек, тогда как в среднем она колебалась в диапазоне 7–8 сек. Показатели в период максимального увеличения температуры тела (в нулевую минуту) и за 2 мин до и после этого события статистически значимо различаются (р = 0.0003 по тесту Манн–Уитни, z = –3.5). Следовательно, существует прямая связь между интенсивностью пения птицы, отражающей ее физиологическое состояние, и колебаниями температуры тела.
Рис. 1.
Примеры динамики интенсивности пения (число трелей в минуту) садовой овсянки (ряд 1) и ежеминутных изменений температуры тела группы мышей (ряд 2). (а) в момент вечернего пения 9 июня (r = 0.41, p = 0.036) и (б) в период утреннего пения 10 июня (r = 0.42, p = 0.02).

Рис. 2.
Распределение методом наложения эпох значений продолжительности интервала между трелями в песне садовой овсянки относительно максимальных ежеминутных изменений температуры тела группы мышей (Утро – ряд 1, Вечер – ряд 2). Представлены медианы и интерквартильный размах.
На рис. 3 представлены примеры параллельной регистрации числа залетевших в скворечник скворцов и ежеминутных изменений температуры мышей: 3 мая преобладала 4-минутная ритмичность, а 5 мая – 6–8-минутная. Коэффициенты корреляции не достигали статистической значимости. Однако распределение всех зарегистрированных залетов в скворечник птиц относительно максимумов медианы ежеминутных изменений температуры тела группы мышей показало их достоверную связь: в период максимума зарегистрировано 92 залета скворцов, а за 2 мин до и после этого только 27–32 случая (р < 0.001 по z тесту для оценки двух выборочных долей, z = 5.99) (рис. 4). Таким образом, установлена связь между общими для птиц и млекопитающих максимальными значениями ежеминутных изменений температуры тела и “принятием решения” о залете на ночлег скворцов.
Рис. 3.
Динамика устройства на ночлег скворцов (ряд 1) и ежеминутные изменения температуры тела группы мышей (ряд 2). (а) 3 мая – пример 4-минутных колебаний (r = 0.17, p = 0.31) ; (б) 5 мая – пример 6–8-минутных колебаний (r = 0.22, p = 0.23).
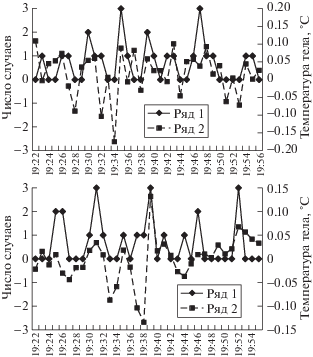
Рис. 4.
Распределение методом наложения эпох числа залетов на ночлег скворцов относительно максимумов в динамике ежеминутных изменений температуры тела группы мышей.

Колебания температуры тела в диапазоне 4–16 мин хорошо коррелируют с изменениями геомагнитного поля, однако существует и ряд других физических факторов, имеющих в своих спектрах те же частоты [8, 9]. Природа внешнего синхронизатора пока остается неизвестной. На основании представленных результатов можно предполагать, что такие формы “спонтанной” активности птиц, как изменения интенсивности пения и “принятие решения” о залете на ночлег в скворечник, не случайны во времени, а связаны с периодическими колебаниями температуры тела в диапазоне 4–16 мин, которые, в свою очередь, определяются активацией симпатической нервной системы и, вероятно, имеют внешние синхронизаторы.
Список литературы
Непомнящих В.А. // Журн. общ. биологии. 2012. Т. 73. № 4. С. 243–252.
Непомнящих В.А., Опаев А.С. // Доклады Академии наук. 2014. Т. 454. № 2. С. 241–243. https://doi.org/10.7868/S0869565214020285
Monto S., Palva S., Voipio J., Palva J.M. // J. Neurosci. 2008. V. 28. 33. P. 8268–8272. https://doi.org/10.1523/JNEUROSCI.1910-08.200
Crevecoeur F., Bollens B., Detrembleur C., Lejeune T.M. // J. Neurosci. Methods. 2010. V. 192. № 3. P. 163–172. https://doi.org/10.1016/j.jneumeth.2010.07.017
Braulke L.J., Heldmaier G. // Cryobiology. 2010. V. 60. № 2. P. 198–203. https://doi.org/10.1016/j.cryobiol.2009.11.001
Blessing W., Ootsuka Y. // Temperature (Austin). 2016. V. 3. № 3. P. 371–383. https://doi.org/10.1080/23328940.2016.1177159
Диатроптов М.Е., Рутовская М.В., Суров А.В. // Доклады Российской Академии наук. Науки о жизни. 2020. Т. 492. № 1. С. 267–271. https://doi.org/10.31857/S2686738920030038
Диатроптов М.Е., Панчелюга В.А., Панчелюга М.С. // Бюлл. эксп. биол. и мед. 2020, Т. 169. № 6. С. 706–711.
Леонович А.С., Мазур В.А. // Солнечно-земная физика. 2005. Вып. 8. С. 99–100.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни