

Доклады Российской академии наук. Науки о жизни, 2021, T. 497, № 1, стр. 143-147
ПРОДУКТИВНОСТЬ И ДИНАМИКА МОРФОЛОГИЧЕСКИХ И ФИЗИОЛОГО-БИОХИМИЧЕСКИХ ПАРАМЕТРОВ КАРТОФЕЛЯ В УСЛОВИЯХ ЗАСУШЛИВОГО КЛИМАТА
О. А. Розенцвет 1, *, Е. С. Богданова 1, В. Н. Нестеров 1, академик РАН С. Н. Шевченко 2, А. Л. Бакунов 3, А. В. Милехин 3, С. Л. Рубцов 3
1 Самарский федеральный исследовательский центр РАН, Институт экологии Волжского бассейна РАН
Тольятти, Россия
2 Самарский федеральный исследовательский центр РАН
Самара, Россия
3 Самарский федеральный исследовательский центр РАН, Самарский научно-исследовательский институт сельского хозяйства им. Н.М. Тулайкова
Безенчук, Россия
* E-mail: olgarozen55@mail.ru
Поступила в редакцию 15.12.2020
После доработки 24.12.2020
Принята к публикации 25.12.2020
Аннотация
Впервые показана взаимосвязь показателей морфологии, физиологии, биохимии и продуктивности растений картофеля на примере среднеспелого сорта (с.) Сиверский и среднераннего с. Третьяковка. Урожайность с. Сиверский составила 512 г/на растение или 24.4 т/га, что оказалось в 1.6 раза выше, чем с. Третьяковка. Надземная биомасса растений картофеля с. Сиверский отличается повышенным содержанием фотосинтетических пигментов, большей изменчивостью показателей белкового и липидного метаболизма, а также более интенсивными процессами окисления и эффективной системой антиокислительной защиты, что может являться залогом его большей урожайности. Многомерным статистическим анализом установлено, что в климатических условиях средней полосы России 2020 г. наибольшая взаимосвязь с продуктивностью выявлена для таких показателей, как число устьиц на единицу площади листа, число стеблей, содержание пигментов, фосфолипидов, нейтральных липидов и водорастворимых белков.
Картофель (Solanum tuberosum L.) является третьим (после риса и пшеницы) и наиболее важным видом сельскохозяйственных культур, употребляемых в пищу человеком [1]. Однако картофель остается уязвимым для биотических и абиотических стрессов [2]. В частности, культивируемый картофель подвержен засухе, и даже кратковременный дефицит воды приводит к снижению урожайности клубней [3, 4]. По имеющимся оценкам одними из общих тенденций изменений биосферы являются аридизация климата и расширение площади пустынных, степных территорий [5]. В течение 21-го века высока вероятность ускорения динамики уже наблюдаемых изменений климата. Ожидаемые перемены могут привести к снижению урожайности картофеля на 26 – 32% в ближайшие десятилетия [6, 7]. В связи с этим необходимо не только создавать новые сорта картофеля, более приспособленные к новым условиям, но и выявлять механизмы толерантности с/хоз. культур с использованием подходов современной фундаментальной науки, что облегчит отбор сортов с нужными свойствами.
Для представителей дикорастущих видов аридной флоры характерна тенденция к снижению запасания углеводов (крахмала) и повышению роли белков и жиров в качестве запасных метаболитов [8]. Адаптивный смысл этой перестройки – более прочное связывание воды белками и липидами и их большая энергоемкость [9]. Это означает, что экологические факторы (например, климатические и биолитогенные) ведут к активизации биохимических адаптивных реакций, которые проявляются в трансформации продуктов метаболизма растений.
Урожайность является важным аспектом в программах селекции [10]. Формирование урожая осуществляется в ходе продукционного процесса – сложной и интегрированной функции растений, основу которой составляют генетически детерминированные процессы роста и развития [11]. Скорость нарастания и размеры листовой поверхности являются одной из важных предпосылок накопления хозяйственно-полезной биомассы и формирования урожая картофеля [12]. Адаптационную способность растений можно оценить по ряду параметров, характеризующих функциональную активность и уровень стресса. Количество пигментов отражает фотосинтетическую способность растений, а их пластичность и адаптивность – устойчивость к факторам среды. Состав клеточных мембран (липидов и белков) характеризует процессы, связанные с переносом веществ через мембраны. Перекисное окисление липидов (ПОЛ) является одним из первых ответов растений на неблагоприятные воздействия, а содержание компонентов, защищающих клетки от свободных радикалов и процессов окисления, свидетельствуют о балансе клеточных процессов катаболизма и анаболизма [13].
Цель работы состояла в выявлении взаимосвязи морфологических, физиологических и биохимических параметров надземной части растений и урожайности клубней картофеля в условиях засушливого климата. Исследования проводили на опытном участке Самарского НИИСХ – филиала СамНЦ РАН в сезон 2020 г. В работе использовали среднеспелый сорт (с.) Сиверский и среднеранний с. Третьяковка. Климатические условия в исследуемый период были крайне неблагоприятными для роста и развития растений картофеля. Так, в периоды завязывания клубней и нарастания их массы отмечено 16.2 мм осадков при среднем многолетнем значении 76 мм. В первой и второй декадах июля осадков не было, а температура воздуха составила в среднем за декаду 25.1 и 24.9°С соответственно. Высадку клубней проводили во второй декаде мая 2020 г. Растения высаживали в четырех повторностях по 50 клубней каждая и выращивали без орошения в однотипных почвенных условиях. Рост растений оценивали по высоте и приросту надземной части 5 случайных растений каждого сорта. Линейные размеры измеряли в 10 боковых долях листа этих растений. Для биохимических анализов из усредненной массы боковых долей листа составляли навески 0.1–0.5 г в трех повторностях для каждого вида анализа. Пробы листьев отбирали в 7.00 и 14.00 в период активного цветения (июль) и увядания ботвы (август). В растениях определяли содержание хлорофилла (Хл) а, Хл b, каротиноидов (Кар), количество мембранных (МБ) и водорастворимых белков (ВБ), мембранных (МЛ) и нейтральных (НЛ) липидов, фосфо- (ФЛ) и гликолипидов (ГЛ), уровень ПОЛ и количество пролина (Прл) и выражали в мг/г сухого веса (с.в.) или ммоль/г с.в. в соответствии с описаниями [14, 15]. Расчеты осуществляли по данным одного биологического опыта. Статистическую обработку проводили в программе Statistica 10 (“StatSoft, Inc.”, США). На рисунках приведены средние арифметические значения (M) со стандартной ошибкой (±SE). Сравнение количественных признаков проводилось на основе t-критерия Стьюдента, результаты статистического анализа считались значимыми при p < 0.05.
Урожайность с. Сиверский составила 512 г/на растение или 24.4 т/га, что оказалось в 1.6 раза выше, чем у с. Третьяковка. По морфологическим признакам установлено, что растения с. Сиверский отличались большим количеством стеблей и большей скоростью роста надземной массы. Боковые доли листьев с. Сиверский имели большие размеры и число устьиц в единице площади листа (табл. 1).
Таблица 1.
Морфологические параметры надземной массы двух сортов картофеля
| Сорт | Целое растение, n = 5 | Боковые доли листа, n = 10 | |||||
|---|---|---|---|---|---|---|---|
| Высота, 50 дней от посадки, см | Высота, 60 дней от посадки, см | При- рост, % | Число стеблей на растение, шт. | Дли- на, см | Шири- на, см | Число устьиц, шт. | |
| Сиверский | 43.4 ± 2.3a | 59.1 ± 6.2a | 36.2 ± 2.9a | 8.6 ± 2.0a | 51.7 ± 1.8a | 30.5 ± 1.0a | 38333 ± 2138a |
| Третьяковка | 52.9 ± 4.1b | 66.6 ± 7.1a | 25.9 ± 2.5b | 4.2 ± 1.5b | 47.7 ± 2.3a | 26.9 ± 1.7b | 28036 ± 1268b |
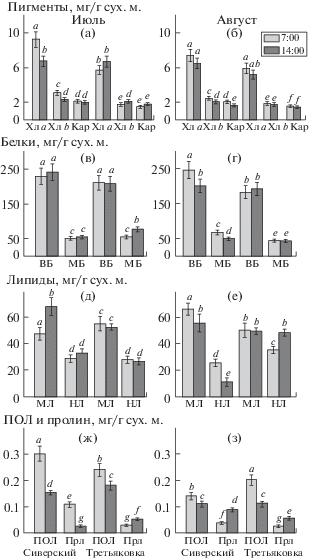
В утреннее время листья с. Сиверский накапливали в 1.3–1.5 раза больше пигментов, чем с. Третьяковка. К середине дня достоверных различий в содержании пигментов в листьях обоих сортов, отобранных в июле, не обнаружено. В период отмирания ботвы у с. Третьяковка содержание пигментов снижалось на 20% (рис. 1 а, б). Общее содержание белка мало различалось между сортами и составляло 295–283 мг/г с.в. в июле и 230–232 мг/г в августе с преобладанием ВБ. Однако в листьях растений с. Сиверский в утреннее время отмечены более высокое содержание белка по сравнению с серединой дня и снижение содержания ВБ по отношению к МБ в августе (рис. 1 в, г). Листья растений с. Сиверский характеризовались также снижением общего содержания липидов в период вегетации за счет снижения количества НЛ, а с. Третьяковка – постоянным количеством суммарных липидов и увеличением НЛ (рис. 1 д, е). Уровень ПОЛ отличался наибольшей изменчивостью значений для обоих сортов. В утреннее время содержание продуктов ПОЛ было выше в 1.5–1.9 раза по сравнению с серединой дня (рис. 1 ж, з). Содержание аминокислоты пролина – активного осмопротектора и антиоксиданта – достоверно выше у с. Сиверский в период отмирания ботвы и в дневное время по сравнению с утренним временем.
Рис. 1.
Биохимические параметры надземной массы картофеля сортов Сиверский и Третьяковка в июле (а, в, д, ж) и августе (б, г, е, з). ВБ – водорастворимые белки, Кар – каротиноиды, МБ – мембранные белки, МЛ – мембранные липиды, НЛ – нейтральные липиды, ПОЛ – перекисное окисление липидов, Прл – пролин, Хл – хлорофилл. М ± SE, n (число определений) = 3. Разными буквами обозначены достоверные различия между вариантами опытов при p < 0.05.
Для анализа взаимосвязей между исследованными показателями морфологии, физиологии, биохимии и продуктивности был использован метод РСА (Principal Components Analysis). Для основного набора характеристик было создано 2-х факторное пространство на основе корреляционной матрицы. Фактор 1 описывает 58% общей вариации, фактор 2 – 33% (рис. 2 а). Круг на графике показывает, насколько хорошо каждая переменная воспроизводится текущим набором факторов – чем она ближе к окружности, тем лучше она воспроизведена. Фактор 1 вносит наибольший вклад в разделение двух сортов картофеля, а фактор 2 – в разделение физиолого-биохимического состояния растений в течение дня (рис. 2 б). Однонаправленность векторов свидетельствует о положительной связи параметров, которая тем сильнее, чем ближе векторы находятся друг к другу. Противоположное направление векторов говорит об отрицательной связи, а угол в 90° – о низкой. Так, вектор “Продуктивность” и векторы “Кар”, “ФЛ”, число устьиц “Nу” и стеблей “Nст” имеют одинаковую направленность и близко расположены друг к другу, т.е. хорошо коррелируют между собой (R > 0.94). Векторы “ПОЛ” и “Прл” также положительно взаимосвязаны (R = 0.89). Векторы “Сух. в.” и “ГЛ” направлены в противоположную сторону от векторов “ПОЛ” и “Прл” (R = –0.90 и R = –0.88 соответственно).
Рис. 2.
Анализ взаимосвязи морфологических, физиологических, биохимических показателей и продуктивности сортов картофеля методом РСА (Principal Components Analysis). Общая вариабельность восемнадцати параметров суммирована с помощью первого и второго основных компонентов (фактор 1 и фактор 2) (2а, б). Для сортов Сиверский (C) и Третьяковка (Т) фактор 1 объяснил 58% общих различий, тогда как фактор 2 – 33%. Обозначения: ВБ – водорастворимые белки, ГЛ – гликолипиды; Кар – каротиноиды, МБ – мембранные белки, МЛ – мембранные липиды, НЛ – нейтральные липиды, H – длина устьиц, ОЛ – общие липиды, ПОЛ – перекисное окисление липидов, Прл – пролин, Продук. – продуктивность, Сух. в. – сухой вес, ФЛ – фосфолипиды, Хл – хлорофилл, Ncт – число стеблей, Nу – число устьиц. 7:00 и 14:00 – время суток.

Проведенные исследования выявили различия морфологических, физиологических и биохимических параметров растений картофеля у высокоурожайного с. Сиверский и низкоурожайного с. Третьяковка. Так, наибольшая взаимосвязь с продуктивностью по данным статистического анализа выявлена для морфологических показателей (число устьиц на единицу площади листа и число стеблей) и биохимических (содержание пигментов, фосфолипидов, нейтральных липидов и водорастворимых белков).
Таким образом, установлено, что как морфологические, так и физиолого-биохимические параметры способны влиять на ход и направленность продукционного процесса и, как следствие, на урожайность определенного сорта.
Список литературы
Deuvax A., Kromann P., Ortiz O. // Potato Res. 2014. V. 57. P. 185–199.
Старовойтов В.И. Современные технологии возделывания картофеля: состояние, перспективы развития. Картофелеводство в регионах России. Актуальные проблемы науки и практики. М.: ВНИИКХ РЦСК, 2006. С. 45–58.
Brown C.R. // Potato Res. 2011. V. 54. P. 287–300.
Obidiegwu J.E., Bryan G.J., Jones H.G., Prashar A. // Front. Plant Sci. 2015. V. 6. P. 542.
Изменение климата, 2014 г.: Обобщающий доклад. Вклад Рабочих групп I, II и III в Пятый оценочный доклад Межправительственной группы экспертов по изменению климата. МГЭИК, Женева, Швейцария, 163 с.
Hijmans R.J. // Am. J. Potato Res. 2003. V. 80. P. 271–279.
Raymundo R., Asseng S., Robertson R., Petsakos A., Hoogenboom G., Quiroz R., Hareau G., Wolf J. // Eur. J. Agron. 2018. V. 100. P. 87–98.
Гамалей Ю.В. // Ботан. журн. 1985. Т. 70. № 10. С. 1302–1313.
Шалпыков К.Т. // Современные проблемы науки и образования. № 5. 2014. http://www.science-education.ru/ru/article/view?id=14499
Keutgen A.J., Keutgen N., Wszelaczyńska E., Pobereżny J., Milczarek D., Tatarowska B., Flis B. // Potato Res. 2020. V. 63. P. 75–95.
Табаленкова Г.Н., Головко Т.К. Продукционный процесс культурных растений в условиях холодного климата. СПб: Наука. 2010. 231 с.
Головко Т.К. Фотосинтез и дыхание в связи с клубнеобразованием у картофеля. Регуляция роста и развития картофеля. М.: Наука. 1990. С. 13–20.
Розенцвет О.А., Нестеров В.Н., Богданова Е.С. // Физиология растений. 2017. Т. 64. № 4. С. 251 – 265.
Rozentsvet O., Nesterov V., Bogdanova E., Kosobryukhov A., Zubova S., Semenova G. // Plant Physiol. Biochem. 2018. V. 129. P. 213–220.
Rozentsvet O.A., Nesterov V.N., Bogdanova E.S. // Phytochemistry. 2014. № 105. P. 37–42.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни