

Доклады Российской академии наук. Науки о жизни, 2021, T. 499, № 1, стр. 385-390
ВЛИЯНИЕ ЭНТОМОПАТОГЕННОГО ГРИБА Beauveria bassiana НА РАЗВИТИЕ ЗАБОЛЕВАНИЙ КОРМОВЫХ БОБОВ (Vicia faba) В ПОЛЕВЫХ УСЛОВИЯХ
Л. Ф. Ашмарина 1, *, Г. Р. Леднёв 2, О. Г. Томилова 3, Т. А. Садохина 1, Д. Ю. Бакшаев 1, М. В. Левченко 2, Н. С. Волкова 2, М. В. Тюрин 3, В. П. Данилов 1, член-корреспондент РАН В. В. Глупов 3
1 Сибирский федеральный научный центр агробиотехнологий Российской академии наук
Новосибирск, Россия
2 Всероссийский научно-исследовательский институт защиты растений
Санкт-Петербург, Россия
3 Институт систематики и экологии животных Сибирское отделение Российской академии наук
Новосибирск, Россия
* E-mail: alf8@yandex.ru
Поступила в редакцию 10.04.2021
После доработки 20.04.2021
Принята к публикации 23.04.2021
Аннотация
Известно, что энтомопатогенные грибы способны выступать в качестве эндофитов. Показано, что гриб Beauveria bassiana (штамм ББК-1) успешно колонизировал растения кормовых бобов Vicia faba в лабораторных и полевых условиях. Реизолят B. bassiana, пропассированный через растения, обладал существенно более высокой антагонистической активностью в отношении фитопатогенных грибов в сравнении с исходным штаммом. Оценка влияния B. bassiana на развитие и распространенность комплекса болезней кормовых бобов в полевых условиях Западной Сибири показала, что предпосевная обработка семян энтомопатогеном уменьшала уровень зараженности семенного материала возбудителями, достоверно снижала развитие и распространенность корневых гнилей. Установлено снижение индекса развития болезней (шоколадная пятнистость, мучнистая роса, фузариоз и прочие пятнистости) в результате применения B. bassiana. Эффективность и пролонгированное действие B. bassiana на растения открывают новые возможности, как в создании биопрепаратов, так и в молекулярно-генетических исследованиях и селекции определенных пар растений и грибов по принципу наибольшего синергизма.
ВВЕДЕНИЕ
С конца 90-х годов ХХ века было показано, что микромицеты, известные как энтомопатогены (представители р. Beauveria, Metarhizium, Isaria, Lecanicillium), способны выступать в качестве эндофитов [1, 2]. К настоящему времени имеется значительное количество сообщений об обнаружении грибов этой группы, как в культурных, так и в дикорастущих растениях [3]. Существует также большое число публикаций по успешной искусственной колонизации энтомопатогенными грибами различных двудольных и однодольных растений [1]. Показано, что присутствие в растениях эндофитных форм энтомопатогенных грибов оказывает негативное влияние на насекомых-фитофагов и фитопатогенов (бактерий и грибов) [4, 3 ], а в ряде случаев приводит к ростстимулирующему эффекту на растениях [2].
Кормовые бобы Vicia faba L. занимают третье место среди возделываемых зернобобовых культур [5]. Потери в производстве бобовых от болезней могут достигать 70–80% [6]. Известно более ста возбудителей болезней этих культур [7]. В условиях континентального климата Западной Сибири бобовые поражаются целым комплексом болезней: корневые гнили (виды р. Fusarium, Alternaria), пятнистости листьев (виды р. Fusarium, Alternaria), мучнистая роса (Erysiphe communis (Wallr.) Grev. f. faba Jacz.), шоколадная пятнистость (Botrytis fabae Sard.) [8]. В связи с этим необходимо проводить поиск и разработку экологически безопасных подходов при выращивании бобовых. Хорошо известно, что энтомопатогенные грибы р. Beauveria могут успешно колонизировать бобовые культуры [9]. Однако в основном результаты были получены только в лабораторных условиях. А в полевых, в том числе в условиях континентального климата России, имеются лишь единичные работы в этом направлении, но по другим культурам [10]. Следует отметить, что в полевых условиях растения обычно поражаются целым комплексом патогенов. В связи с этим целью наших исследований была оценка возможности колонизации кормовых бобов штаммом гриба B. bassiana и его антагонистическая активность к фитопатогенам в лабораторных условиях, а также его влияния на комплекс основных болезней в агроценозе данной культуры в Западной Сибири.
МАТЕРИАЛЫ И МЕТОДЫ
Объектом исследований являлся штамм гриба Beauveria bassiana (Bals.-Criv.) Vuill. (ББК-1), изолированный из трупа итальянского пруса Calliptamus italicus L., собранного в Карасукском р-не Новосибирской обл. из коллекции ФБГНУ ВИЗР. Для оценки антагонистической активности был также взят реизолят этого штамма (ББК-1(Re)), пропассированный через растения бобов. Видовая и внутривидовая идентификация B. bassiana проводилась по межгенному локусу B (Bloc). Для лабораторных опытов гриб выращивали в чашках Петри на агаризированной среде Сабуро. Конидиальную массу для полевого эксперимента нарабатывали методом двухфазного культивирования (первая фаза в глубинной культуре на среде Сабуро, вторая – поверхностно на лущеном зерне ячменя). Титр – 5 × 109 конидий/г.
В опытах использовали семена бобов (V. faba) сорт “Русский черный” (лаборатория) и “Сибирские” (полевые условия).
В лабораторных условиях растения выращивали на стерильной почвенно-песчаной смеси (соотношение 1:1). Инокуляцию проводили методом замачивания семян в суспензии конидий, пролива почвы (5 мл/растение) и опрыскивания вегетирующих растений (до полного его смачивания) по стандартным методикам [11]. Титр – 5 × 107 конидий/мл. Оценку эндофитной колонизации растений (в ходе лабораторного и полевого экспериментов) и выделение реизолятов (лабораторный опыт) проводили согласно [12]. Количественную оценку КОЕ в почве ризосферы (полевой опыт) проводили по методикам, описанным ранее [10].
Оценку антагонистической активности штамма ББК-1 B. bassiana и реизолята ББК-1(Re) проводили in vitro в отношении грибных фитопатогенов из р. Fusarium oxysporum (штамм 4.5), F. solani (4.9), Rhizoctonia sp. (6.2), Bipolaris sorokiniana (2.7) и Botrytis cinerea (3.4) (из рабочей коллекции ВИЗР) по методике совместного культивирования [13]. Ингибирующую активность (ИА) B. bassiana оценивали по изменению диаметра колоний фитопатогена в присутствии B. bassiana в сравнении с контролем [14].
Полевые исследования проводили в 2020 г. на стационаре Сибирского института кормов СФНЦА РАН (Новосибирская область, 54°55 N, 82°56′Е). Тип почвы – чернозем выщелоченный, среднемощный, среднесуглинистый, содержание органического углерода в почве 3.48%, pH – 5.3. Семена кормовых бобов инокулировали суспензией конидий B. bassiana с титром 5 × 107 конидий/мл из расчета 2.5 л на 20 кг семян. На пятый день после обработки был проведен их микологический анализ по стандартным методикам. Визуальные учеты поражения растений различными заболеваниями проводили в поле на 28-й (корневые гнили) и 80-й (листостеблевые болезни) день после посева. Через 28 дней после посева отбирали растения для микологического анализа на пораженность возбудителями корневых гнилей.
Нормальность распределения данных проверяли с помощью W критерия Шапиро–Уилка. Нормально распределенные данные анализировали с помощью t-критерия Стьюдента. Ненормально распределенные данные были проанализированы с помощью теста Манна–Уитни. Для сравнения доли колонизации семян и растений грибами использовали точный критерий Фишера.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В лабораторных условиях при всех трех способах инокуляции бобов грибом B. bassiana уровень колонизации был достаточно высок – 67, 78 и 78% соответственно. Статистически достоверных различий между вариантами не было (p ≥ 0.3). Анализ уровня локализации гриба при разных способах колонизации показал значительные различия по данному показателю. При обработке семян гриб был зафиксирован в корнях и стеблях растений (58 и 42% соответственно). При проливе почвы он был локализован во всех проанализированных частях растений, хотя на листья приходилось лишь 8%. При обработке вегетирующих растений гриб был зафиксирован преимущественно в стеблях и листьях (56 и 40%). Для дальнейших полевых испытаний была выбрана инокуляция семян водной суспензией конидий как более технологичный и менее затратный способ.
Оценка антагонистической активности гриба B. bassiana (ББК-1) in vitro показала, что выбранный для полевого эксперимента штамм способен ингибировать рост ряда почвенных фитопатогенов (F. oxysporum (Schleht.) Snyd. et Hans, F. solani (Mart) App. et Wr., Rhizoctonia spp., B. sorokiniana (Sacc.) Shoemaker, B. cinerea Pers.). ИА штамма ББК-1 составила 22.6–44.4% в зависимости от вида фитопатогена (рис. 1). Антагонистические свойства реизолята, полученного из растений бобов, были достоверно выше в отношении всех оцениваемых культур фитопатогенов (t-тест, p ≤ 0.01), за исключением Rhizoctonia sp.
Рис. 1.
Ингибирующая активность штаммов B. bassiana в отношении фитопатогенов (15-е сутки совместного роста). Разными буквами отмечено наличие достоверных различий в сравнении с исходным штаммом (t-тест, p < 0.05).

Анализ подземных частей растений в полевом эксперименте на присутствие в них B. bassiana показал относительно невысокий уровень эндофитной активности гриба. В фазу стеблевания (28 дней после посева) было колонизировано только 32% растений, а в период цветения (62 дня после посева) лишь 14% (рис 2а).
Рис. 2.
Колонизация растений бобов и ризосферной почвы B. bassiana. Разными буквами отмечено наличие достоверных различий в сравнении с исходным штаммом (Манна–Уитни тест, p < 0.05).
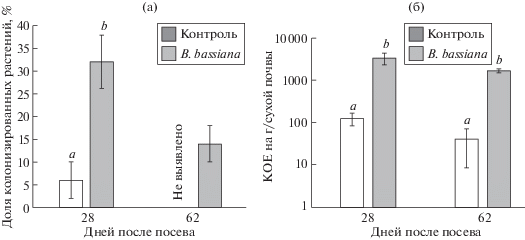
При этом в почве ризозосферы гриб присутствовал в достаточно высокой плотности (в течение двух месяцев после инокуляции выше 1 × × 104 КОЕ/г сухой почвы) (рис. 2б).
Инокулированные для данного эксперимента семена бобов B. bassiana изначально были довольно сильно инфицированы возбудителями корневых гнилей: Alternaria –57.0%, Fusarium – 8.0%.
Анализ зараженности посевного материала показал, что обработка семян бобов конидиями B. bassiana угнетала развитие патогенов (табл. 1).
Таблица 1.
Влияние предпосевной обработки семян бобов конидиями B. bassiana на их зараженность фитопатогенами (среднее по повторности ± ошибка ср.)
| Вариант | Доля семян, заселенных грибными патогенами, % | ||||
|---|---|---|---|---|---|
| Alternaria | Fusarium | Cladosporium | Penicillium | Aspergillus | |
| Контроль | 60.71 ± 5.35 | 8.14 ± 3.76 | 5.43 ± 3.18 | 1.43 ± 1.43 | 16.29 ± 5.74 |
| B. bassiana | 21.29 ± 5.46* | 2.86 ± 1.91 | 2.43 ± 1.65 | 4.29 ± 4.29 | 9.71 ± 5.70 |
Биологическая эффективность обработки в отношении грибов р. Alternaria и Fusarium на семенах составила 64.9%. Существенный уровень достоверности получен в снижении зараженности семян только грибами р. Alternaria (точный тест Фишера, p < 0.0001).
Установлено, что обработка семян B. bassiana приводит к снижению развития корневых гнилей на ранних стадиях роста растений (4 нед. после посадки) (рис. 3).
Рис. 3.
Развитие и распространенность корневых гнилей подземных органов бобов (среднее по повторности ± ошибка ср.). * – наличие достоверных различий в сравнении с контрольным вариантом (Манна–Уитни тест, p < 0.05).

У растений, инокулированных B. bassiana, индекс развития болезни (ИРБ) был в 2.37 раза ниже в сравнении с контролем (тест Манна–Уитни, p = 0.01). Статистически значимые различия наблюдались также и при оценке распространенности корневых гнилей на подземных органах бобов (p = 0.01). Биологическая эффективность обработки семян B. bassiana в снижении развития инфекции составила 57.78%.
Из подземных органов растений бобов были изолированы преимущественно грибы р. Fusarium, Alternaria, Cladosporium (табл. 2), что подтверждает их участие в этиологии заболеваний бобов.
Таблица 2.
Анализ подземных органов бобов на зараженность возбудителями корневых гнилей (среднее по повторности ± ошибка ср.)
| Вариант | Количество колоний в чашке Петри | ||||
|---|---|---|---|---|---|
| Fusarium | Alternaria | Cladosporium | Mucor | Прочие | |
| Контроль | 8.00 ± 0.55 | 2.40 ± 0.81 | 1.00 ± 0.32 | – | 0.40 ± 0.25 |
| B. bassiana | 6.80 ± 0.66 | 0.60 ± 0.25* | 0.40 ± 0.25 | 0.40 ± 0.25 | – |
Обработка семян B. bassiana оказывала существенное влияние только на инфицирование видами р. Alternaria, вызывая 4-кратное снижение зараженности (точный тест Фишера, p = 0.01). Также отмечено несущественное снижение численности грибов р. Cladosporium и Fusarium (2.5 и 1.18 раза соответственно). Таким образом, не- смотря на изначальную зараженность семян и инфицирование корневой системы почвенными фитопатогенами, обработка семенного материала B. bassiana оказала значительный защитный эффект.
Выявлено существенное влияние B. bassiana на проявление различных листовых болезней, максимальное развитие которых зафиксировано в фазу молочной спелости (рис. 4).
Рис. 4.
Интенсивность развития листовых болезней бобов в фазу молочной спелости бобов (среднее значение по растению). Разными буквами отмечено наличие достоверных различий в сравнении с контрольным вариантом (Манна–Уитни тест, p < 0.05).
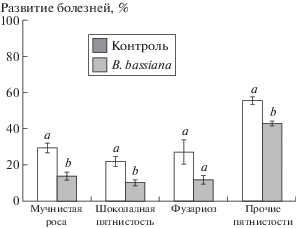
ИРБ мучнистой росы в контроле в 2.1 раза выше, чем в опытном варианте (Манна–Уитни тест, p = 0.0001). Для шоколадной пятнистости при использовании B. bassiana также отмечено статистически значимое снижение развития болезни (в 2.2 раза) (p = 0.0005). Наблюдения за развитием фузариоза показали, что снижение ИРБ составило 2.3 раза, хотя и не являлось статистически значимым (p = 0.28). Статистически значимые различия между контролем и вариантом с обработкой B. bassiаna выявлены по развитию листовых пятнистостей, вызываемых комплексом грибов из рода Alternaria, Cladosporium и Fusarium (p < 0.0001).
В результате проведенных исследований в континентальных условиях Западной Сибири мы впервые установили, что обработка семян кормовых бобов V. faba грибом B. bassiana, выращенных в полевых условиях, повышает устойчивость растений к комплексу заболеваний. Растения, после обработки B. bassiana, в меньшей степени поражались корневыми гнилями, мучнистой росой, шоколадной и другими пятнистостями. Этот эффект мы наблюдали в течение всего периода вегетации бобов.
Аналогичные исследования были проведены на различных видах других растений, но в основном в лабораторных условиях, где не регистрировался комплекс заболеваний [15]. Следует также отметить, что эндофитные взаимоотношения между грибом B. bassiana и растениями, в данном случае с бобами, могут носить факультативно-мутуалистический характер. С одной стороны, снижается повреждаемость растений насекомыми и фитопатогенами [16], с другой, энтомопатогенные грибы получают дополнительный источник питательных веществ и, за счет локализации в прикорневой зоне (ризосфере) или в тканях растений, могут длительно персистировать в ценозах [17].
Стоит отметить, что энтомопатогенные грибы могут подавлять развитие фитопатогенов в растениях-хозяевах в результате конкуренции и микопаразитизма (своеобразный антибиоз) [4], за счет синтеза первичных и вторичных метаболитов, ферментов [14]. Хорошо известно, что ряд вторичных метаболитов B. bassiana, в частности ооспорейн и боверицин, обладают ярко выраженной фунгицидной активностью [18, 19].
Ранее было показано, что данный гриб экспрессирует антимикробный пептид (BbAFP1), локализирующийся в клеточной стенке конидий, затем пептид высвобождается наружу и подавляет рост конкурирующих грибов, в частности Alternaria brassicae [20]. Второй механизм подавления фитопатогенов может быть основан на индуцировании системной резистентности с одновременной стимуляцией роста растений [4, 21]. Можно предположить, что обработка растений, в данном случае семян, энтомопатогенным грибом запускает несколько физиологических механизмов, приводящих к повышенной устойчивости против комплекса патогенов в течение всего периода вегетации. Эффективность и пролонгированное действие B. bassiana на растения открывают новые возможности, как в создании биопрепаратов, так и в молекулярно-генетических исследованиях и селекции определенных пар растений и грибов по принципу наибольшего синергизма.
Список литературы
Vega F.E. Insect pathology and fungal endophytes // J. Invertebr. Pathol. 2008. V. 98. № 3. P. 277–279.
Hu S., Bidochka M.J. Root colonization by endophytic insect-pathogenic fungi // J. Appl. Microbiol. 2021. V. 130. № 2. P. 570–581.
Vega F.E. The use of fungal entomopathogens as endophytes in biological control: a review // Mycologia. 2018. V. 110. № 1, P. 4–30.
Ownley B., Gwinn K., Vega F. Endophytic fungal entomopathogens with activity against plant pathogens: ecology and evolution // BioControl. 2010. V. 55. P. 113–128.
Sing A.K., Bharati R., Pedapati A. An assessment of faba bean (Vicia faba L.) current status and future prospect // African J. of Agricultural Research. 2013. V. 8. № 50. P. 6634–6641.
Horoszkiewicz-Janka J., Jajor E., Korbas M. Potential risk of infection of pathogenic fungi to legumes (Fabales) and possibilities of their control // Prog. Plant Protect. 2013. V. 53. P. 762–767.
Hebblethwaite P.D. The Faba Bean // Butterworths, London, U.K., 1983.
Ашмарина Л.Ф., Горобей И.М., Коняева Н.М., и др. Атлас болезней кормовых культур в Западной Сибири. Новосибирск, 2010.
Akutse K.S., Maniania N.K., Fiaboe K.K., et al. Endophytic colonization of Vicia faba and Phaseolus vulgaris (Fabaceae) by fungal pathogens and their ef ects on the life – history parameters of Liriomyza huidobrensis (Diptera: Agromyzidae) // Fungal Ecology. 2013. V. 6. P. 293–301.
Tomilova O.G., Shaldyaeva E.M., Kryukova N.A., et al. Entomopathogenic fungi decrease Rhizoctonia disease in potato in field conditions // Peer J. 2020. V. 8. P. e9895.
Gurulingappa P., Sword G.A., Murdoch G., et al. Colonization of crop plants by fungal entomopathogens and their effects on two insect pests when in planta // Biol. Control. 2010. V. 55. P. 34–41.
Parsa S., Ortiz V., Vega F.E. Establishment of fungal entomopathogens as endophytes: towards endophytic biological control // J. Vis. Exp. 2013. V. 11. № 74. P. 50360.
Sobowale A.A., Odebode A.C., Cardwell K.F., et al. Antagonistic potential of Trichoderma longibrachiatum and T. hamatum resident on maize (Zea mays) plant against Fusarium verticillioides (Nirenberg) isolated from rotting maize stem // Archives of Phytopathology and Plant Protection. 2010. V. 43. P. 744–753.
Barra-Bucarei L., Iglesias A.F., González M.G., et al. Antifungal activity of Beauveria bassiana endophyte against Botrytis cinerea in two solanaceae crops // Microorganisms. 2020. V. 8. № 1. P. 65.
Jensen R.E., Enkegaard A., Steenberg T. Increased fecundity of Aphis fabae on Vicia faba plants following seed or leaf inoculation with the entomopathogenic fungus Beauveria bassiana // PLoS One. 2019. V. 14. № 10. P. e0223616.
White J.F., Belanger F., Meyer W., et al. Clavicipitalean fungal epibionts and endophytes-development of symbiotic interactions with plants // Symbiosis. 2002. V. 33. № 3. P. 201–213.
Berg G., Smalla K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere // FEMS Microbiol Ecol. 2009. V. 68. № 1. P. 1–13.
Wang Q., Xu L. Beauvericin, a bioactive compound produced by fungi: a short review // Molecules. 2012. V. 17. № 3. P. 2367–2377.
Nagaoka T., Nakata K., Kouno K., et al. Antifungal Activity of Oosporein from an Antagonistic Fungus against Phytophthora infestans // Zeitschrift fur Naturforschung. 2004. V. 59. № 3–4. P. 302–304.
Tong S., Li M., Keyhani N.O., Liu Y., et al. Characterization of a fungal competition factor: production of a conidial cell-wall associated antifungal peptide // PLOS Pathogens. 2020. V. 16. № 4. P. e1008518.
Raad M., Glare T.R., Brochero H.L., et al. Transcriptional reprogramming of Arabidopsis thaliana defence pathways by the entomopathogen Beauveria bassiana correlates with resistance against a fungal pathogen but not against insects hormones, plant–microbe interaction, Plutella xylostella, Myzus persicae, Sclerotinia sclerotiorum // Frontiers in Microbiology. 2019. V. 10. P. 615.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни