

Доклады Российской академии наук. Науки о жизни, 2021, T. 499, № 1, стр. 381-384
ИССЛЕДОВАНИЕ СПОСОБНОСТИ К ГОМОДИМЕРИЗАЦИИ N-КОНЦЕВЫХ ДОМЕНОВ БЕЛКОВ C КЛАСТЕРАМИ ЦИНКОВЫХ ПАЛЬЦЕВ ЧЕЛОВЕКА
Д. В. Фурсенко 1, академик РАН П. Г. Георгиев 1, А. Н. Бончук 1, *
1 Федеральное государственное бюджетное учреждение науки Институт биологии гена Российской академии наук (ИБГ РАН)
Москва, Россия
* E-mail: bonchuk_a@genebiology.ru
Поступила в редакцию 31.03.2021
После доработки 20.04.2021
Принята к публикации 22.04.2021
Аннотация
CTCF принадлежит к большому семейству транскрипционных факторов, имеющих кластеры доменов цинковых пальцев С2Н2 типа (С2Н2 белки), и является основным архитектурным белком млекопитающих. На N-конце CTCF человека находится гомодимеризующийся неструктурированный домен, который может участвовать в организации дистанционных взаимодействий. Для исследования существования аналогичных N-концевых доменов у других С2Н2 белков человека была использована дрожжевая двугибридная система. В общей сложности способность неструктурированных N-концевых доменов к гомодимеризации была исследована для шести С2Н2 белков человека, имеющих сходный с CTCF профиль экспрессии. Полученные данные свидетельствуют об отсутствии способности к гомодимеризации у выбранных участков. С другой стороны, три С2Н2 белка, содержащие на N-конце структурированный DUF3669 домен в дрожжевой двугибридной системе, демонстрируют способность как к гомо-, так и к гетеродимеризации.
СTCF, наиболее изученный архитектурный белок млекопитающих, состоит из неструктурированных концевых областей и расположенного в центральной части кластера из 11 цинковых пальцев С2Н2 типа (С2Н2 домены) [1]. В составе CTCF человека 3–7 С2Н2 домены отвечают за специфичное связывание с консенсусной последовательностью длиной 15 п.н. [2]. Кластер С2Н2 доменов – единственная консервативная часть белка СTCF, имеющая высокий уровень гомологии у большей части позвоночных, насекомых и некоторых нематод [3]. Белки с кластерами из 5 и более С2Н2 доменов (C2H2 белки) обладают способностью к специфическому узнаванию протяженных последовательностей ДНК и у разных таксонов составляют значительную часть ДНК связывающих транскрипционных факторов, функции которых остаются до настоящего времени слабо исследованными [1, 4].
Среди порядка 170 C2H2 белков дрозофилы многие выполняют архитектурную функцию, поддерживая дистанционные взаимодействия. У большинства, помимо кластера C2H2 доменов, на N-конце имеются структурированные домены, способные к специфичной гомодимеризации [5]. Было показано, что гомодимеризация N-доменов у таких белков необходима для поддержания дистанционных взаимодействий [4, 6]. Мы предполагаем, что доля C2H2 белков млекопитающих, выполняющих архитектурную функцию, гораздо больше, чем принято считать сейчас, и что механизм поддержания дистанционных взаимодействий принципиально не отличается между млекопитающими и насекомыми. Однако только сравнительно небольшая часть С2Н2 белков млекопитающих содержит структурированные N-концевые домены SCAN или ВТВ, способные к гомодимеризации [4]. ВТВ домены формируют преимущественно стабильные гомодимеры [7], в то время как для SCAN доменов была показана способность к гомодимеризации и выборочной гетеродимеризации [8]. Для еще одного малоизученного домена, DUF3669, была выборочно показана способность к гомо- и гетеродимеризации [9].
Отсутствие охарактеризованных димеризующих N-концевых доменов у C2H2 белков млекопитающих можно объяснить способностью неструктурированных доменов к олигомеризации, как было нами показано для основного архитектурного белка человека СTCF и его ортологов [10]. С целью проверки этого предположения были выбраны шесть С2Н2 белков, которые имеют паттерн экспрессии, похожий на таковой для белка CTCF (рис. 1). Также в анализ были взяты три С2Н2 белка с N-концевым DUF3669 доменом, для более полной характеристики димеризующей способности данного домена (рис. 2а).
Рис. 1.
Исследование способности к гомодимеризации N-концевых участков C2H2-содержащих белков человека в дрожжевой двугибридной системе. Для проверки способности к гомодимеризации наблюдали способность к росту на селективной среде дрожжей, коэкспрессирующих N-концевой участок, соединенный с ДНК-связывающим доменом GAL4 (BD) или с активационным доменом GAL4 (AD). На схеме доменной структуры белков исследованный участок обозначен толстой линией, ниже указаны соответствующие номера аминокислотных остатков. Ниже представлены результаты дрожжевой двугибридной системы, где + и – означают наличие или отсутствие взаимодействия между соответствующими конструкциями. В качестве положительного контроля использовалась способность к гомодимеризации N-конца белка CTCF человека (1-264 а.к.), а в качестве отрицательного контроля – тестирование на наличие взаимодействия только с активационным (AD) или ДНК-связывающим (BD) доменом белка GAL4. KRAB – Krüppel associated box домен, ZnF C2H2 – домен цинковых пальцев C2H2-типа.
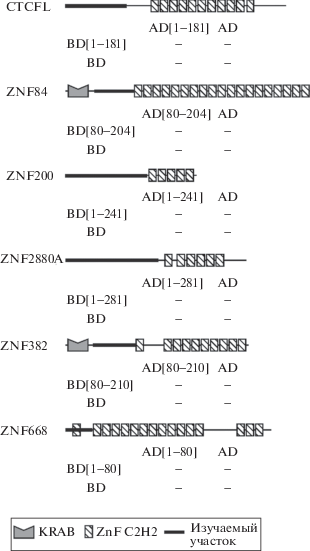
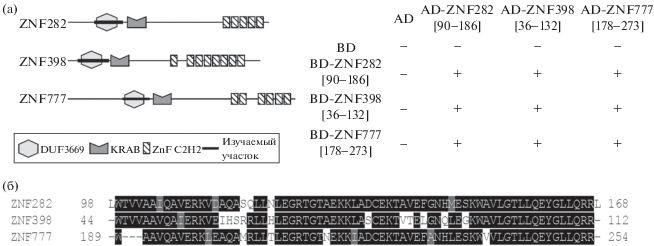
Рис. 2.
(а) Доменные структуры DUF3669-содержащих белков (слева) и результаты дрожжевой двугибридной системы (справа). Результаты представлены аналогично рис. 1. AD – активационный домен, BD – ДНК-связывающий домен. (б) Выравнивание аминокислотных последовательностей доменов DUF3669 белков ZNF282, ZNF398, ZNF777 человека. DUF3669 – домен с неизвестной функцией 3669, KRAB – Krüppel associated box домен, ZnF C2H2 – домен цинковых пальцев C2H2-типа.
Исследование способности N-концевых доменов к гомодимеризации было проведено с использованием дрожжевой двугибридной системы. Ранее данный метод позволил идентифицировать способность к гомодимеризации неструктурированного N-концевого участка белка СTCF разных видов животных [10]. В отличие от неструктурированного N-концевого домена CTCF человека, димеризация которого служила положительным контролем в данном эксперименте, не было обнаружено взаимодействия N-концевых участков белков ZNF84, ZNF200, ZNF280A, ZNF382, ZNF668, а также CTCFL, паралога CTCF (рис. 1). Последний результат представляет особый интерес, поскольку CTCFL имеет значительное сходство с CTCF в структуре цинковых пальцев и может связывать его сайты, однако экспрессируется преимущественно в семенниках млекопитающих [11]. Предполагается, что CTCFL участвует в регуляции архитектурных функций CTCF, подменяя его в определенных случаях на сайтах и тем самым нарушая формирование хроматиновых петель.
У человека предсказано 7 белков, имеющих DUF3669 домен, однако только у трех из них (ZNF282, ZNF398 и ZNF777) на C-конце есть кластер из 5 и более цинковых пальцев C2H2-типа (рис. 2а, слева). Данные белки находятся в одном кластере в геноме человека и многих других видов животных, а аминокислотная последовательность самого домена DUF3669 этих белков у человека имеет высокую степень гомологии (рис. 2б). Помимо высокой степени сходства, в дрожжевой двугибридной системе домен DUF3669 у всех взятых в работу С2Н2 белков способен не только к гомо-, но и к гетеродимеризации (рис. 2а, справа).
Наличие способности к специфическому связыванию длинных мотивов ДНК и к гомодимеризации являются отличительными свойствами архитектурных белков дрозофилы. У млекопитающих из ~800 С2Н2 белков лишь 84 имеют димеризующие SCAN или BTB домены [1]. В нашей работе мы подтвердили и показали, что малоизученный N-концевой домен DUF3669 так же способен к образованию гомо- и гетеро- димеров у трех C2H2 белков человека. Согласно модели, разработанной на дрозофиле, предполагается, что большая часть С2Н2 белков участвует в организации архитектуры хромосом. Преимущественная гомодимеризация N-концевых доменов С2Н2 белков дрозофилы является ключевой в организации специфичных дистанционных взаимодействий [12]. Однако среди С2Н2 белков человека димеризация N-концевых доменов имеет незначительное распространение по сравнению с аналогичными белками дрозофилы [4]. Также результаты нашей работы не подтверждают предположения, что среди C2H2 белков человека широко распространена гомодимеризация за счет неструктурированного N-концевого участка. Стоит отметить, что у наиболее исследованного архитектурного/инсуляторного белка дрозофилы Su(Hw) на N-конце также не было обнаружено домена, способного к гомодимеризации [13]. Возможным, но слабо исследованным, механизмом гомодимеризации для таких белков может является участие самих С2Н2 доменов в этом процессе. Так, было показано, что белок YY1 человека гомодимеризуется с участием С2Н2 доменов [14]. Таким образом, требуется дальнейшее изучение механизмов формирования гомодимеров у C2H2 белков человека.
Список литературы
Максименко О.Г., Фурсенко Д.В., Белова Е.В. и др. // Acta Naturae. 2021. V. 13. № 1. P. 80–95.
Hashimoto H., Wang D., Horton J.R., et al. // Mol. Cell. 2017. V. 66. № 5. P. 711–720.
Heger P., Marin B., Bartkuhn M., et al. // Proc. Natl. Acad. Sci. U. S. A. 2012. V. 109. № 43. P. 17507–17512.
Федотова A.A., Бончук A.Н., Могила В.А., и др. // Acta Naturae. 2017. V. 9. P. 47–58.
Bonchuk A., Boyko K., Fedotova A., et al. // Nucleic Acids Res. 2021. V. 49. № 4. P. 2375–2389.
Zolotarev N., Fedotova A., Kyrchanova O., et al. // Nucleic Acids Res. 2016. V. 44. № 15. P. 7228–7241.
Stogios P.J., Downs G.S., Jauhal J.J.S., et al. // Genome Biol. 2005. V. 6. № 10. P. 1–18.
Liang Y., Huimei Hong F., Ganesan P., et al. // Nucleic Acids Res. 2012. V. 40. № 17. P. 8721–8732.
Al Chiblak M., Steinbeck F., Thiesen H.J., et al. // BMC Mol. Cell Biol. 2019. V. 20. № 1. P. 1–23.
Bonchuk A., Kamalyan S., Mariasina S., et al. // Sci. Rep. 2020. V. 10. № 1. P. 2677.
Loukinov D.I., Pugacheva E., Vatolin S., et al. // Proc. Natl. Acad. Sci. U. S. A. 2002. V. 99. № 10. P. 6806–6811.
Maksimenko O., Kyrchanova O., Klimenko N., et al. // Biochim. Biophys. Acta. Gene Regul. Mech. 2020. V. 1863. № 1. P. 1863.
Melnikova L., Kostyuchenko M., Molodina V., et al. // Open Biol. 2017. V. 7. № 10. P. 170150.
Li L., Williams P., Ren W., et al. // Nat. Chem. Biol. 2021. V. 17. № 2. P. 161–168.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни