

Доклады Российской академии наук. Науки о жизни, 2021, T. 499, № 1, стр. 291-295
НОВЫЕ ДАННЫЕ О РАСПРОСТРАНЕНИИ МАЛОГО ПЕЩЕРНОГО МЕДВЕДЯ (Ursus ex gr. savini-rossicus) НА УРАЛЕ
Д. О. Гимранов 1, *, М. В. Павлова 2, О. Г. Нанова 3, П. А. Косинцев 1, член-корреспондент РАН Н. Г. Смирнов 1
1 Институт экологии растений и животных УрО РАН
Екатеринбург, Россия
2 Уральский государственный педагогический университет
Екатеринбург, Россия
3 Зоологический музей МГУ им. М.В. Ломоносова
Москва, Россия
* E-mail: djulfa250@rambler.ru
Поступила в редакцию 02.03.2021
После доработки 23.03.2021
Принята к публикации 23.03.2021
Аннотация
Изучен нижний первый коренной зуб (m1) пещерных медведей методами традиционной и геометрической морфометрии из позднеплейстоценовых местонахождений Урала. На основе анализа его размеров и формы впервые установлено, что малый пещерный медведь (Ursus ex gr. savini-rossicus) входил в состав фаун из пещер Сказка, Виашер, Динамитная, Чудесница и Черные кости. Подтверждено его присутствие в фаунах из пещер Медвежья, Махневская ледяная, Аша 1, Игнатьевская, Барсучий Дол. Ареал малого пещерного медведя в позднем плейстоцене занимал Северный, Средний и Южный Урал. Ареалы малого пещерного медведя и большого пещерного медведя (Ursus kanivetz) перекрывались на Урале от начала (морская изотопная стадия 5е) до середины (середина морской изотопной стадии 3) позднего плейстоцена.
В настоящее время на основе морфологических и молекулярных данных выделяют 7 видов пещерных медведей. Это медведь Денингера (Ursus deningeri von Reichenau, 1904), денингероидный медведь (U. deningeroides Mottl, 1964), кавказский пещерный медведь (U. kudarensis Baryshnikov, 1985), группа малых пещерных медведей, включающая малого (U. savini Andrews, 1922) и русского (U. rossicus Borissiak, 1930) пещерных медведей, и группа больших пещерных медведей, включающая европейского (U. spelaeus Rosenmüller, 1794) и уральского (U. kanivetz Vereshchagin, 1973) пещерных медведей [1–4]. Наиболее плохо изученной на сегодняшний день является группа малых пещерных медведей (U. ex gr. savini-rossicus) из-за ограниченности молекулярных данных и отсутствия таксономической ревизии. Малый пещерный медведь (U. savini) впервые был описан в 1922 г. Чарльзом Эндрюсом из местонахождения Bacton Forest Bed в Англии [5]. Десять лет спустя А.А. Борисяк описал мелкую форму пещерного медведя с территории Северного Кавказа (Краснодарский край) и назвал ее U. spelaeus rossicus [6]. В 1973 г. Н.К. Верещагин описал костные остатки мелкого медведя из пещеры Кизеловская как U. uralensis [7]. Позднее медведь из Кизеловской пещеры стал рассматриваться в качестве подвида U. rossicus uralensis [8]. В 2007 г. Г.Ф. Барышников [9] выделяет три подвида малого пещерного медведя: U. savini savini (Великобритания и Германия), U. s. rossicus (степная зона от Украины до Забайкалья) и U. s. uralensis (Средний Урал). Позднее Н. Спассов на основе изучения серийного краниологического материала из Болгарии предложил разделить мелких пещерных медведей на 2 вида: U. savini (который включает в себя U. rossicus) и U. uralensis [10]. В новых работах Г.Ф. Барышников [2, 3] рассматривает U. savini и U. rossicus как два самостоятельных вида, последний из которых включает подвид U. r. uralensis. На сегодняшний день группа малых пещерных медведей нуждается в ревизии на основании морфологического изучения ископаемых образцов и проведении новых молекулярно-генетических исследований. Поэтому в данной публикации мы рассматриваем малого пещерного медведя как группу savini-rossicus (U. ex gr. savini-rossicus). По сравнению с большим пещерным медведем малый пещерный медведь является почти не изученным видом, в том числе мало данных о его географическом распространении. На данный момент в музее Института экологии растений и животных УрО РАН собрано большое количество костных остатков пещерных медведей из местонахождений Северного, Среднего и Южного Урала. Это дает возможность оценить особенности распространения малого пещерного медведя на Урале.
Исследовано 232 зуба из 17 местонахождений Урала. Материалы из 12 местонахождений либо не были изучены ранее, либо нуждаются в проверке их принадлежности к тому или иному таксону. На основе радиоуглеродных дат и биостратиграфических данных все местонахождения с остатками пещерных медведей датируются началом и серединой (морская изотопная стадия (МИС) 5е – МИС 3) позднего плейстоцена [11–18].
Применены методы классической и геометрической морфометрии. В морфометрическом анализе использовались признаки, опубликованные ранее: наибольшая длина зуба и ширина талонида [9]. Морфометрические данные анализировались с помощью двумерных графиков. Расчеты выполнены в программе Past 2.17 [19]. Из литературных источников известно, что размеры зубов большого пещерного медведя значительно превосходят размеры зубов малого пещерного медведя [6, 9, 10]. Для проверки этого заключения были использованы две группы данных о размерах m1 известной таксономической принадлежности. Первая группа включает выборки зубов большого пещерного медведя (U. kanivetz) из пещер Среднего (Тайн (слой 1), Геологов 3) и Южного (Победа, Игнатьевская (слои 2–9), Заповедная) Урала [13–15]. Вторая группа включает выборки зубов малого пещерного медведя (U. ex gr. savini-rossicus) из ряда местонахождений: пещеры Иманай [16], Краснодара [6], Сибири [9], Bacton Forest Bed [9], пещеры Кизеловская [9].
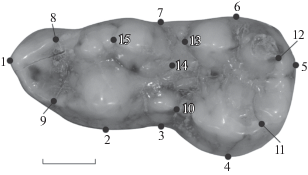
Исследование формы m1 проведено по 14 точкам, расположенным на окклюзиальной поверхности зуба. Точки 1–7 маркируют антериальный, постериальный, буккальный и лингвальный края коронки. Точки 8 и 9 маркируют границы параконида, точки 13 и 10 располагались в зоне контакта тригонида и талонида. Точки 11 и 12 маркируют края гипоконида и энтоконида. Точки 14 и 15 характеризуют границы главного бугра метаконидного комплекса (рис. 1). Для получения переменных формы проводили прокрустов анализ на координатах 14 точек. Для выявления структуры изменчивости данных провели анализ главных компонент на прокрустовых координатах. Расчеты выполнены в программе MorphoJ 1.07 [20].
Наши исследования подтвердили, что зубы большого и малого пещерных медведей хорошо различаются по размерам. Выборки зубов пещерных медведей с известной таксономической принадлежностью образовали две слабо перекрывающиеся между собой размерные группы (рис. 2).
Рис. 2.
Отношение наибольшей длины (L) к ширине талонида (W) m1 пещерных медведей, в мм. Крест – U. kanivetz из местонахождений Урала, ромб – U. ex gr. savini-rossicus из пещеры Иманай, квадрат – U. ex gr. savini-rossicus из местонахождения близ Краснодара [6], треугольник – U. ex gr. savini-rossicus из местонахождений Сибири [9]. Сплошная линия (прямоугольник) показывает границы изменчивости размеров m1 U. ex gr. savini-rossicus из местонахождения Bacton Forest Bed [9]. Пунктирная линия (прямоугольник) показывает границы изменчивости размеров m1 U. ex gr. savini-rossicus из пещеры Кизеловская [9]. Черный круг – пещерные местонахождения Урала: 1 – Кизеловская, 2 – Махневская Ледяная, 3 – Медвежья, 4 – Барсучий дол, 5 – Виашер, 6 – Чудесница, 7 – Тайн (слой 2), 8 – Сказка, 9 – Динамитная, 10 – Игнатьевская (10 слой), 11 – Черные кости, 12 – Куаламатская.
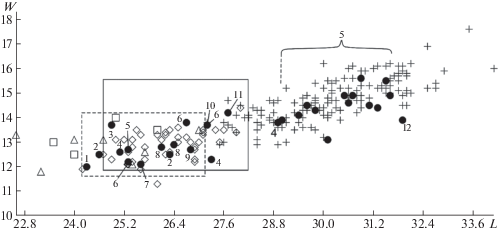
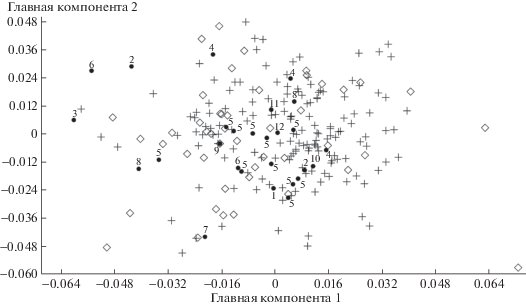
Первую группу (отличающуюся небольшими размерами) сформировали зубы U. ex gr. savini-rossicus, вторую группу (с более крупными размерами) сформировали зубы U. kanivetz. Зубы из 12 местонахождений попали в одну из образованных групп (рис. 2), что послужило основанием для определения их таксономической принадлежности. По форме m1 выборки разделились хуже (рис. 3). Можно заключить, что определение таксономической принадлежности пещерных медведей методом геометрической морфометрии по предложенной нами форме m1 значительно уступает возможности их дифференциации по размерам зуба и может служить только вспомогательным методом.
Рис. 3.
Результаты анализа формы m1 пещерных медведей методом главных компонент. Крест – U. kanivetz из местонахождений Урала, ромб – U. ex gr. savini-rossicus из пещеры Иманай. Черный круг – пещерные местонахождения Урала: 1 – Кизеловская, 2 – Махневская, 3 – Медвежья, 4 – Барсучий дол, 5 – Виашер, 6 – Чудесница, 7 – Тайн (слой 2), 8 – Сказка, 9 – Динамитная, 10 – Игнатьевская (10 слой), 11 – Черные кости, 12 – Куаламатская.
В результате исследований подтверждено присутствие U. kanivetz и U. ex gr. savini-rossicus в местонахождении пещера Медвежья [14] на Северном Урале. Для обоих таксонов это самая северная точка (62°05' с.ш., 58°05' в.д.) их ареалов на Урале. Впервые установлено наличие малого пещерного медведя в фаунах из пещер Виашер, Динамитная, Чудесница и, вероятно, Черные кости и подтверждено его наличие в фауне из пещеры Махневская ледяная [17] на Среднем Урале. На Южном Урале впервые установлено наличие малого пещерного медведя в фауне из пещеры Сказка и подтверждено его присутствие в фаунах из пещер Барсучий Дол [18], Аша 1 [13, 14] и Игнатиевская пещера (слой 10) [13]. Впервые установлено, что костные остатки пещерного медведя из пещеры Куаламатская принадлежат U. kanivetz.
В результате исследования размеров и формы m1 пещерных медведей впервые установлено, что малый пещерный медведь присутствует в фаунах из пещер Сказка, Виашер, Динамитная, Чудесница и Черные кости. Подтверждено, что U. kanivetz и U. ex gr. savini-rossicus обитали на Северном, Среднем и Южном Урале. В ряде местонахождений их остатки найдены в составе одних фаун, что указывает на перекрывание их ареалов на Урале. Время их совместного обитания составляет от начала (микулинское, казанцевское межледниковье; МИС 5е) до середины (середина брянского, каргинского интерстадиалов; середина МИС 3) позднего плейстоцена.
Список литературы
Rabeder G., Pacher M., Withalm G. Early Pleistocene bear remains from Deutsch-Altenburg (Lower Austria) // Mitt. Komm. Quartärkomm. Österr. Akad. Wiss. 2010. V. 17. P. 1–135.
Baryshnikov G.F., Puzachenko A.Y. Morphometry of upper check teeth of cave bears (Carnivora, Ursidae) // Boreas. 2019. V. 48. № 3. P. 581–604.
Baryshnikov G.F., Puzachenko A.Y. Morphometry of lower cheek teeth of cave bears (Carnivora, Ursidae) and general remarks on the dentition variability // Boreas. 2020. V. 49. № 3. P. 562–593.
Барышников Г.Ф. Кударский пещерный медведь: эволюция, тафономия, вымирание // Записки института истории материальной культуры. 2020. Вып. 22. С. 158–175.
Andrews C.W. XXII. Note on a bear (Ursus savini sp. n.) from the Cromer Forest-bed // Journal of Natural History. 1922. V. 9. № 50. P. 204–207.
Борисяк A.A. Новая раса пещерного медведя из четвертичных отложений Сев. Кавказа // Труды палеозоологического института. 1932. Т. 1. С. 137–201.
Верещагин Н.К. Краниологическая характеристика современных и ископаемых медведей // Зоологический журнал. 1973. Т. 52. № 6. С. 920–930.
Vereschagin N., Baryshnikov G. Small cave bear Ursus (Spelearctos) rossicus uralensis from the Kizel Cave in the Ural (Russia) // Geoloski zbornik. 2000. V. 15. P. 53–66.
Барышников Г.Ф. Семейство медвежьих (Carnivora, Ursidae). СПб.: Наука; 2007.
Spassov N., Hristova L., Ivanova S., Georgiev I. First record of the “small cave bear” in Bulgaria and the taxonomic status of bears of the Ursus savini Andrews – Ursus rossicus Borissiak group // Fossil Imprint. 2017. V. 73. P. 275–291.
Pacher M., Stuart A. J. Extinction chronology and palaeobiology of the cave bear (Ursus spelaeus) // Boreas. 2009. V. 38. P. 189–206.
Barlow A., Paijmans J.L.A., Federica A., et al. Middle Pleistocene genome calibrates a revised evolutionary history of extinct cave bears // Current Biology. 2021. V. 31. № 8. P. 1771–1779.
Смирнов Н.Г., Большаков В.Н., Косинцев П.А., и др. Историческая экология животных Южного Урала. Свердловск: УрО АН СССР; 1990.
Kosintsev P.A., Gasilin V.V., Gimranov D.O., Bachura O.P. Carnivores of the Ural in the late pleistocene and holocene // Quaternary International. 2016. V. 420. P. 145–155.
Baryshnikov G., Gimranov D., Kosintsev P. Variability of the upper incisors in the cave bears (Carnivora, Ursidae) from the Caucasus and Urals // Comptes Rendus Palevol. 2019. V. 18. № 2. P. 209–222.
Gimranov D.O., Kosintsev P.A. Quaternary large mammals from the Imanay Cave // Quaternary International. 2020. V. 546. P. 125–134.
Fadeeva T., Kosintsev P., Lapteva E., et al. Makhnevskaya Ledyanaya Cave (Middle Urals, Russia): Biostratigraphical Reconstruction // Quaternary International. 2020. V. 546. P. 135–151.
Kosintsev P.A., Yakovlev A.G., Plasteeva N.A., Gimranov D.O. Mammalian fauna from the Barsuchiy Dol Cave (the Southern Urals) // Russian Journal of The-riology. 2021. In Press.
Hammer O., Harper D.A.T., Ryan P.D. PAST: Paleontological statistics software package for education and data analysis // Palaeontologia Electronica. 2001. V. 4. № 1. P. 1–8.
Klingenberg C.P. MorphoJ: an integrated software package for geometric morphometrics // Molecular Ecology Resources. 2011. V. 11. P. 353–357.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни