

Доклады Российской академии наук. Науки о жизни, 2021, T. 501, № 1, стр. 586-592
ЦЕЛОМИЧЕСКАЯ СИСТЕМА ПРЕДСТАВИТЕЛЯ МОРСКИХ ЕЖЕЙ STRONGYLOCENTROTUS PALLIDUS (ECHINODERMATA, ECHINOIDEA) И ПРОБЛЕМА ЦЕЛОМИЧЕСКОЙ МЕТАМЕРИИ У ИГЛОКОЖИХ
О. В. Ежова 1, *, Ю. А. Кокуркина 1, К. И. Белолюбская 1, академик РАН В. В. Малахов 1
1 Московский государственный университет
имени М.В. Ломоносова
Москва, Россия
* E-mail: olga.ejova@gmail.com
Поступила в редакцию 01.04.2021
После доработки 04.06.2021
Принята к публикации 04.06.2021
Аннотация
Метамерия имеется у представителей основных филогенетических ветвей Bilateria. Среди Deuterostomia метамерия характерна для полухордовых и хордовых. Показано, что у морских ежей имеется 7 метамерных целомических колец, расположенных вдоль орально-аборальной оси тела. Подобная целомическая метамерия характерна также для представителей других классов иглокожих. Предложена гипотеза, согласно которой метамерные целомические кольца иглокожих представляют собой наследие метамерии, свойственной общему предку Deuterostomia. Диссимметрия в строении целомов иглокожих является результатом лежания предковых форм на правом боку, что привело к редукции целомов правой стороны. При переходе к сидячему образу жизни левые целомы сформировали метамерные целомические кольца.
В современной системе животного царства выделяется четыре основных группы билатерально-симметричных животных: Trochozoa, Lophophorata, Ecdysozoa и Deuterostomia [1, 2]. Целомическая метамерия имеется у представителей всех четырех стволов Bilateria, что позволяет предполагать, что и общий предок Bilateria был сегментированным организмом [2–4]. Среди вторичноротых несомненной метамерией обладают хордовые животные (Chordata). Метамерия Hemichordata выражается в наличии метамерных жаберных щелей и жаберных пор, происхождение которых, вероятно, связано с метамерией метанефридиев и энтодермальных кишечных карманов [5]. Все это позволяет поставить вопрос о том, сохраняется ли метамерия у иглокожих (Echinodermata). У иглокожих известны метамерные структуры, однако эта метамерия затрагивает придатки тела: имеется в виду метамерное расположение склеритов в стебельке морских лилий, метамерные позвонки в руках офиур и др. Проблема наличия у иглокожих целомической метамерии, сравнимой с метамерией хордовых, членистоногих, аннелид и других истинно метамерных животных, обсуждалось с позиции теории первичной гетерономной метамерии П.П. Иванова [6]. Согласно взглядам П.Г. Светлова и О.М. Ивановой-Казас [7, 8], три пары целомов диплеврулы рассматриваются как ларвальные сегменты, тогда как у хордовых оказываются развитыми не только ларвальные, но и постларвальные сегменты (туловищные сомиты) [9]. В настоящей работе мы попытаемся найти ответ на вопрос, сохраняются ли у иглокожих остатки соматоцельных целомических сегментов, сопоставимых с метамерными туловищными сомитами хордовых. Возможно, у иглокожих сохраняются остатки метамерии, которой обладал общий предок Deuterostomia, а может быть, и общий предок Bilateria. Мы предприняли новое исследование целомической организации типичного представителя иглокожих – морского ежа Strongylocentrotus pallidus (G.O. Sars, 1871) – с использованием современных методов 3D-реконструкции, чтобы получить объективное представление о взаимном расположении целомических компартментов.
Материалом для работы послужили 7 экземпляров S. pallidus (Echinoidea), собранные в районе биостанции “Дальние Зеленцы” (Баренцево море), с глубин 20–30 м. Диаметр собранных экземпляров составлял приблизительно 10 мм. Материал был зафиксирован в жидкости Буэна, а затем перенесен для консервации в 70-ный этиловый спирт. Для декальцинации применялся раствор ЭДТА (Трилон Б, Хелатон-3). Для гистологического исследования использовалась стандартная методика дегидратации материала в спиртах восходящей концентрации, пропитки парапластом и разложения на серии срезов толщиной 5 мкм с помощью микротома “Leica RM 2125”. Срезы окрашивались гематоксилином Караччи. Изучение срезов производилось с помощью микроскопа “Микмед-6” (LOMO, Санкт-Петербург, Россия, 2018) с цифровой камерой “MC-12”. По сериям гистологических срезов была осуществлена детальная 3D-реконструкция целомической системы в программе AMIRA, версия 6.5.0. Для реконструкции использовалась орально-аборальная серия, состоящая из 240 срезов.
Амбулакральное кольцо S. pallidus (производное левого гидроцеля) лежит в средней части тела и окружает глотку над Аристотелевым фонарем (рис. 1, 2, wvr). Оно оказывается как бы вставленным между двумя другими целомическими кольцами: перивисцеральным целомом (рис. 1, hgc), представляющим главную полость тела морского ежа, и обширным периоральным целомом (рис. 1, poc), окружающим челюстной аппарат. Между целотелиями всех трех целомических колец – амбулакрального, перивисцерального и периорального – проходит оральное кровеносное кольцо.
Рис. 1.
Организация целомической системы Strongylocentrotus pallidus. amp – мадрепоровая ампула; an – анальное отверстие; axc – осевой целом; egc – эпигастрический целом; gnc – генитальный целом; gut – кишечная трубка; hgc – гипогастрический целом; m – рот; mdp – мадрепоровая пластинка; pac – перианальный целом; pcd – перикардиальный целом; poc – периоральный целом; sc – каменистый канал; spr – соматоцельное перигемальное кольцо; wvr – амбулакральное кольцо.
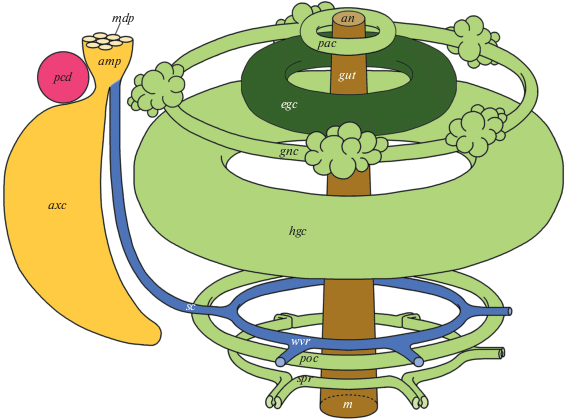
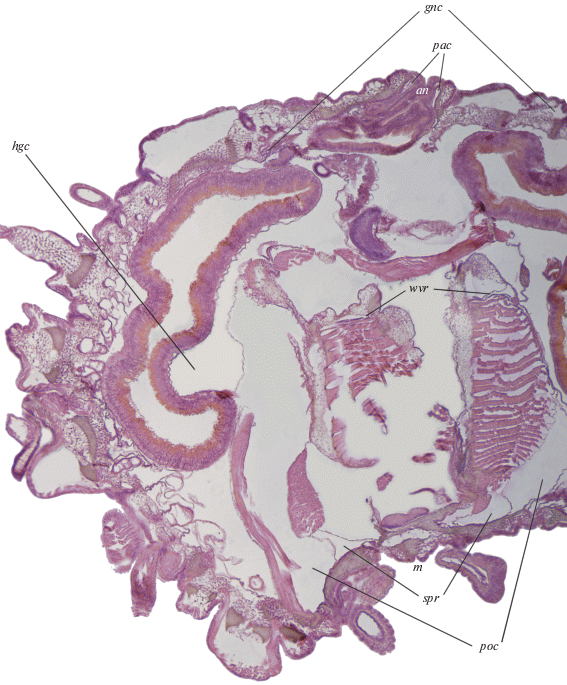
Рис. 2.
Производные левого соматоцеля Strongylocentrotus pallidus на сагиттальном срезе в области интеррадиуса CD. an – анальное отверстие; gnc – генитальный целом; hgc – перивисцеральный целом, образованный в результате слияния гипогастрического и эпигастрического целомов; m – рот; pac – перианальный целом; poc – периоральный целом; spr – соматоцельное перигемальное кольцо; wvr – амбулакральное кольцо.
Каменистый канал (рис. 1, sc) располагается в верхней, аборальной половине тела S. pallidus, и на своем оральном конце впадает в амбулакральное кольцо (рис. 1, wvr). Аборальный конец каменистого канала открывается в ампулу мадрепорита (рис. 1, amp), которая представляет собой очень уплощенную целомическую цистерну (производное левого аксоцеля). С внешней средой мадрепоровая ампула сообщается посредством множества (на изученных экземплярах до 200) тонких поровых каналов, которые пронизывают толщу мадрепоровой пластинки (рис. 1, mdp). Снизу, со стороны радиуса D, в мадрепоровую ампулу открываются каменистый канал (рис. 1, sc) и осевой целом (производное левого аксоцеля) (рис. 1, axc).
Осевой целом S. pallidus (рис. 1, axc) вытянут вдоль каменистого канала, к которому примыкает вплотную со стороны радиуса С. В аборальной области осевой целом представляет собой обширную, хорошо заметную на гистологических срезах полость. В других участках полость осевого целома практически вытеснена разросшимся осевым органом и поэтому представлена узкими щелевидными пространствами. На аборальной стороне осевой целом сообщается с ампулой мадрепорита (рис. 1, amp), а на оральной стороне оканчивается слепо в области впадения каменистого канала в амбулакральное кольцо. Аксоцельного перигемального целомического кольца осевой целом S. pallidus не образует.
Перикардиальный целом S. pallidus (производное правого аксоцеля) (рис. 1, pcd) представляет собой дисковидный целомический компартмент, не сообщающийся ни с какими иными целомическими полостями. С аборальной стороны перикард граничит с мадрепоровой ампулой (рис. 1, amp). На оральной стороне, со стороны радиуса С, к перикарду примыкают осевой целом (рис. 1, axc) и генитальный целом (рис. 1, gnc), а со стороны радиуса D – перивисцеральный целом (рис. 1, hgc).
Генитальный целом (производное левого соматоцеля) (рис. 1, 2, gnc) представляет собой целомическое кольцо, примыкающее к аборальной стенке тела S. pallidus и окружающее заднюю кишку. Генитальное кровеносное кольцо проходит в мезентерии между генитальным целомом и перивисцеральным целомом.
В самой аборальной части тела S. pallidus имеется маленькое целомическое кольцо, окружающее заднюю кишку. Это перианальное кольцо (рис. 1, 2, pac), происходящее от левого соматоцеля. Оно примыкает к обширному перивисцеральному целому.
Перивисцеральный целом (рис. 2, hgc) представляет собой очередное целомическое кольцо, окружающее кишку. Он образуется в результате слияния двух целомических полостей: одного из производных левого соматоцеля (гипогастрическое целомическое кольцо) (рис. 1, hgc) и единственного производного правого соматоцеля (эпигастрическое целомическое кольцо) (рис. 1, egc).
Периоральное (или перифарингеальное) целомическое кольцо S. pallidus (рис. 1, 2, poc) располагается в оральной половине тела животного и окружает глотку. Оно имеет очень сложную форму, поскольку вдается во внутренние желоба пирамид, в пространства между самими пирамидами, заключает в себя зубы, т.е. как бы “обволакивает” многочисленные известковые элементы Аристотелева фонаря. Периоральный целом окружен перивисцеральным целомом с боков и с аборальной стороны. В циркуморальном мезентерии между перивисцеральным и периоральным целомами проходит оральное кровеносное кольцо.
Соматоцельное перигемальное целомическое кольцо S. pallidus (производное левого соматоцеля) (рис. 1, 2, spr) лежит в самой оральной части тела. Здесь оно окружает челюстной аппарат и примыкает к периоральному целому. От соматоцельного перигемального кольца в каждый радиус отходит по паре целомических выростов.
Таким образом, целомы S. pallidus, за исключением осевого и перикардиального, образуют серию из 7 циркуминтестинальных колец, расположенных вдоль орально-аборальной оси тела. Подобное расположение целомических колец характерно и для других групп иглокожих. В частности, у разных видов морских звезд можно насчитать от 6 до 7 колец, у офиур – 7, у голотурий – 4, у морских лилий – 4 [10–12].
Наличие метамерных целомических колец у Echinodermata позволяет предполагать, что метамерия иглокожих происходит от метамерии, свойственной общим предкам вторичноротых животных (рис. 3). Метамерия Hemichordata складывается из следующих элементов: 1) предротовой сегмент, содержащий диссимметричные левый (хоботный) и правый (перикардиальный) целомы, 2) околоротовой сегмент, содержащий симметричные воротниковые целомы, и 3) туловищный целом, в котором диссепименты не сохраняются, но метамерия туловищного отдела очевидна в метамерии жаберных пор, жаберных щелей и дивертикулов кишечника. У хордовых животных метамерия выражается в наличии премандибулярных, мандибулярных и туловищных сомитов. Это позволяет предполагать, что общий предок Deuterostomia обладал метамерией, сходной с таковой у Annelida или зародышей Arthropoda, т.е. обладал предротовым сегментом, околоротовым сегментом и метамерным туловищем. В каждом из сегментов имелась пара целомических мешков. У Hemichordata и Cephalochordata имеет место диссимметрия в организации целомов предротового отдела. Как известно, иглокожие обладают значительно более выраженной диссимметрией целомов, чем все остальные вторичноротые. У иглокожих сохраняется только левый гидроцель (гомолог левого воротникового целома). Как следует из данных настоящей работы и данных других исследователей [10–12], диссимметричным оказывается и строение туловищных целомов. В частности у S. pallidus имеется 5 целомических колец, происходящих от левого соматоцеля личинки, и только одно происходящее от правого соматоцеля. Такое же соотношение характерно и для морских звезд, офиур, голотурий и морских лилий.
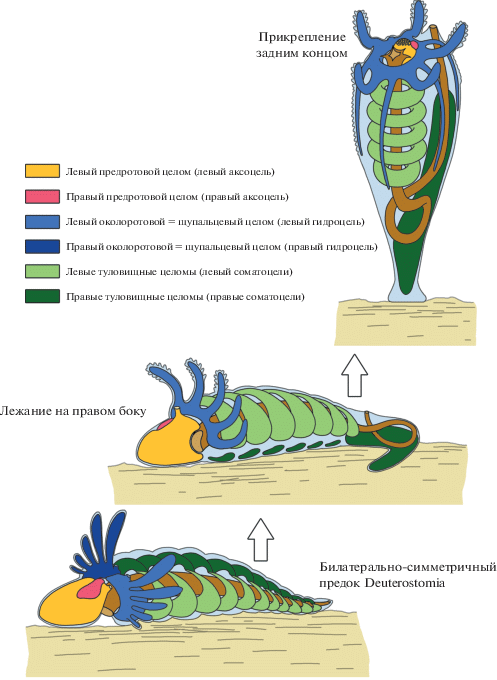
Для объяснения диссимметричной организации иглокожих предлагались гипотезы, согласно которым предполагаемые предки иглокожих были обращены к субстрату правой стороной тела, в результате чего и возникла диссимметрия целомов [13, 14]. При этом щупальцевый аппарат левой стороны тела и левый гидроцель сохранились, тогда как щупальцевый аппарат на правой стороне и соответствующий ему правый гидроцель подверглись полной редукции [13, 14]. Можно предполагать, что в этом случае редукции подверглись также и большинство метамерных соматоцельных целомов правой стороны (рис. 3).
С другой стороны, Холланд предполагает, что редукция правых переднего и среднего целомов объясняется лежанием на левом боку [15]. Впоследствии предки иглокожих перешли к прикрепленному образу жизни, в результате чего сформировалась вторичная радиальная симметрия, а сохранившиеся левые целомы воротникового и туловищного отделов сформировали циркуминтестинальные целомические кольца (рис. 3). Любопытно, что у Echinodermata даже расположение целомических колец соответствует тому порядку, в котором располагаются метамерные целомы у полухордовых и хордовых. Аксоцельное перигемальное кольцо (которое отсутствует у морских ежей) соответствует самому переднему хоботному целому полухордовых (премандибулярным сомитам хордовых), за ним следует амбулакральное кольцо, гомологичное левому воротниковому целому полухордовых (левому мандибулярному сомиту хордовых), далее следуют соматоцельный перигемальный целом, периоральный целом, гипогастрический целом и генитальный целом, которые соответствуют левым туловищным сомитам хордовых. Эпигастрический целом, происходящий от правого соматоцеля личинки, вероятно, соответствует стебельку предполагаемого предка, который использовался как заякоривающий орган. В рамках предлагаемой гипотезы предки иглокожих заякоривались в субстрате модифицированным задним концом тела, на основе которого сформировался стебелек сидячих представителей этого типа. На первый взгляд последнему предположению противоречит тот факт, что личинки современных морских лилий прикрепляются к субстрату предротовым участком тела. Этот факт обычно интерпретируется как свидетельство того, что стебелек морских лилий происходит от предротовой лопасти личинки. Однако если бы это было справедливо, то в стебельке морских лилий находились бы производные аксоцеля (предротового целома). Но в стебельке морских лилий находятся производные правого соматоцеля (камерный орган). По нашему мнению, в онтогенезе морских лилий нашли отражение два процесса: 1) ползание предковых форм на предротовой лопасти, подобно тому, как это сейчас имеет место у крыложаберных полухордовых (Graptolithoidea, бывшие Pterobranchia); 2) прикрепление задним концом тела, как это свойственно молодым кишечнодышащим полухордовым (Enteropneusta).
Разумеется, предложенная гипотеза нуждается в обосновании данными современной биологии развития и в подкреплении палеонтологическим материалом.
Список литературы
Halanych K.M. The new view of animal phylogeny // Annu. Rev. Ecol. Evol. Syst. 2004. V. 35. P. 229–256.
Малахов В.В. Новая система билатерий // Вестник РАН. 2010. Т. 80. № 1. С. 27–44.
Balavoine G., Adoutte A. The Segmented Urbilateria: A Testable Scenario // Integr. Comp. Biol. 2003. V. 43. P. 137–147.
Seaver E.C. Segmentation: mono- or polyphyletic? // Int. J. Dev. Biol. 2003. V. 47. P. 583–595.
Ezhova O.V., Malakhov V.V. The nephridial hypothesis of the gill slit origin // J. Exp. Zool. (Mol. Dev. Evol.). 2015. № 324B. P. 647–652.
Иванов П.П. Первичная и вторичная метамерия тела // Журнал общей биологии. 1944. Т. 5. Вып. 1. С. 61–94.
Светлов П.Г. О первичной гетерономии состава тела позвоночных (к 15-летию со дня смерти П.П. Иванова) // Архив анатомии, гистологии и эмбриологии. 1957. Т. 34. № 2. С. 3–22.
Иванова-Казас О.М. Сравнительная эмбриология беспозвоночных животных: Иглокожие и полухордовые. М.: Наука, 1978. 164 с.
MacBride E.W. Text-book of embryology. Vol. I: Invertebrates. London: Macmillan & Co, 1914. 692 p.
Gemmill J. F. The development of the starfish Solaster endeca Forbes // Trans. Zool. Soc. London. 1912. V. 20. № 1. P. 1–71.
Smiley S. Metamorphosis of Stichopus californicus (Echinodermata: Holothuroidea) and its phylogenetic implication // Biol. Bull. 1986. V. 171. P. 611–631.
Ezhova O.V., Malakhov V.V. Axial complex of Crinoidea: comparison with other Ambulacraria // J. Morph. 2020. V. 281. № 11. P. 1456–1475.
Малахов В.В. Проблема происхождения иглокожих в свете данных по их эмбриональному развитию // Проблемы изучения ископаемых и современных иглокожих. Таллин: АН Эстонской ССР. Ин-т геологии, 1989. С. 14–23.
Jefferies R.P.S., Brown N.A., Daley P.E.J. The early phylogeny of chordates and echinoderms and the origin of chordate left-right asymmetry and bilateral symmetry // Acta Zool. (Stockh). 1996. V. 77. P. 101–122.
Holland N.D. The meaning of developmental asymmetry for echinoderm evolution: A new interpretation // In Echinoderm phylogeny and evolutionary biology (Paul C.R.C., Smith A.B., eds.). Oxford: Clarendon Press, 1988. P. 13–25.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни