

Доклады Российской академии наук. Науки о жизни, 2021, T. 501, № 1, стр. 505-510
НОВЫЙ ВИД Typhlomys (Platacanthomyidae, Rodentia) ИЗ СРЕДНЕГО ПЛЕЙСТОЦЕНА СЕВЕРНОГО ВЬЕТНАМА
Академик РАН А. В. Лопатин *
Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 25.06.2021
После доработки 16.07.2021
Принята к публикации 16.07.2021
Аннотация
Из среднеплейстоценового пещерного местонахождения Там-Хай в северном Вьетнаме (провинция Лангшон) по фрагменту верхнечелюстной кости и изолированным зубам описан новый вид роющих сонь Typhlomys stegodontis sp. nov. Эта первая находка ископаемых Platacanthomyidae во Вьетнаме заполняет среднеплейстоценовый пробел в палеонтологической летописи семейства.
В настоящее время на севере Вьетнама обитает весьма редкая роющая соня Typhlomys chapensis Osgood, 1932, представитель небольшого семейства колючих сонь (Platacanthomyidae) [1]. Этот мелкий мышеобразный грызун, живущий в тропических горных лесах, парадоксальным образом приспособлен к древолазанию и к рытью; при этом он почти слеп и ориентируется при передвижении с помощью ультразвуковой эхолокации, подобно рукокрылым [2]. Современный ареал вида ограничивается высокогорным районом Шапа в провинции Лаокай на севере Вьетнама [1, 3, 4] и прилегающей областью Южного Китая (= T. cinereus jingdongensis Wu et Wang, 1984; Юньнань, Цзиндун) [5–8]. Палеонтологическая история Typhlomys во Вьетнаме не изучена, тогда как в Китае остатки нескольких видов роющих сонь обнаружены в верхнем миоцене [9, 10], плио-плейстоцене [11], нижнем плейстоцене [12–16] и низах верхнего плейстоцена [17].
Ниже описаны ископаемые остатки Typhlomys из Вьетнама – фрагмент верхнечелюстной кости и изолированные зубы T. stegodontis sp. nov. из среднего плейстоцена пещеры Там-Хай, провинция Лангшон, уезд Биньзя (Bình Gia), собранные экспедиционным отрядом Палеонтологического института им. А.А. Борисяка РАН (ПИН) в марте 2021 г. в ходе работ Совместного Российско-Вьетнамского Тропического научно-исследовательского и технологического центра (Тропический центр, Ханой). Пещера Там-Хай (Thẩm Hai, Tham Hai) входит в пещерный комплекс Там-Кхюиен (Thẩm Khuyên), известный совместными находками Homo erectus (Dubois, 1893) и Gigantopithecus blacki von Koenigswald, 1935 и датированный средним плейстоценом (Там-Хай: 300–200 тыс. лет, 250 тыс. лет; Там-Кхюиен: 475 ± 125 тыс. лет) [см. 18–20]. В сборах 2021 г. из пещеры Там-Хай обильно представлены остатки мелких млекопитающих, включая насекомоядных (Erinaceidae, Soricidae), рукокрылых (в том числе Pteropodidae) и грызунов. Среди последних преобладают разнообразные Muridae, сравнительно многочисленны Hystricidae и Sciuridae, а наиболее редки Cricetidae (Arvicolinae) и Platacanthomyidae (Typhlomys stegodontis sp. nov.).
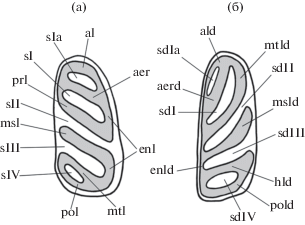
Описанный в статье материал хранится в коллекции ПИН, Москва. Терминология строения зубов по [9], с изменениями (см. рис. 1). Фотографии сделаны с помощью сканирующего электронного микроскопа Tescan Vega 3 XMU в ПИН.
Рис. 1.
Терминология элементов строения моляров Typhlomys (по [9], с изменениями): а – правый M1; б – правый M1. Обозначения: гребни: aer – передний дополнительный гребень (на верхних молярах); aerd – передний дополнительный гребень (на нижних молярах); al – антеролоф; ald – антеролофид; enl – эндолоф; enld – эндолофид; hld – задний дополнительный гребень (гиполофид); msl – мезолоф; msld – мезолофид; mtl – металоф; mtld – металофид; pol – постеролоф; pold – постеролофид; prl – протолоф; складки и долинки: sI – парафоссетта (синклин I); sIa – антерофоссетта (синклин Ia); sII – мезофоссетта (синклин II); sIII – метафоссетта (синклин III); sIV – постерофоссетта (синклин IV); sdI – мезофоссеттид (синклинид I); sdIa – антерофоссеттид (синклинид Ia); sdII – эктофоссеттид (синклинид II); sdIII – гипофоссеттид (синклинид III); sdIV – постерофоссеттид (синклинид IV).
Отряд Rodentia Bowdich, 1821
Семейство Platacanthomyidae Alston, 1876
Род Typhlomys Milne-Edwards, 1877
Typhlomys stegodontis Lopatin, sp. nov.
Название вида по совместному нахождению с вымершим родом хоботных Stegodon.
Голотип – ПИН, № 5826/501, фрагмент правой верхнечелюстной кости с M1–M2 (рис. 2); северный Вьетнам, провинция Лангшон, пещера Там-Хай; средний плейстоцен.
Рис. 2.
Typhlomys stegodontis sp. nov., голотип ПИН, № 5826/501, фрагмент правой верхнечелюстной кости с M1–M2 с окклюзиальной стороны; Вьетнам, провинция Лангшон, пещера Там-Хай; средний плейстоцен.
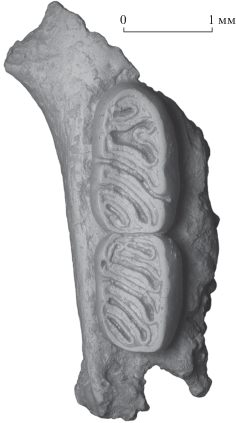
Описание (рис. 2, 3). Задний край основания переднего корня скуловой дуги расположен на уровне передней части M1. Моляры лофодонтные, с плоской или немного вогнутой жевательной поверхностью, состоящей из шести поперечных и диагональных гребней и пяти складок и долинок (на M3 – из пяти гребней и четырех долинок). Верхние моляры трехкорневые, нижние – двухкорневые.
Рис. 3.
Typhlomys stegodontis sp. nov.: а – голотип ПИН, № 5826/501, правые M1–M2; б – экз. ПИН, № 5826/551, левый M1; в – экз. ПИН, № 5826/552, правый M2; г – экз. ПИН, № 5826/502, правый M1; д – экз. ПИН, № 5826/503, левый M1; е – экз. ПИН, № 5826/553, левый M1; ж – экз. ПИН, № 5826/554, правый M3; Вьетнам, провинция Лангшон, пещера Там-Хай; средний плейстоцен.

Верхние моляры со слабо выраженной лингвальной гипсодонтией. На M1–M2 диагональные средние гребни (mid-ridges; передний дополнительный гребень, протолоф, мезолоф и металоф) и соответствующие долинки (парафоссетта, мезофоссетта и метафоссетта) ориентированы под углом около 45–60° к продольной оси зуба (рис. 2).
M1 длинный и узкий, заметно сужающийся спереди (рис. 3а). Антеролоф короткий, сильно изогнут вперед. Передний дополнительный гребень (anterior extra ridge) полный, ограничивает сзади короткую и узкую замкнутую антерофоссетту (синклин Ia). Парафоссетта (синклин I) закрыта лабиально и лингвально, на голотипе раздвоенная – имеет антеролингвальное ответвление, достигающее эмалевой стенки зуба. Протолоф и мезолоф широкие, субпараллельные, в лингвальной части сближаются, но не сливаются. Мезофоссетта (синклин II) открыта лабиально и лингвально, оба устья узкие и неглубокие (рис. 3а). При сильном стирании мезофоссетта лингвально замкнута, образован непрерывный эндолоф (рис. 3б). Метафоссетта (синклин III) открыта лабиально, ее лингвальный конец касается эмалевой стенки зуба в постеролингвальном углу жевательной поверхности. Металоф и постеролоф короткие. Постерофоссетта (синклин IV) маленькая, закрытая.
М2 субпрямоугольных очертаний, сужающийся сзади, по строению в целом близок M1. Антеролоф длинный, прямой. Имеется относительно длинная антерофоссетта. На голотипе парафоссетта имеет небольшое переднее ответвление, примыкающее к антерофоссетте с лабиальной стороны. Все долинки закрыты лингвально, эндолоф полный. При слабом стирании мезофоссетта лабиально открытая, другие долинки замкнуты с лабиальной стороны, хотя соответствующие гребни не сращены. Таким образом, на ранней стадии стирания эктолоф не сформирован (рис. 3а), тогда как на более поздней – хорошо развит (рис. 3в).
Судя по частично сохранившимся на голотипе альвеолам, M3 по длине существенно уступает M2 (рис. 2).
На M1 диагональные средние гребни (передний дополнительный гребень, металофид и мезолофид) и соответствующие долинки (мезофоссеттид и эктофоссеттид) ориентированы под углом около 35°–55° к продольной оси зуба, задний дополнительный гребень (гиполофид) и гипофоссеттид – субпоперечные (около 70°–85°). M1 удлиненный, узкий в передней части. Антеролофид плавно изогнут вперед. Передний дополнительный гребень отчетливый, диагонально ориентированный (сильно скошен постеролингвально), соединяется с антеролофидом и эндолофидом и замыкает длинный антерофоссеттид (синклинид Ia), иногда разделенный надвое (рис. 3г). Металофид, соединяющий протоконид с метаконидом, является самым длинным из диагональных гребней. Вместе с передним дополнительным гребнем он обрамляет мезофоссеттид (синклинид I). При слабом стирании (рис. 3д) дентиновые поля антеролофида и металофида в передней части жевательной поверхности разделены, а их эмалевые стенки соприкасаются, но не сливаются. В сочетании с небольшим углублением стенки зуба в области соединения указанных гребней это свидетельствует о первоначальном наличии узкого и мелкого антеролабиального выхода мезофоссеттида. С лингвальной стороны мезофоссетид открыт на ранней стадии стирания (рис. 3д), но замкнут на более поздних (рис. 3г, 3е). Эктофоссеттид (синклинид II) открыт лабиально и лингвально либо упирается в эндолофид (рис. 3е). Мезолофид хорошо развит, имеет три варианта строения (морфотипа): (1) полный мезолофид, связывающий мезоконид с эндолофидом (рис. 3г); (2) свободный мезолофид, лингвально ограниченный слитыми долинками эктофоссеттида и гипофоссеттида (рис. 3д; см. сходное строение мезолофида M1 у позднемиоценового T. primitivus [9, табл. II , фиг. 11, 12 , рис. 44a ] и современного T. cinereus [13, рис. 1–7]); (3) укороченный мезолофид, в лингвальной части присоединенный к заднему дополнительному гребню (рис. 3е). Гипофоссеттид (синклинид III) открыт лабиально, но замкнут лингвально (или имеет общий выход с эктофоссеттидом, см. рис. 3д). Задний дополнительный гребень (posterior extra ridge) соединяет гипоконид с энтоконидом (рис. 3г, 3д) или мезолофидом (рис. 3е). Постеролофид длинный, субпоперечный, параллельный заднему дополнительному гребню. Постерофоссеттид (синклинид IV) широкий, замкнутый (однако при слабом стирании постеролофид и гиполофид лингвально не слиты, см. рис. 3д), в антеролингвальной части иногда связан с эктофоссеттидом (рис. 3е).
M3 субтреугольный, короткий, сужающийся сзади (рис. 3ж). Антерофоссеттид очень маленький, антеролингвальный по положению. Передний дополнительный гребень, металофид, мезолофид и постеролофид широкие, ориентированы диагонально. Мезофоссеттид протяженный, замкнутый. Эктофоссеттид открыт лабиально, а в своей лингвальной части достигает эндолофида. Гипофоссеттид закрытый, касается лингвальной и лабиальной эмалевых стенок зуба. Задний дополнительный гребень и постерофоссеттид отсутствуют.
Размеры в мм. Длина M1–M2: 3.25 (голотип).
Размеры зубов (L – длина, W – ширина, EH – лабиальная высота коронки, IH – лингвальная высота коронки): M1: L – 1.8, W – 1.1, EH – 0.55, IH – 0.75 (голотип); L – 1.7, W – 0.95, EH – 0.5, IH – 0.7 (экз. ПИН, № 5826/551); M2: L – 1.4, W – 1.1, EH – 0.5, IH – 0.7 (голотип); L – 1.4, W – 0.9, EH – 0.45, IH – 0.6 (экз. ПИН, № 5826/552); M1: L – 1.95, W – 1.15, EH – 0.75, IH – 0.75 (экз. ПИН, № 5826/502); L – 1.9, B – 1.1, EH – 0.9, IH – 0.8 (экз. ПИН, № 5826/553); L – 1.8, W – 1.05, EH – 0.8, IH – 0.8 (экз. ПИН, № 5826/503); M3: L – 1.0, W – 0.8, EH – 0.3, IH – 0.4 (экз. ПИН, № 5826/554).
Сравнение. По размерам зубов новый вид существенно мельче позднемиоценового T. storchi Qiu et Ni, 2019 и раннеплейстоценового T. macrourus Zheng, 1993, меньше позднемиоценового T. hipparionum Qiu, 1989, но крупнее T. primitivus Qiu, 1989 (поздний миоцен), T. intermedius Zheng, 1993, T. anhuiensis Jin et al., 2009 (ранний плейстоцен), современных T. nanus Cheng et al., 2017, T. cinereus Milne-Edwards, 1877, T. chapensis Osgood, 1932, T. daloushanensis Wang et Li, 1996 и T. huangshanensis Hu et Zhang in Hu et al., 2021 (T. chapensis из Вьетнама и Южного Китая, остальные виды из Китая). От всех перечисленных видов T. stegodontis sp. nov. отличается наличием морфотипа M1 с раздвоенной парафоссеттой и морфотипа M1 с лингвальным соединением мезолофида с задним дополнительным гребнем. Кроме того, новый вид отличается от T. primitivus относительно более широким в передней части M1; от T. hipparionum – относительно менее удлиненными M1 и M3; от T. storchi – менее вытянутым в длину M1 с открытыми эктофоссеттидом и гипофоссеттидом; от T. macrourus – более низкокоронковыми и менее удлиненными молярами, относительно более узким в передней части M1 и наличием антерофоссетты на M2; от T. intermedius – лабиально открытыми мезофоссеттой и метафоссеттой M1; от T. anhuiensis – сильнее редуцированным M3; от T. chapensis – наличием антерофоссетты на M2 и лабиально закрытым мезофоссеттидом M1; от T. cinereus – относительно более узким в передней части M1 и наличием антерофоссетты на M2; от T. daloushanensis – относительно более узкими M1–M2 и сильнее вытянутым M2; от T. nanus – наличием антерофоссетты на M2 и постерофоссеттида на M1, а также закрытым мезофоссеттидом M1; от T. huangshanensis – более узкими M1–M2 и наличием антерофоссетты на M2.
Замечания. Большинство ископаемых роющих сонь известно преимущественно по изолированным зубам, поэтому сравнение зубных размеров T. stegodontis sp. nov. с вымершими видами Typhlomys проведено по верхним и нижним молярам соответствующих локусов (данные по [9–16]). В случае современных видов Typhlomys сравниваемым параметром является длина ряда верхних моляров [см. 6–8]. Сохранность голотипа T. stegodontis sp. nov. позволяет провести его сравнение с современными видами по длине M1–M3. Расстояние от переднего конца коронки M1 до заднего края сохранившегося фрагмента верхнечелюстной кости, обломанной приблизительно на уровне середины альвеолы заднего лабиального корня M3 (см. рис. 2), составляет 4.0 мм. С учетом предполагаемых размеров отсутствующего M3 аппроксимированная длина ряда верхних моляров голотипа T. stegodontis sp. nov. достигает приблизительно 4.2–4.3 мм, что превосходит средние значения длины M1–M3 всех современных видов и находится вблизи верхней границы пределов изменчивости этого параметра у самых крупных из них (минимум – среднее – максимум: T. nanus: 3.13–3.31–3.49 мм, 3 экз. [7]; T. cinereus: 3.2–3.3–3.4 мм, 2 экз. [7]; 3.4–3.46–3.62 мм, 4 экз., самки; 3.39–3.74–3.86 мм, 4 экз., самцы [1]; 3.4–3.6–3.7 мм, 6 экз. [6]; T. chapensis: 3.46–3.61–3.85 мм, 21 экз. [7]; 3.55–3.85–4.09 мм, 11 экз., самки; 3.57–3.96–4.14 мм, 4 экз., самцы [1]; 3.9–4.0–4.2 мм, 9 экз. [6]; T. huangshanensis: 3.56–3.74–4.05 мм, 9 экз. [8]; T. daloushanensis: 3.57–3.81–4.05 мм, 29 экз. [7]; 3.7–3.9–4.2 мм, 49 экз. [6]).
Материал. Помимо голотипа, из местонахождения Там-Хай шесть изолированных зубов: M1 (экз. ПИН, № 5826/551), M2 (экз. ПИН, № 5826/552), три M1 (экз. ПИН, №№ 5826/502, 503, 553) и M3 (экз. ПИН, № 5826/554). Голотип и экз. ПИН, №№ 5826/502, 503 происходят из основного костеносного слоя красно-бурых алевролитов верхней галереи пещеры (21°55′51.7″ с.ш., 106°25′3.6″ в.д.); экз. ПИН, №№ 5826/551–554 – из обособленной северной галереи (21°55′52.5″ с.ш., 106°25′4.1″ в.д.).
Стегодонтовая роющая соня Typhlomys stegodontis sp. nov. из пещеры Там-Хай – первая находка ископаемых Platacanthomyidae во Вьетнаме. Биохронологически она заполняет среднеплейстоценовый пробел в известной палеонтологической летописи семейства.
Анализ палеонтологических данных позволяет сделать некоторые наблюдения относительно эволюционной истории Typhlomys. Уже в позднем миоцене этот род был представлен тремя филогенетическими ветвями, к одной из которых принадлежит мелкий T. primitivus, ко второй – более крупный T. hipparionum, а к третьей – уклоняющийся по строению M1 и относительно очень крупный T. storchi [9, 10]. В раннем плейстоцене первая линия включала T. intermedius и T. anhuiensis [12–16], и именно с этой группой видов связывается среднеплейстоценовый T. stegodontis sp. nov., а также рецентный T. cinereus и все прочие современные виды, недавно выделенные из его состава при поддержке молекулярно-генетических данных [см. 1, 7, 8]. Вторая линия оборвалась в раннем плейстоцене на стадии T. macrourus, обладавшего относительно высокими коронками моляров [13]. Третья ветвь, видимо, не имела потомков. Следует отметить, что симпатрические ископаемые виды (T. primitivus, T. hipparionum и T. storchi в позднем миоцене, T. intermedius и T. macrourus в раннем плейстоцене) существенно отличались друг от друга по величине.
С учетом новых данных о современном разнообразии Typhlomys [1, 7, 8] видовая принадлежность сравнительно крупного представителя этого рода из раннеплейстоценового пещерного местонахождения Лунгудун (Longgudong) в провинции Хубэй Китая, отнесенного к ныне живущему виду T. cinereus [14, рис. 5.27, A –F], подлежит пересмотру. Указанная раннеплейстоценовая форма отличается от T. cinereus более крупными размерами зубов [14, табл. 5.35 ], относительно более узким в передней части M1 и наличием антерофоссетты на M2. Этими признаками, а также общим строением M1 и M3, данная форма сближается с T. stegodontis sp. nov.; при этом она отличается от нового вида строением мезофоссетты M2, закрытой лабиально, но открытой лингвально (у стегодонтовой роющей сони мезофоссетта M2, напротив, открыта лабиально и закрыта лингвально). Вывод о видовой принадлежности раннеплейстоценовой хубэйской формы может быть сделан по итогам специального изучения морфотипической изменчивости массовых зубных остатков из пещеры Лунгудун [см. 14]; пока ее следует рассматривать как Typhlomys sp.
В верхнем плейстоцене идентифицирован только Typhlomys cinereus из Южного Китая [17]. При этом в настоящее время считается, что современное довольно высокое видовое разнообразие роющих сонь (не менее пяти аллопатрических видов) на территории юго-восточного Китая и северного Вьетнама является результатом таксономической диверсификации в условиях изолированных горных областей [1, 6]. Открытие T. stegodontis sp. nov. в плейстоцене северного Вьетнама и современное видовое разнообразие рода позволяют предполагать существование нескольких средне- и позднеплейстоценовых видовых линий Typhlomys. Современный ареал T. chapensis (горы Хоангльеншон на юге Китая и северо-западе Вьетнама) и среднеплейстоценовая область обитания T. stegodontis sp. nov. (северо-восточный Вьетнам) разделены долиной Красной реки (Хонгха), которая в плейстоцене могла быть одной из главных преград для распространения мелких млекопитающих в данном регионе.
Список литературы
Abramov A.V., Balakirev A.E., Rozhnov V.V. An enigmatic pygmy dormouse: molecular and morphological evidence for the species taxonomic status of Typhlomys chapensis (Rodentia: Platacanthomyidae) // Zoological Studies. 2014. V. 53. Art. № 34. P. 1–9.
Panyutina A.A., Kuznetsov A.N., Volodin I.A., et al. A blind climber: the first evidence of ultrasonic echolocation in arboreal mammals // Integrative Zoology. 2017. V. 12. № 2. P. 172–184.
Osgood W.H. Mammals of the Kelley-Roosevelts and Delacour Asiatic expeditions // Field Museum of Natural History. Zoological Series. 1932. V. 18. № 10. P. 193–339.
Кузнецов Г.В. Млекопитающие Вьетнама. М.: Т-во научных изданий КМК, 2006.
Wu D., Wang G. A new subspecies of Typhlomys cinereus Milne-Edwards from Yunnan, China // Acta Theriologica Sinica. 1984. V. 4. № 3. P. 213–215.
Wang Y., Li C., Chen Z. Taxonomy, distribution and differentiation on Typhlomys cinereus (Platacanthomyidae, Mammalia) // Acta Theriologica Sinica. 1996. V. 16. № 1. C. 54–66.
Cheng F., He K., Chen Z.-Z., et al. Phylogeny and systematic revision of the genus Typhlomys (Rodentia, Platacanthomyidae), with description of a new species // Journal of Mammalogy. 2017. V. 98. № 3. P. 731–743.
Hu T.-L., Cheng F., Xu Z., et al. Molecular and morphological evidence for a new species of the genus Typhlomys (Rodentia: Platacanthomyidae) // Zoological Research. 2021. V. 42. № 1. P. 100–107.
Qiu Z. Fossil platacanthomyids from the hominoid locality of Lufeng, Yunnan // Vertebrata PalAsiatica. 1989. V. 27. № 4. P. 268–283.
Qiu Z., Ni X. Platacanthomyids (Rodentia, Mammalia) from the Late Miocene Yuanmou hominoid locality of Yunnan, China // Fossil Imprint. 2019. V. 75. № 3–4. P. 383–396.
Qiu Z.D., Jin C.Z. Platacanthomyid remains from the late Cenozoic deposits of East China // Vertebrata PalAsiatica. 2017. V. 55. № 4. P. 315–330.
Zheng S. Quaternary rodents of Sichuan-Guizhou area, China. Beijing: Science Press; 1993.
Fejfar O., Kalthoff D.C. Aberrant cricetids (platacanthomyines, Rodentia, Mammalia) from the Miocene of Eurasia // Berliner geowissenschaftliche Abhandlungen. Ser. E. 1999. V. 30. P. 191–206.
Zheng S. Rodentia. In: Zheng S., ed. Jianshi hominid site. Beijing: Science Press; 2004. pp. 121–181, 387–394.
Jin C.Z., Zhang Y.Q., Wei G.B., et al. Rodentia Bowdich, 1821. In: Jin C., Liu J., eds. Paleolithic site – the Renzidong Cave, Fanchang, Anhui Province. Beijing: Science Press; 2009. P. 166–220, 401–412.
Wang Y., Jin C., Mead J.I. New remains of Sinomas-todon yangziensis (Proboscidea, Gomphotheriidae) from Sanhe karst Cave, with discussion on the evolution of Pleistocene Sinomastodon in South China // Quaternary International. 2014. V. 339–340. P. 90–96.
Wei G., Huang W., Boëda E., et al. Recent discovery of a unique Paleolithic industry from the Yumidong Cave site in the Three Gorges region of Yangtze River, southwest China // Quaternary International. 2017. V. 434. Pt A. P. 107–120.
Olsen J.W., Ciochon R.L. A review of evidence for postulated Middle Pleistocene occupations in Viet Nam // Journal of Human Evolution. 1990. V. 19. P. 761–788.
Ciochon R., Vu The Long, Larick R., et al. Dated co-occurrence of Homo erectus and Gigantopithecus from Tham Khuyen Cave, Vietnam // Proceedings of the National Academy of Sciences. 1996. V. 93. № 7. P. 3016–3020.
Zeitoun V., Chinnawut W., Debruyne R., et al. A sustainable review of the Middle Pleistocene benchmark sites including the Ailuropoda–Stegodon faunal complex: the Proboscidean point of view // Quaternary Internatio-nal. 2016. V. 416. P. 12–26.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни