

Доклады Российской академии наук. Науки о жизни, 2021, T. 501, № 1, стр. 511-516
РАННЕПЛЕЙСТОЦЕНОВАЯ РЫСЬ Lynx issiodorensis (Felidae, Carnivora) ИЗ ПЕЩЕРЫ ТАВРИДА, КРЫМ
А. В. Лавров 1, Д. О. Гимранов 2, *, Б. А. Вахрушев 3, академик РАН А. В. Лопатин 1
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
2 Институт экологии растений и животных Уральского отделения Российской академии наук
Екатеринбург, Россия
3 Крымский федеральный университет
им. В.И. Вернадского
Симферополь, Россия
* E-mail: djulfa250@rambler.ru
Поступила в редакцию 18.05.2021
После доработки 16.07.2021
Принята к публикации 16.07.2021
Аннотация
Из раннеплейстоценового местонахождения пещера Таврида (Крым, поздний виллафранк, возраст 1.8–1.5 млн. лет) описаны краниальные и нижнечелюстные остатки двух взрослых особей Lynx issiodorensis (Croizet et Jobert, 1828). Этот вид рысей был типичным представителем виллафранкской фауны Восточного Средиземноморья. Отмечена высокая краниологическая изменчивость L. issiodorensis.
Пещера Таврида была открыта в 2018 г. Она расположена на северном макросклоне Крымских гор, в междуречье рек Бештерек и Фундуклы (притоки р. Зуи, бассейн р. Салгир). В южном коридоре пещеры было обнаружено большое число ископаемых костей наземных позвоночных. Этот комплекс соответствует поздневиллафранкским фаунам Европы (зоны MNQ18–MNQ19) и псекупскому фаунистическому комплексу России. Время существования ассоциации определено в интервале 1.8–1.5 млн. л.н. [1, 2]. В поздневиллафранкское время пещера через горизонтальный южный коридор открывалась в склоне долины р. Зуя и использовалась хищниками как логово [3–5].
В ходе раскопок 2020 г. в южном коридоре пещеры в 150 м от входа были найдены остатки двух особей исуарской рыси Lynx issiodorensis (Croizet et Jobert, 1828): мозговая часть черепа с фрагментом левой глазницы сравнительно молодой особи (экз. ПИН, № 5644/90; рис. 1) и почти полная нижняя челюсть (без резцов и клыков) старой особи (экз. ПИН, № 5644/89; рис. 2). Кроме того, там же и в 100 м от выхода были найдены фрагменты трех лопаток, две плечевые кости, две локтевые кости, фрагменты пяти ребер, семи позвонков, две полные большие берцовые кости и другие фрагменты не менее чем еще от пяти взрослых особей.
Рис. 1.
Lynx issiodorensis (Croizet et Jobert, 1828), экз. ПИН, № 5644/90, мозговая часть черепа с фрагментом левой глазницы: a – с левой латеральной стороны; б – с дорсальной стороны; в – с вентральной стороны; Крым, пещера Таврида; нижний плейстоцен. Обозначения: 1 – сагиттальный гребень; 2 – левый теменной гребень; 3 – сосцевидный отросток; 4 – положение заглазничного перехвата; 5 – околозатылочный отросток; 6 – передний край глазницы; 7 – подъязычное отверстие; 8 – заднее рваное отверстие.
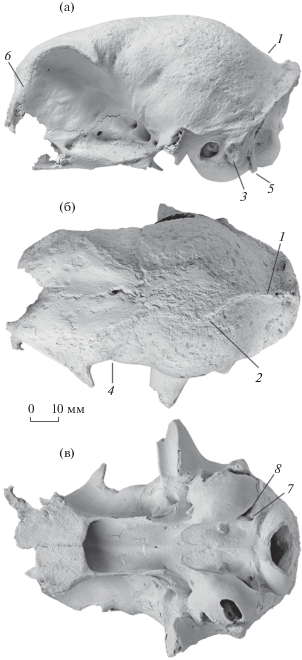
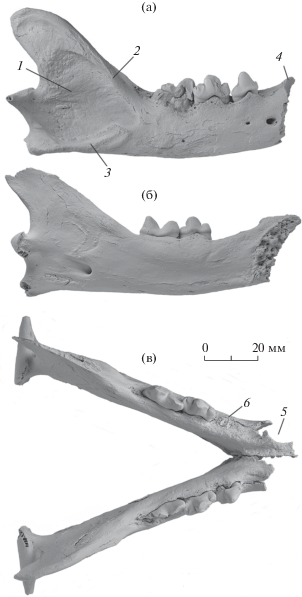
Рис. 2.
Lynx issiodorensis (Croizet et Jobert, 1828), экз. ПИН, № 5644/89, парные ветви нижней челюсти: а – экз. ПИН, № 5644/89a, правая нижнечелюстная кость с р3–m1 с лабиальной стороны; б – экз. ПИН, № 5644/89б, левая нижнечелюстная кость с р4–m1 с лингвальной стороны; в – левые и правая ветви нижней челюсти в анатомическом сочленении с окклюзиальной стороны; Крым, пещера Таврида; нижний плейстоцен. Обозначения: 1 – массетерная ямка; 2 – массетерный гребень; 3 – площадка для крепления средней порции массетера (m. masseter medialis); 4 – положение постеролингвального края альвеолы правого с1; 5 – край альвеолы левого с1; 6 – задний корень разрушенного при жизни левого р3 (альвеола переднего корня оссифицирована).
Описанные в статье материалы хранятся в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН), Москва. Принятые сокращения: Hbc – высота мозговой коробки; Lpo – посторбитальная длина черепа; Wbc – максимальная ширина мозговой коробки; Wpo – ширина заглазничного перехвата; L – длина; W – ширина; Ld – длина диастемы; La–p3 – длина нижней челюсти от заднего края сочленовного отростка до переднего края р3; dex – правый; sin – левый; n – число экземпляров; ЗММУ – Зоологический музей Московского государственного университета им. М.В. Ломоносова; APL – коллекция из местонахождения Апполония-1, Университет имени Аристотеля в Салониках, Греция; ING – коллекция из местонахождения Ингарано (Апулия), Зоологический музей, Рим, Италия; MHNB Prr – коллекция из местонахождения Перье, Музей естественной истории, Базель, Швейцария; MHNL QSV – коллекция из местонахождения Сен-Валье, Музей естественной истории, Лион, Франция; SBAU – Управление архивами археологии Умбрии, Италия; TB – коллекция из местонахождения Туркобуния, Афинский университет, Греция.
Сохранившаяся часть черепа (экз. ПИН, № 5644/90) включает мозговую коробку и большую часть левой глазницы (рис. 1). Поверхность кости умеренно эродирована. Череп принадлежал сравнительно мелкой особи. По форме мозговая коробка сходна с экз. MHNB Prr 200 L. issiodorensis из местонахождения Этуэр, ранний виллафранк Франции [6]. У экз. ПИН, № 5644/90 высота нейрокраниума по дорсовентральной оси больше, чем у экз. MHNL QSV 1133 из Сен-Валье, поздний плиоцен Франции [7]. Экз. SBAU 337653 L. issiodorensis valdarnesis Werdelin, 1981 из Панталлы (поздний виллафранк Италии, Перуджа) резко отличается от экз. ПИН, № 5644/90 уплощенной формой мозговой коробки [8].
Сагиттальный гребень развит на черепах L. issiodorensis из Сен-Валье (экз. MHNL QSV 1133), Панталлы (экз. SBAU 337653) и Перье (экз. MHNB Prr 200), также как у многих экземпляров современной обыкновенной рыси L. lynx (Linnaeus, 1758). Этот гребень у Lynx очень низкий – на большей части протяжения его высота 1–2 мм. Сагиттальный гребень на черепе из пещеры Таврида развит только в задней части свода черепа – его длина всего 15 мм. Теменные гребни находятся в 13–17 см друг от друга (рис. 1б). Форма лирообразной фигуры, образованной этими гребнями, соответствует типу С, выделенному для L. pardinus (Temmink, 1827) [9]. У современной L. lynx степень развития сагиттального гребня очень изменчива (n = 104, ЗММУ): у 36.7% экземпляров он хорошо выражен, у 38.3% случаев он не развит, и положение теменных гребней близко к таковому у экз. ПИН, № 5644/90 (рис. 1). В 25% случаев состояние теменных гребней близко к слиянию: расстояние между ними составляет 3–4 мм, но сагиттальный гребень не сформирован. Передний край мозговой коробки широкий, что типично для Lynx. Отношение высоты мозговой коробки к заглазничной длине черепа (Hbc/Lpo) у образца из Крыма близко к значениям у L. lynx (табл. 1).
Таблица 1.
Размеры нейрокраниума современной Lynx lynx (Linnaeus, 1758) и L. issiodorensis (Croizet et Jobert, 1828) из нижнего плейстоцена Перуджи (Италия) и Крыма
| Параметр | L. lynx, ЗММУ, n = 24, min–max | L. issiodorensis | ||
|---|---|---|---|---|
| Перуджа, экз. SBAU 337653 [8] | Крым, экз. ПИН, № 5644/90 | |||
| 1 | Максимальная ширина мозговой коробки (Wbc) | 54.8–64.5 | 62.3 | 58.8 |
| 2 | Длина от носо-лобного шва до края затылочного гребня | 94.9–118.2 | 105.31 | 88.7 |
| 3 | Ширина заглазничного перехвата (Wpo) | 35.2–44.1 | 43.6 | 44.8 |
| 4 | Ширина в сосцевидных отростках | 58.1–71.8 | 58.3 | 58.9 |
| 5 | Высота мозговой коробки (от уровня основной затылочной кости до верхней точки теменной кости, Hbc) | 46.3–55.6 | 50.71 | 53.9 |
| 6 | Высота мозговой коробки от верхней точки теменной кости до вентрального края слухового барабана | 57.1–69.2 | 56.91 | 49.8 |
| 7 | Длина от заглазничного перехвата до заднего края затылочного гребня (заглазничная длина черепа, Lpo) | 65.9–78.6 | 71.4 | 78.1 |
| 8 | Отношение высоты мозговой коробки к заглазничной длине черепа (Hbc/Lpo) | 0.644–0.749, n = 22; среднее = 0.678 | 0.797 | 0.637 |
| 9 | Отношение ширины заглазничного перехвата к ши-рине мозговой коробки (Wpo/Wbc) | 0.631–0.728 n = 22; среднее = 0.693 |
0.699 | 0.762 |
Форма и размер слухового барабана у экз. ПИН, № 5644/90 как у современной L. lynx: он удлиненный и выпуклый (рис. 1в), в отличие от округлого слухового барабана у экз. SBAU 337653 из Перуджи. Заглазничный перехват выражен слабо – отношение ширины заглазничного сужения к ширине мозговой коробки (индекс Wpo/Wbc) у Lynx больше, чем у Puma и многих других родов Felinae. Значение этого индекса у современных и ископаемых видов Lynx имеет невысокую изменчивость (табл. 1). На черепе экз. ПИН, № 5644/90 отверстие подъязычного нерва (for. hypoglossum) и заднее рваное отверстие (for. lacerum posterius) расположены близко в небольшом углублении: образуется подобие единого синуса, что свойственно L. pardinus, L. rufus (Schreber, 1777) и L. issiodorensis (рис. 1в) [9, 10]. Такое состояние данного признака считается ключевой характеристикой этих трех видов [10].
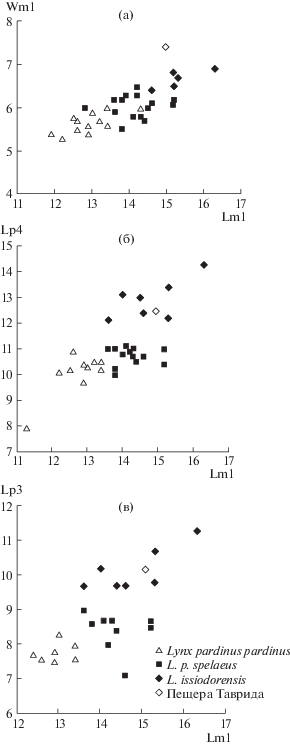
Левая и правая нижнечелюстные кости, экз. ПИН, № 5644/89, принадлежали одной старой особи. Ветви нижней челюсти идеально стыкуются между собой в сохранившейся части симфиза, имеют одинаковые размеры и особенности зубной морфологии (рис. 2). Угол смыкания ветвей нижней челюсти в симфизе равен 50° (как у современной L. lynx). По соотношению размеров Lp3/Lm1 экз. ПИН, № 5644/89 занимает среднее положение в поле значений L. issiodorensis (рис. 3). L. pardinus характеризуется меньшими размерами р3, р4 и m1 (рис. 3) [11].
Рис. 3.
Диаграммы соотношения размеров нижних щечных зубов (в мм) у некоторых представителей рода Lynx (по [11] c учетом экз. ПИН, № 5644/89): а – отношение длины к ширине m1 (Lm1/Wm1); б – отношение длины р4 к длине m1 (Lp4/Lm1); в – отношение длины р3 к длине m1 (Lp3/Lm1).
Левый р3 сломан при жизни (альвеола его переднего корня оссифицирована), правый m1 расколот вдоль длинной оси и его лабиальная часть отсутствует. Оставшиеся зубы имеют следы значительного износа, особенно на вершинах бугорков, в их лабиальной части (рис. 2). Особенность морфологии нижней челюсти L. issiodorensis – компактность щечного зубного ряда р3–m1 (при более крупных размерах челюсти Lр3–m1 у нее меньше, чем у L. lynx). Превосходство в длине челюсти у L. issiodorensis обеспечивается за счет большего размера области массетерной ямки. Эта особенность выражается отношением Lp3–m1/La–p3 (табл. 2). Данные различия в пропорциях отражают только индивидуальную изменчивость. Форма венечного и углового отростков, длина диастемы, положение заднего края симфиза и другие детали морфологии нижней челюсти у L. issiodorensis и L. lynx очень сходны.
Таблица 2.
Размеры нижней челюсти и зубов (p3, p4, m1) Lynx issiodorensis (Croizet et Jobert, 1828) и L. lynx (Linnaeus, 1758) (L – длина, W – ширина; Ld – длина диастемы)
| Вид, экземпляр, местонахождение | р3 | р4 | m1 | Ld | Lp3–m1 | |||
|---|---|---|---|---|---|---|---|---|
| L | W | L | W | L | W | |||
| L. issiodorensis, экз. ПИН, № 5644/89; Таврида, Крым, Россия | 10.2 | 5.8 | 13.3 | 6.8 | 15.1 | 7.3 | 8.0 | 38.4 |
| L. issiodorensis, экз. FP1-2001-0120; Фонелас P-1, Гранада, Испания [14] | 9.3 | 4.6 | 11.4 | 5.3 | 14.1 | 5.8 | 9.8 | 34.7 |
| L. issiodorensis, экз. APL-14; Aполлония 1, Греция [13] | 9.8 | 5.3 | 11.6 | 6.0 | 13.8 | 6.8 | 8.5 | 35.0 |
| L. issiodorensis, экз. CA-101; пещеры Альманзора, Испания [11] | 9.7 | 4.2 | 12.1 | 5.3 | 13.6 | 5.9 | 8.3 | 34.4 |
| L. issiodorensis, экз. SBAU 167337; Панталла, Перуджа, Италия [8] | 9.4 | – | 11.0 | – | 14.5 | – | 10.8 | 35.2 |
| L. issiodorensis [17, табл. X (I), фиг. 1–3], Туркобуния (TB-5), Греция [12, 17] | 10.2 | 6.1 | 15.2 | 7.2 | 16.2 | 8.6 | 11.2 | 40.4 |
| L. lynx, ЗММУ, n = 24 | 8.5–11.1 | 4.6–6.1 | 11.3–12.4 | 5.3–6.4 | 14.1–16.2 | 5.5–7.3 | 5.3–11.1 | 34.5–40.2 |
В строении зубов экз. ПИН, № 5644/89 имеются следующие особенности. Главный бугорок р3 невысокий (длина зуба больше его высоты), в отличие от экземпляра из Аполлонии-1, Греция [12]. Для L. issiodorensis характерно развитие переднего базального бугорка р3, который хорошо выражен и на экз. ПИН, № 5644/89а (рис. 2а). Последний премоляр р4 имеет очень крупный базальный бугорок, высота р4 меньше длины (строение сходно с р4 экз. ТВ-5 из нижнего виллафранка Греции [13]). На экз. ПИН, № 5644/89 Lm1 больше среднего значения для вида. При этом по массивности зуба (Wm1) он занимает одно из первых мест в выборке (рис. 3). По длине лезвие параконида равно лезвию протоконида. Высота параконида равна высоте протоконида. Задний гребень протоконида хорошо развит. Талонид рудиментарный, метаконид отсутствует, что типично для L. issiodorensis (рис. 2). Прикорневая часть альвеолярного края челюсти в области обоих m1 имеет следы патологической деградации (рис. 2а, 2в), вероятно, связанной с периодонтитом.
L. issiodorensis близка по краниальной морфологии к L. lynx и L. canadensis Kerr, 1792. Наиболее надежным признаком, разделяющим их, является полуобъединенное состояние заднего рваного отверстия и подъязычного отверстия (их расположение в едином углублении) у L. issiodorensis. Такое полуобъединенное состояние этих отверстий характерно также для современных и ископаемых L. pardina. Считается, что L. issiodorensis, L. pardina и L. rufus отличаются от L. lynx и L. canadensis еще одним краниальным признаком – отсутствием сагиттального гребня [9, 10]. Однако развитие сагиттального гребня у L. lynx очень изменчиво. В то же время черепа L. issiodorensis из Сен-Валье, Перье и Панталлы имеют сагиттальный гребень. У черепов из Италии и Испании, отнесенных к L. pardina, сагиттальный гребень тоже иногда сформирован (экз. ING 76) [10]. Таким образом, степень развития этого признака у Lynx варьирует.
По морфологии посткраниального скелета виды Lynx неотличимы.
Сравнение находок из пещеры Таврида с материалами по L. issiodorensis [6–8, 10, 11, 14] позволяет заключить, что крымские экземпляры принадлежат этому виду. Они имеют минимальное сходство с наиболее уклоняющейся формой вида L. issiodorensis valdarnesis. В этой статье мы не рассматриваем подвиды исуарской рыси. Обзор разнообразия форм, объединяемых в L. issiodorensis, и уровень изменчивости краниологических признаков у ископаемых и современных представителей Lynx приводят к заключению о недостаточной разработанности таксономии ископаемых рысей рубежа плиоцена–плейстоцена в Евразии.
Lynx issiodorensis иногда считается прямым предком всех современных видов рода Lynx [15]; при этом несоответствие ряда признаков этой гипотезе объясняется реверсивной эволюцией. Анализ ДНК современных Felidae не подтверждает эту гипотезу [16].
Экземпляры L. issiodorensis из пещеры Таврида являются типичными представителями данного вида – как по размерам, так и по пропорциям черепа, зубов и деталям их морфологии. При этом отмеченные выше особенности строения черепа L. issiodorensis из европейских местонахождений свидетельствуют о высокой внутривидовой краниологической изменчивости.
Находка L. issiodorensis в Крыму подтверждает ранее определенный поздневиллафранкский возраст костеносного слоя пещеры Таврида [1, 2] и является связующим звеном между восточной и западной частями ареала этого вида в раннем плейстоцене.
Список литературы
Лопатин А.В., Вислобокова И.А., Лавров А.В., и др. Пещера Таврида – новое местонахождение раннеплейстоценовых позвоночных в Крыму // Доклады Академии наук. 2019. Т. 485. № 3. С. 381–385.
Вислобокова И.А., Титов В.В., Лавров А.В., и др. Раннеплейстоценовые винторогие антилопы (Artiodactyla, Bovidae) из пещеры Таврида (Крым, Россия) // Палеонтологический журнал. 2020. № 1. С. 78–88.
Гимранов Д.О., Лавров А. В., Старцев Д.Б., и др. Первая находка этрусского медведя (Ursus etruscus, Ursidae, Carnivora) в Крыму (пещера Таврида, ранний плейстоцен) // Доклады РАН. Науки о жизни. 2020. Т. 491. № 1. С. 130–133.
Оксиненко П.В., Лавров А.В. История формирования пещеры Таврида – памятника фауны позвоночных позднего виллафранка // Вестник МГУ. Сер. геогр. 2021. № 1. С. 27–42.
Лавров А.В., Гимранов Д.О., Старцев Д.Б., Зайцев Б.А. Саблезубые кошки (Carnivora, Felidae, Machairodontinae) из нижнего плейстоцена пещеры Таврида, Крым // Доклады РАН. Науки о жизни. 2020. Т. 495. С. 567–571.
Kurten B. The lynx from Etouaires, Lynx issiodorensis (Croizet and Jobert), Late Pliocene // Annales Zoologici Fennici. 1978. V. 15. P. 314–322.
Viret M.J. Le loess a bancs durcis de Saint-Vallier (Drôme) et sa faune de mammifères villafranchiens // Nouvelles Archives du Muséum d’Histoire Naturelle de Lyon. 1954. V. 4. P. 1–195.
Cherin M., Iurino D.A., Sardella R. New well-preserved material of Lynx issiodorensis valdarnensis (Felidae, Mammalia) from the Early Pleistocene of Pantalla (central Italy) // Bollettino della Societá Paleontologica Italiana. 2013. V. 52. № 2. P. 103–111.
García-Perea R., Gisbert J., Palacios F. Review of the biometrical and morphological features of the skull of the Iberian lynx, Lynx pardinus (Temminck, 1824) // Säugetierkundl. Mitt. 1985. V. 32. P. 249–259.
Boscaini A., Madurell-Malapeira J., Llenas M., Martínez-Navarro B. The origin of the critically endangered Iberian lynx: speciation, diet and adaptive changes // Quaternary Science Reviews. 2015. V. 123. P. 247–253.
Montoya P., Morales J., Sendra J. Una mandíbula de Lynx issiodorensis (Croizet y Jobert, 1828) (Carnivora, Mammalia) en el Plioceno inferior de Cuevas de Almanzora (Almería, España) // Revista Española de Paleontología. 2001. Núm. ext. P. 125–131.
Koufos G.D. The Villafranchian carnivoran guild of Greece: implications for the fauna, biochronology and paleoecology // Integrative Zoology. 2014. V. 9. P. 444–460.
Koufos G.D. The Pleistocene carnivores of the Mygdonia basin (Macedonia, Greece) // Annales de Paléontologie. 1992. V. 78. P. 205–257.
Garrido G. Linces y guepardos (Felidae, Carnivora, Mammalia) en el Plioceno superior terminal del Fonelas P-1 (Cuenca de Guadix, Granada) // Cuadernos del Museo Geominero. 2008. № 10. P. 231–249.
Werdelin L. The evolution of lynxes // Annales Zoologici Fennici. 1981. V. 18. P. 37–71.
Li G., Davis B.W., Eizirik E., Murphy W.J. Phylogenomic evidence for ancient hybridization in the genomes of living cats (Felidae) // Genome Research. 2016. V. 26. № 1. P. 1–11.
Symeonidis N., De Vos J. Grossäuger-Funde aus den Pleistozänen spaltenfülungen von Turkovunia in Athen // Annales géologiques des Pays helléniques. 1977. V. 28. P. 135–144.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни