

Доклады Российской академии наук. Науки о жизни, 2022, T. 502, № 1, стр. 70-76
ЗЕМЛЕРОЙКИ РОДА CHODSIGOA (SORICIDAE, LIPOTYPHLA) ИЗ ПЛЕЙСТОЦЕНА ВЬЕТНАМА
Академик РАН А. В. Лопатин *
Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 20.10.2021
После доработки 02.11.2021
Принята к публикации 03.11.2021
- EDN: PVOJGF
- DOI: 10.31857/S2686738922010152
Аннотация
Из плейстоцена северного Вьетнама описаны первые за пределами Китая ископаемые находки представителей рода Chodsigoa: C. caovansunga Lunde, Musser et Son, 2003 (фрагмент верхнечелюстной кости с A3–M3, изолированный M1 и нижнечелюстная кость с I1 и P4–M2) из среднего плейстоцена пещеры Там-Хай (провинция Лангшон) и C. hoffmanni Chen et al., 2017 (фрагмент верхнечелюстной кости с P4–M2) из верхнего плейстоцена пещеры Лангчанг (провинция Тханьхоа). Описанные экземпляры являются первыми ископаемыми находками указанных видов. Они свидетельствуют, что во Вьетнаме представители рода Chodsigoa в плейстоцене распространялись на юг значительно дальше, чем в настоящее время.
Азиатские землеройки рода Chodsigoa Kastschenko, 1907 относятся к числу наименее изученных представителей семейства Soricidae. Согласно современным молекулярно-генетическим и морфологическим данным [1], этот род включает девять рецентных видов, обитающих на территории восточной Азии от Центрального Китая на севере до северных районов Мьянмы, Таиланда и Вьетнама на юге. В северном Вьетнаме представлены три редких вида: землеройка Лове C. parca Allen, 1923 (эндемичный подвид C. p. lovei Osgood, 1932) в провинции Лаокай, землеройка Као Ван Шунга C. caovansunga Lunde, Musser et Son, 2003 и землеройка Хоффманна C. hoffmanni Chen et al., 2017 в провинции Хазянг [1–3]. Вид C. caovansunga известен также из Южного Китая (провинция Юньнань) [4], а C. hoffmanni – из Южного Китая (Юньнань, Гуйчжоу) [1, 5] и Центрального Китая (провинция Хубэй) [6]. Подвид C. parca parca Allen, 1923 отмечен в Юньнани, на севере Мьянмы и Таиланда [1].
Ископаемая летопись рода Chodsigoa довольно скудна и при этом ограничена плио-плейстоценом Китая и осложнена таксономическими проблемами, связанными с взаимоотношениями с близкими родами Soriculus Blyth, 1854 и Episoriculus Ellerman et Morrison-Scott, 1951. Для Chodsigoa характерны наличие трех (а не четырех, как у Soriculus и Episoriculus) одновершинных верхних антемоляров, сильно редуцированная пигментация зубов, передний наклон венечного отростка, крупный нижнечелюстной мыщелок и специфическое переднее положение его нижней суставной фасетки [7, 8]. Кроме того, P4–M2Chodsigoa характеризуются очень глубокой выемкой (эмаргинацией) заднего края коронки [7], а P4 демонстрирует более высокое положение в зубном ряду по сравнению с Episoriculus [8]. Таким образом, представителей Chodsigoa можно уверенно идентифицировать в ископаемом состоянии не только по черепным остаткам, но и по находкам верхнечелюстных фрагментов, изолированных верхних щечных зубов и нижнечелюстных фрагментов.
С учетом указанных признаков раннеплиоценовый вид Soriculus praecursus Flynn et Wu, 1994 из провинции Шаньси [8] не должен рассматриваться в составе Chodsigoa. К ископаемым представителям Chodsigoa следует относить вымерший вид C. bohlini (Young, 1934) из среднеплейстоценового местонахождения Чжоукоудянь в Пекине [7, 9], форму неясного систематического положения из нижнего плейстоцена пещеры Женьцзидун (Renzidong) в провинции Аньхой, указанную [10, рис. 4.10 ] как “Chodsigoa cf. C. praecursu”, а также находки, отнесенные к рецентным видам – C. parca из нижнего плейстоцена провинции Сычуань [11], C. hypsibia (de Winton et Styan, 1899) (= Sori-culus hypsibius) из нижнего плейстоцена провинции Хубэй [12, рис. 5.8 ] и C. smithii Thomas, 1911 из верхнего плейстоцена Чунцина (Chongqing) [13]. “Chodsigoa youngi sp. nov.” из среднего плейстоцена провинции Аньхой [14] является nomen nudum. Определения “Chodsigoa n. sp.” из нижнего плиоцена провинции Ганьсу [15, рис. 3], C. sa-lenskii (Kastschenko, 1907) из среднего плейстоцена провинции Хубэй [16], C. hybsibia (“C. lamula”) и “C. salenskii?” из среднего – верхнего плейстоцена провинции Гуйчжоу [17] не сопровождаются описаниями и иллюстрациями материалов.
В настоящей статье из плейстоцена северного Вьетнама описаны первые ископаемые находки представителей Chodsigoa за пределами Китая: C. caovansunga из среднего плейстоцена пещеры Там-Хай (провинция Лангшон, уезд Биньзя) и C. hoffmanni из низов верхнего плейстоцена пещеры Лангчанг (провинция Тханьхоа, уезд Батхыок). Материалы собраны экспедиционными отрядами Палеонтологического института им. А.А. Борисяка РАН (ПИН) в марте – апреле 2021 г. в ходе работ Совместного Российско-Вьетнамского Тропического научно-исследовательского и технологического центра (Тропический центр, Ханой).
Пещера Там-Хай (Thẩm Hai, Tham Hai) входит в пещерный комплекс Там-Кхюиен (Thẩm Khuyên), известный совместными находками Homo erectus (Dubois, 1893) и Gigantopithecus blacki von Koenigswald, 1935 и датированный средним плейстоценом (Там-Хай: 300–200 тыс. лет или 250 тыс. лет; Там-Кхюиен: 475 ± 125 тыс. лет) [см. 18].
Костеносные слои пещеры Лангчанг (Lang Trang) содержат многочисленные остатки млекопитающих фауны Stegodon–Ailuropoda, которые датируются началом позднего плейстоцена [см. 19].
Описанный материал хранится в коллекции ПИН, Москва. Терминология и методика измерения зубов и нижнечелюстных костей по [20], с изменениями. Размеры в мм. Обозначения измерений зубов: AW – передняя ширина; BH – лабиальная высота; BL – лабиальная длина; L – наибольшая длина; LL – лингвальная длина; PE – длина до выемки заднего края (posterior emargination); PW – задняя ширина; TAW – ширина талонида; TRW – ширина тригонида; W – наибольшая ширина. PE-индекс (индекс эмаргинации) рассчитан по формуле (LL+BL)/2PE – 1 [20]. Фотографии сделаны с помощью сканирующего электронного микроскопа Tescan Vega II XMU в ПИН.
Отряд Lipotyphla Haeckel, 1866
Семейство Soricidae Fischer von Waldheim, 1817
Подсемейство Soricinae Fischer von Waldheim, 1817
Триба Nectogalini Anderson, 1879
Род Chodsigoa Kastschenko, 1907
Chodsigoa caovansunga Lunde, Musser et Son, 2003
Chodsigoa caovansunga: [2, с. 37, рис. 5b, 5d, 6b, 7 ].
Голотип – Американский музей естественной истории, AMNH 274154, череп и тушка взрослой самки; Вьетнам, провинция Хазянг, уезд Висюен, община Каобо, местонахождение Тэйконлинь II (Tay Con Linh II); современный.
Описание (рис. 1, 2). Размеры мелкие для рода. Переднее отверстие подглазничного канала находится над M1, передний край заднего отверстия – на уровне метакона M2. Небольшое слезное отверстие располагается на уровне метастиля M1. Рудимент корня скуловой дуги находится на уровне задней части M2.
Рис. 1.
Chodsigoa caovansunga Lunde, Musser et Son, 2003, экз. ПИН, № 5826/560, фрагмент левой верхнечелюстной кости с A3–M3: а – с лабиальной стороны, б – с окклюзиальной стороны, в – с окклюзиальной стороны с лингвальным наклоном; Вьетнам, провинция Лангшон, пещера Там-Хай; средний плейстоцен.
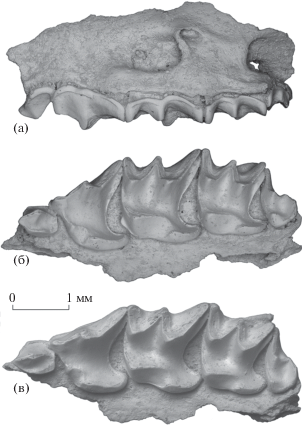
Рис. 2.
Chodsigoa caovansunga Lunde, Musser et Son, 2003, экз. ПИН, № 5826/562, левая нижнечелюстная кость с I1, P4–M2 и альвеолами A1 и M3: а – с лабиальной стороны, б – с окклюзиальной стороны, в – с лингвальной стороны, г – с задней стороны; Вьетнам, провинция Лангшон, пещера Там-Хай; средний плейстоцен.

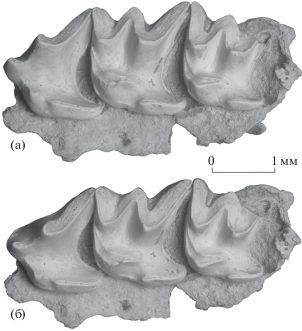
Рис. 3.
Chodsigoa hoffmanni Chen et al., 2017, экз. ПИН, № 5792/525, фрагмент левой верхнечелюстной кости с P4–M2 и альвеолами M3: а – с окклюзиальной стороны, б – с окклюзиальной стороны с лингвальным наклоном; Вьетнам, провинция Тханьхоа, пещера Лангчанг; низы верхнего плейстоцена.
A3 мелкий, низкий, с округло-треугольной окклюзиальной поверхностью, резкими передним и задним продольными гребнями и мощными лабиальным и лингвальным цингулюмами. Вершина главного бугорка слабо пигментированная с лабиальной стороны.
P4–M1 с глубокой плавной выемкой заднего края (задней вырезкой, или эмаргинацией).
P4 относительно высокий и длинный, с прямым лабиальным краем. Вершина паракона имеет маленький слабо пигментированный участок на антеролабиальной стороне. Парастиль умеренно развитый, сильно выступающий вперед, связан с передним основанием паракона широким гребнем. Метастилярный выступ длинный, ориентирован постеролабиально. Гребень паракристы широкий и высокий, с отчетливой вырезкой. Протокон мелкий (меньше парастиля), в виде бугорка, связан с лингвальным основанием парастиля четким гребнем. Имеется короткий и тонкий гребень постпротокристы. Выступ гипокона сильно вытянут постеролингвально, с резким цингулоподобным гребнем по лингвальному краю. Лабиальный цингулюм очень тонкий, постцингулюм отчетливый.
M1–M2 относительно длинные и узкие, с мощным W-образным эктолофом. Паракон меньше и ниже метакона. Гребень препротокристы тонкий, резкий, постпротокриста широкая. Выступ гипокона сильно выдается назад (на M2 слабее, чем на M1), заметно сужается дистально. Лабиальный цингулюм тонкий, постцингулюм отчетливый, заметно расширен в основании задней стороны метастилярного выступа. Лингвальный цингулюм, расположенный на уровне задней части протокона, слабый. На M1 сильно выступает метастиль, на M2 – парастиль. На M2 задняя выемка развита слабее, чем на M1, лингвальный цингулюм едва заметный. На M1 в основании передней стороны протокона имеется короткий, но широкий прецингулюм.
M3 сильно редуцирован, расширен поперечно. Парастилярная доля сильно выступает лабиально. Гребни паракона соединены L-образно. Метакон рудиментарный, гребневидный. Препротокриста хорошо развита, постпротокристы нет. Основание протокона соединено с метаконом широким цингулоподобным гребнем, формирующим задний край коронки.
Горизонтальная ветвь нижнечелюстной кости сравнительно высокая и массивная, особенно под P4–M1. Подбородочное отверстие расположено под серединой M1. Симфиз достигает уровня передней части M1. Нижнечелюстное отверстие мелкое, узкое, вытянуто вертикально. Венечный отросток массивный, относительно низкий, его вершина узкая, сильно отогнута мезиально. Наружная височная ямка хорошо очерчена, простирается от области вершины венечного отростка до уровня немного вентральнее верхней сигмовидной вырезки. Спикула венечного отростка высокая, протягивается более чем на половину глубины верхней сигмовидной вырезки. Внутренняя височная ямка округленно-треугольной формы, большая, простирается до середины высоты венечного отростка. Область ниже внутренней височной впадины углубленная, отделена от впадины сильно выступающим толстым гребнем. На уровне нижней части верхней сигмовидной вырезки на дорсомедиальной поверхности основания сочленовного отростка хорошо заметна углубленная верхняя ямка крыловидного мускула. Сочленовный мыщелок сравнительно крупный. Дистальная сторона мыщелка относительно высокая, межсуставная область небольшая. Верхняя и нижняя фасетки мыщелка образуют L-образную фигуру с глубокой выемкой на медиальной стороне. Нижняя фасетка широкая, вынесена далеко вперед по отношению к верхней фасетке.
Вершины I1 и P4 ярко пигментированные.
Нижний резец I1 относительно короткий, слегка изогнутый, с крупным задним бугорком и двумя слабо выраженными дополнительными зубцами на слегка волнистой режущей кромке. Вершина резца загнута дорсально, лабиальный и лингвальный цингулиды широкие. Медиальный гребень узкий, тянется от передней трети резца к лингвальному цингулиду. Задний край лабиальной стороны основания коронки резца находится на уровне вершины P4, задний край лингвальной стороны – перед альвеолой A1.
P4 с заостренным передним концом, высокой конической вершиной, двумя ветвями постерокристида, субпоперечным гребнем, соединяющим ветви постерокристида дистально, коротким талонидом и мощно развитыми цингулидами. Нижний край основания коронки P4 как с лабиальной, так и с лингвальной стороны расположен немного (приблизительно на высоту цингулидов) вентральнее оснований нижних моляров.
На M1–M2 талонид немного шире тригонида. Гребень энтоконида тонкий, но полный, высокий, закрывает талонидный бассейн с лингвальной стороны. Прецингулид, лабиальный и лингвальный цингулиды широкие, постцингулид тонкий. Лабиальная входящая складка (гипофлексид) на M1–M2 открывается заметно дорсальнее цингулида.
Судя по альвеолам, M3 относительно короткий и узкий.
Размеры в мм. Экз. ПИН, № 5826/560: длина P4–M3 – 4.6; A3: L – 0.85; W – 0.65; P4: BL – 1.55; LL – 1.15; PE – 0.9; AW – 1.15; PW – 1.4; M1: BL – 1.5; LL – 1.45; PE – 1.12; AW – 1.4; PW – 1.7; M2: BL – 1.3; LL – 1.25; PE – 1.02; AW – 1.5; PW – 1.35; M3: L – 0.65; W – 1.15; PE-индекс: P4 – 0.5; M1 – 0.317; M2 – 0.25.
Экз. ПИН, № 5826/561, M1: BL – 1.35; LL – 1.35; PE – 1.05; AW – 1.45; PW – 1.65; PE-индекс – 0.286.
Экз. ПИН, № 5826/562: длина нижнечелюстной кости (расстояние от вершины I1 до заднего края сочленовного мыщелка) – 10.5; расстояние от переднего края P4 до заднего края сочленовного мыщелка – 8.1; длина I1–M3 (до дистального края альвеолы заднего корня M3) – 6.9; длина P4–M3 – 4.5; I1: L – 0.8; W – 0.6; BH – 3.15; BL – 2.25; P4: L – 1.0; W – 0.7; M1: L – 1.5; TRW – 0.75; TAW – 0.85; M2: L – 1.4; TRW – 0.75; TAW – 0.8; альвеолярная длина M3 – 1.0; расстояние от подбородочного отверстия до верхней точки дуги перед угловым отростком – 4.7; высота горизонтальной ветви под M1 – 1.3, под M2 – 1.15; высота венечного отростка от верхней точки дуги перед угловым отростком – 3.7; сочленовный мыщелок: высота – 1.4; длина верхней фасетки – 0.9; длина нижней фасетки – 1.35.
Сравнение. Существенно мельче C. salenskii (Kastschenko, 1907), C. smithii Thomas, 1911, C. parca Allen, 1923 (= C. lowei Osgood, 1932), C. furva Anthony, 1941, C. hybsibia (de Winton et Styan, 1899) (= C. lamula Thomas, 1912), C. hoffmanni Chen et al., 2017 и C. sodalis Thomas, 1913, но крупнее C. parva Allen, 1923 и C. bohlini (Young, 1934). От C. hypsibia и C. furva также отличается относительно более мелким выступом гипокона и менее глубокой задней выемкой P4–M2.
Замечания. Сравнение размеров проведено в основном по длине P4–M3 (минимум – среднее – максимум в мм: C. smithii: 5.47–5.78–6.1, 9 экз.; C. parca: 5.57–5.71–5.84, 10 экз.; C. furva: 5.07–5.39–5.56, 4 экз.; C. hypsibia: 4.66–5.27–5.86, 52 экз.; C. hoffmanni: 4.59–4.82–5.03, 12 экз.; C. sodalis: 4.82–4.85–4.88, 2 экз.; C. caovansunga: 4.57–4.65–4.77, 3 экз.; C. parva: 3.57–3.94–4.12, 29 экз.) и длине I1–M3 (минимум – среднее – максимум в мм: C. smithii: 8.3–8.78–9.2, 9 экз.; C. parca: 7.96–8.15–8.34, 10 экз.; C. furva: 7.88–8.06–8.26, 4 экз.; C. hypsibia: 7.31–8.1–9.12, 53 экз.; C. hoffmanni: 7.19–7.5–7.67, 12 экз.; C. caovansunga: 7.12–7.25–7.39, 3 экз.; C. sodalis: 6.7–6.95–7.2, 2 экз.; C. parva: 5.7–5.95–6.23, 28 экз.) [см. 1].
Дополнительные сравнительные данные по размерам представителей Chodsigoa из Вьетнама (в мм): длина I1–M3: C. hoffmanni: 7.21–7.4–7.51, 3 экз. [2, 3]; 7.46–7.55–7.66, 3 экз. [3]; C. caovansunga: 6.93–7.04–7.25, 5 экз. [2, 3]; 7.23–7.31–7.4, 4 экз. [3]; длина нижнечелюстной кости: C. hoffmanni: 11.79–11.87–11.92, 3 экз. [2, 3]; 11.95–12.16–12.51, 3 экз. [3]; C. caovansunga: 10.57–10.87–11.1, 5 экз. [2, 3]; 11.03–11.19–11.39, 4 экз. [3]; высота нижнечелюстной кости в венечном отростке: C. hoffmanni: 4.05–4.17–4.28, 3 экз. [2, 3]; 4.12–4.26–4.34, 3 экз. [3]; C. caovansunga: 3.72–3.96–4.25, 5 экз. [2, 3]; 3.54–3.83–4.09, 4 экз. [3].
Длина I1–M3C. caovansunga из Китая (1 экз.) – 7.39 мм [4].
У C. bohlini расстояние от переднего края P4 до заднего края сочленовного мыщелка – 7.5 мм; длина P4–M3 – 4.2 мм; высота нижнечелюстной кости в венечном отростке – 3.3 мм [9].
Морфологические признаки и размеры изученных экземпляров из пещеры Там-Хай позволяют сделать вывод об их принадлежности к C. caovansunga. Это первые ископаемые находки данного вида.
Распространение. Средний плейстоцен северного Вьетнама; ныне в Южном Китае и северном Вьетнаме.
Материал. Из пещеры Там-Хай (обособленная северная галерея, 21°55′52.5″ с.ш., 106°25′4.1″ в.д.) фрагмент левой верхнечелюстной кости с A3, P4–M3 (экз. ПИН, № 5826/560), изолированный левый M1 (экз. ПИН, № 5826/561) и фрагмент левой нижнечелюстной кости с I1, P4–M2 и восходящей ветвью (экз. ПИН, № 5826/562).
Chodsigoa hoffmanni Chen et al., 2017
Chodsigoa hoffmanni: [1, с. 706, рис. 6].
Голотип – Куньминский институт зоологии, KIZ 032249, череп, скелет и шкурка взрослой самки; Китай, Юньнань, гора Айлао (Ailao); современный.
Описание (рис. 3). Размеры средние для рода. P4–M1 с глубокой, но плавной выемкой заднего края.
P4 относительно высокий и длинный. Вершина паракона отчетливо пигментированная с антеролабиальной стороны. Парастиль крупный, высокий, сильно выступающий вперед, связан с передним основанием паракона резким гребнем. Метастилярный выступ крупный, ориентирован постеролабиально. Гребень паракристы высокий, очень широкий и мощный. Протокон хорошо выраженный, конусовидный, соединен с лингвальным основанием парастиля четким гребнем с маленькой вырезкой. Постпротокристы нет. Выступ гипокона сильно вытянут постеролингвально, по его лингвальному краю протягивается резкий гребень. Лабиальный цингулюм тонкий, постцингулюм отчетливый, с заметным расширением в наиболее лабиальной части.
M1–M2 относительно длинные и узкие, с мощным W-образным эктолофом. Паракон немного меньше и ниже метакона. Протокон связан с антеролингвальным основанием паракона высоким гребнем препротокристы. Выступ гипокона сильно выдается назад (на M2 в меньшей степени, чем на M1). Помимо лабиального цингулюма и постцингулюма имеется короткий лингвальный цингулюм, расположенный на уровне задней части протокона. На M1 выступ парастиля поперечно короткий, ориентирован антеролабиально, на M2 – более удлиненный и направлен строго лабиально. На M2 задняя выемка менее глубокая, чем на M1, лингвальный цингулюм более короткий и менее отчетливый.
Судя по сохранившимся альвеолам, M3 сильно редуцирован, вытянут поперечно.
Размеры в мм. Экз. ПИН, № 5792/525: длина P4–M2 – 4.5; длина P4–M3 – ок. 5.1; P4: BL – 1.75; LL – 1.3; PE – 1.05; AW – 1.05; PW – 1.85; M1: BL – 1.65; LL – 1.65; PE – 1.3; AW – 1.65; PW – 1.90; M2: LL – 1.45; PE – 1.15; AW – 1.65; PE-индекс: P4 – 0.452; M1 – 0.269.
Сравнение. Существенно мельче C. salenskii, C. smithii и C. parca, но заметно крупнее C. parva, C. caovansunga, C. sodalis и C. bohlini. От C. hypsibia и C. furva отличается более мелким выступом гипокона и менее глубокой эмаргинацией P4–M2.
Замечания. Сравнение размеров проведено в основном по длине P4–M3 [1] (см. выше). При этом учтены данные о значительной изменчивости общей длины верхнего зубного ряда (I1–M3) в разных популяциях C. hoffmanni (минимум – среднее – максимум в мм): 7.68–8.11–8.31, 12 экз. [1]; 8.1–8.2–8.4, 6 экз.; 8.31–8.44–8.52, 3 экз. [2, 3]; 8.46–8.57–8.71, 3 экз. [3]; 8.29–8.32–8.37, 3 экз. [5]; 8.2, 1 экз. [6]. В целом размеры и строение верхних щечных зубов позволяют отнести экземпляр из пещеры Лангчанг к C. hoffmanni. Это первая идентификация ископаемых остатков указанного вида.
Описанный образец является самой южной ископаемой находкой Chodsigoa из известных. Данная находка свидетельствует, что во Вьетнаме в плейстоцене представители этого рода распространялись на юг значительно дальше (приблизительно на 250 км), чем в настоящее время [см. 1, рис. 1].
Распространение. Верхний плейстоцен северного Вьетнама; ныне в Центральном и Южном Китае и северном Вьетнаме.
Материал. Из пещеры Лангчанг (20°20′29″ с.ш., 105°12′21″ в.д.) фрагмент левой верхнечелюстной кости с P4–M2 и альвеолами M3 (экз. ПИН, № 5792/525).
Список литературы
Chen Z.-Z., He K., Huang C., et al. Integrative systematic analyses of the genus Chodsigoa (Mammalia: Eulipotyphla: Soricidae), with descriptions of new species // Zoological Journal of the Linnean Society. 2017. V. 180. P. 694–713.
Lunde D.P., Musser G.G., Nguyen Truong Son. A survey of small mammals from Mt. Tay Con Linh II, Vietnam, with the description of a new species of Chodsigoa (Insectivora: Soricidae) // Mammal Study. 2003. V. 28. P. 31–46.
Saito H., Bui Tuan Hai, Ly Ngoc Tu, et al. Shrews (Mammalia: Eulipotyphla: Soricidae) from Mt. Tay Con Linh, Ha Giang Province, northeast Vietnam // Bulletin of National Museum of Nature and Science. Ser. A. 2021. V. 47. № 1. P. 43–53.
He K., Deng K., Jiang X.-L. First record of Van sung’s shrew (Chodsigoa caovansunga) in China // Zoological Research. 2012. V. 33. № 5. P. 542–544.
Liu Z., Jiang X.-T., Wang Q.-Q., et al. Hoffmann’s long-tailed shrew (Chodsigoa hoffmanni) found in Bijie, Guizhou Province, China // Chinese Journal of Zoo-logy. 2021. V. 56. № 5. P. 776–781.
Lei B.-Y., Cui J.-F., Yue Y., et al. Hoffmann’s long-tailed shrew (Chodsigoa hoffmanni) found in Xingshan, Hubei Province, China // Chinese Journal of Zoology. 2019. V. 54. № 6. P. 820–824.
Repenning C.A. Subfamilies and genera of the Soricidae // Geological Survey Professional Paper. 1967. № 565. P. 1–74.
Flynn L.J., Wu W.-Y. The Lipotyphla of Yushe Basin. In: Flynn L.J., Wu W.-Y., eds. Late Cenozoic Yushe Basin, Shanxi Province, China: geology and fossil mammals. V. II. Small mammal fossils of Yushe Basin. Dordrecht: Springer; 2017. P. 11–26.
Young C.C. On the Insectivora, Chiroptera, Rodentia and Primates other than Sinanthropus from Locality 1 at Choukoutien // Palaeontologia Sinica. Ser. C. 1934. V. 8. № 3. P. 30–41.
Jin C.-Z., Zhang Y.-Q., Wei G.-B., et al. Rodentia Bowdich, 1821. In: Jin C., Liu J., eds. Paleolithic site – the Renzidong Cave, Fanchang, Anhui Province. Beijing: Science Press; 2009. P. 166–220, 401–412.
Ye X., Fang Q., Hou L., et al. Fossil vertebrates. In: Wushan hominid site. Beijing: China Ocean Press; 1991. P. 24–149.
Zheng S. Rodentia. In: Zheng S., ed. Jianshi hominid site. Beijing: Science Press; 2004. P. 121–181, 387–394.
Wu X., Wang Z. Small mammalian remains from the Wazhuozui site in Fengdu, Chongqing // Acta Anthropologica Sinica. 2018. V. 37. № 3. P. 452–466.
Zheng S. Micromammals from the Hexian man locality // Vertebrata PalAsiatica. 1983. V. 21. № 3. P. 230–240.
Zheng S.-H., Zhang Z.-Q. Late Miocene–Early Pleistocene biostratigraphy of the Leijiahe area, Lingtai, Gansu // Vertebrata PalAsiatica. 2001. V. 39. № 3. P. 215–228.
Wei G., Huang W., Boëda E., et al. Recent discovery of a unique Paleolithic industry from the Yumidong Cave site in the Three Gorges region of Yangtze River, southwest China // Quaternary International. 2017. V. 434. Pt. A. P. 107–120.
Zhao L., Zhang L., Du B., et al. New discovery of human fossils and associated mammal fauna from Mawokou Cave in Bijie, Guizhou Province of Southern China // Acta Anthropologica Sinica. 2016. V. 35. № 1. P. 24–35.
Zeitoun V., Chinnawut W., Debruyne R., et al. A sustainable review of the Middle Pleistocene benchmark sites including the Ailuropoda–Stegodon faunal complex: the proboscidean point of view // Quaternary International. 2016. V. 416. P. 12–26.
Лопатин А.В., Мащенко Е.Н., Вислобокова И.А., и др. Плейстоценовые млекопитающие из пещеры Лангчанг (Вьетнам): новые данные // Доклады Российской академии наук. Науки о жизни. 2021. Т. 496. № 1. С. 5–9.
Reumer J.W.F. Ruscinian and early Pleistocene Soricidae (Insectivora, Mammalia) from Tegelen (The Netherlands) and Hungary // Scripta Geologica. 1984. V. 73. P. 1–173.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни