

Доклады Российской академии наук. Науки о жизни, 2022, T. 503, № 1, стр. 113-116
Первые данные о следах микроизноса неокклюзиальной поверхности резцов пещерных медведей Урала
Д. О. Гимранов 1, 2, *, С. В. Зыков 1, П. А. Косинцев 1
1 Институт экологии растений и животных УрО РАН
Екатеринбург, Россия
2 Уральский федеральный университет
им. первого президе́нта России Б.Н. Ельцина
Екатеринбург, Россия
* E-mail: djulfa250@rambler.ru
Поступила в редакцию 29.11.2021
После доработки 21.12.2021
Принята к публикации 21.12.2021
- EDN: WPGNNM
- DOI: 10.31857/S2686738922020093
Аннотация
Проведен сравнительный анализ микроизноса неокклюзиальной поверхности резцов (I1, I2) малого (Ursus ex gr. savini-rossicus) и уральского (Ursus kanivetz) пещерных медведей из плейстоценовых отложений Среднего и Южного Урала. Выявлены межвидовые различия в качественных характеристиках элементов микроизноса резцов. У малого пещерного медведя отмечены более грубые повреждения на неокклюзиальной поверхности резцов. С учетом специфики микроизноса неокклюзиальных поверхностей зубов на основании полученных данных сделаны предположения о различиях в трофической специализации изученных видов.
Согласно современным морфологическим и молекулярным данным на Урале в позднем плейстоцене обитало два вида пещерных медведей [1, 2]. Это большой, или уральский, пещерный медведь (Ursus kanivetz Verestchagin, 1973) и малый, или русский, пещерный медведь (U. rossicus Borissiak, 1930). На сегодняшний день группа малых пещерных медведей нуждается в ревизии [2]. В данной работе мы рассматриваем малого пещерного медведя как принадлежащего к группе savini-rossicus (U. ex gr. savini-rossicus).
Малый пещерный медведь по сравнению с большим пещерным медведем является плохо изученным видом. Работы, касающиеся особенностей экологии малого пещерного медведя, очень редки [3–6], исследования диеты единичны [7]. Одним из методов изучения особенностей питания млекопитающих является анализ следов микроизноса зубов. В настоящее время на основе анализа микроизноса зубов проведены реконструкции диеты представителей различных отрядов млекопитающих [8–12]. Особый интерес в данном направлении представляет анализ микроизноса неокклюзиальных поверхностей зубов. Исследования микроизноса буккальных и лабиальных поверхностей зубов приматов показали важность данного подхода не только к выяснению состава диеты, но и реконструкции пищевого поведения [7, 13–15]. Пищевые предпочтения ряда видов медведей также изучены с использованием метода анализа микроизноса зубов [7, 10, 12, 16]. Однако сведения о микроповреждениях и износе резцов пещерных медведей в литературе крайне скудны.
Целью данной работы была оценка качественных и количественных характеристик микроизноса лабиальных поверхностей резцов у малого и большого пещерных медведей с территории Уральского региона. Для анализа микроследов нами были выбраны верхние первый (I1) и второй (I2) резцы. В изолированном состоянии отличить первый от второго резца у пещерных медведей достаточно затруднительно. Эти зубы прорезаются в одно время и стираются синхронно, поэтому далее рассматриваем эти резцы вместе. Были выбраны резцы со среднем уровнем стертости, когда достаточно сильно обнажается дентин верхней части коронки, но зуб стерт не более чем на 1/3. Исследовано 10 верхних резцов (I1–2), принадлежащие малому пещерному медведю и происходящих из позднеплейстоценового местонахождения пещера Иманай. Резцы большого пещерного медведя (n = 11 экз.) происходят из пещер Тайн (n = 2), Игнатьевская (n = 3), Заповедная (n = 3) и Аша 1 (n = 3). Пещера Тайн расположена на Среднем Урале, остальные пещеры – на Южном Урале. На основе радиоуглеродных дат и биостратиграфических данных все местонахождения с остатками пещерных медведей датируются началом и серединой (морская изотопная стадия (МИС) 5 – МИС 3) позднего плейстоцена [2].
Анализ микроизноса проводили на лабиальной поверхности резцов на микрофотографиях, полученных при увеличении ×30 на электронном сканирующем микроскопе TESCAN VEGA3. Количественный и качественных анализ элементов микроизноса зубов проводился на площади 6 мм2 с использованием полуавтоматической программы Microwear 4.02 (Ungar, 1994–2002, США) (рис. 1). Оценку межвидовых различий микроизноса проводили с помощью дисперсионного анализа (ANOVA), используя программу STATISTICA 8.0. (“StatSoft, Inc.”, 1984–2007, США).
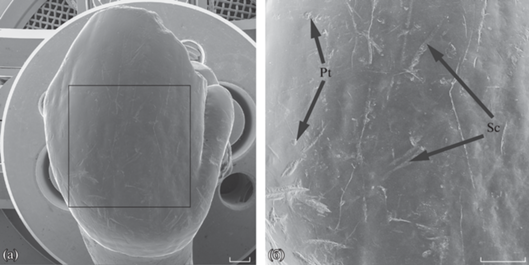
Рис. 1.
Общий вид и расположение учетной площади (а) на лабиальной поверхности резца (на примере U. ex gr. savini-rossicus). Ямки (Pt) и царапины (Sc) на учетной площади (б). Масштаб 1 миллиметр.
В результате проведенного анализа не выявлено значимых межвидовых различий в количестве микроповреждений на лабиальной поверхности резцов у двух видов пещерных медведей (рис. 2). Наблюдается общий тренд к уменьшению количества элементов микроизноса эмалевой поверхности у большого пещерного медведя. Среднее количество ямок/царапин у малого медведя составило 15.8/54.6, у большого пещерного медведя 7/37.2 соответственно (рис. 2б), однако различия и по числу ямок (F(1; 19) = 4.13, p > 0.05) так и по числу царапин (F(1;19) = 3.88, p > 0.05) оказались незначимы.
Рис. 2.
Средние значения и доверительный интервал ширины (а) и общего числа (б) ямок и царапин лабиальной поверхности резцов малого и большого пещерных медведей.
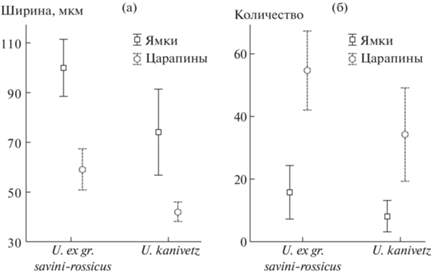
Межвидовые различия были выявлены по ширине ямок/царапин на эмали. Среднее значение ширины ямок/царапин у малого пещерного медведя составило 99.9/59.1, у большого пещерного медведя 74.11/42.0 микрометров соответственно (рис. 2а). Различия по ширине ямок (F(1;19) = 7.38, p < 0.05) и царапин (F(1;19) = 18.74, p < 0.001) статистически достоверны. На лабиальной поверхности резцов малого пещерного медведя повреждения носят более грубый характер, ширина и ямок и царапин превышает значения аналогичных элементов у большого пещерного медведя (рис. 2а).
В результате проведенного анализа микроповреждений эмали зубов, который широко используется в палеореконструкциях трофических особенностей наземных млекопитающих, установлены значимые статистические различия по размерам микроследов на неокклюзиальной поверхности резцов большого и малого пещерных медведей.
На данный момент интерпретация микроизноса зубов медведей имеет дискуссионный характер. На основе анализа следов микроизноса окклюзиальных поверхностей моляров делались выводы о большей плотоядности пещерного медведя по сравнению с бурым [17], всеядности [18], смешанной диеты [19]. Учитывая последние данные комплексного анализа палеодиеты на основе изотопного анализа и микроизноса моляров, мы придерживаемся гипотезы о растительной диете пещерных медведей [20].
Ранее было отмечено, что количество грубых повреждений на окклюзиальной поверхности моляров у большого пещерного медведя больше по сравнению с малым, что может свидетельствовать о различиях в составе кормов [7]. Учитывая полученные нами результаты по размерным характеристикам повреждений на резцах данных видов, можно предположить, что при добывании корма на резцы малого пещерного медведя оказывалось более грубое воздействие, что возможно связано как с субстратом, на котором произрастали кормовые растения, так и со смещением растительноядной диеты малого пещерного медведя в сторону ризофагии.
Различия характеристик элементов микроизноса лабиальных поверхностей резцом изученных видов в совокупности с данными по микроизносу окклюзиальных поверхностей моляров [7] свидетельствуют о разнице в составе кормов и/или пищевом поведении данных видов. Для более полного описания различий в питании и пищевых предпочтениях пещерных медведей необходимо проведение комплексного анализа микро- и макроизноса поверхностей нижних резцов, клыков и щечных зубов.
Список литературы
Barlow A., Paijmans J.L.A., Federica A., et al. Middle Pleistocene genome calibrates a revised evolutionary history of extinct cave bears // Current Biology. 2021. V. 31. № 8 P. 1771–1779.
Гимранов Д.О., Косинцев П.А. Пещерные медведи (Ursus spelaeus sensu lato) Урала // Палеонтологический журнал. 2022. № 1. С. 97–106.
Борисяк A.A. Новая раса пещерного медведя из четвертичных отложений Сев. Кавказа // Труды палеозоологического института. 1932. Т. 1. С. 137–201.
Vereschagin N., Baryshnikov G. Small cave bear Ursus (Spelearctos) rossicus uralensis from the Kizel Cave in the Ural (Russia) // Geoloski zbornik, Liubljana. 2000. V. 15. P. 53–66.
Барышников Г.Ф. Семейство медвежьих (Carnivora, Ursidae). СПб.: Наука; 2007.
Spassov N., Hristova L., Ivanova S., Georgiev I. First record of the “small cave bear” in Bulgaria and the taxonomic status of bears of the Ursus savini Andrews – Ursus rossicus Borissiak group // Fossil Imprint. 2017. V. 73. P. 275–291.
Ramirez Pedraza I., Baryshnikov G.F., Prilepskaya N.E., Belyaev R.I., et al. Paleodiet and niche partitioning among the easternmost European cave bears based on tooth wear analysis // Historical Biology. 2021. P. 1–9.
Ungar P.S. Dental microwear of European Miocene catarrhines: evidence for diets and tooth use // J. Hum. Evol. 1996. V. 31. P. 335–366.
Nelson S., Badgley C., Zakem E. Microwear in modern squirrels in relation to diet // Palaeontol. Electron. 2005. V. 8. № 1. P. 14A: 15 p.
Pinto-Llona A.C. Macrowear and occlusal microwear on teeth of cave bears Ursus spelaeus and brown bears Ursus arctos: Inferences concerning diet // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013. V. 370. P. 41–50.
Merceron G., Schulz E., Kordos L., et al. Paleoenvironment of Dryopithecus brancoi at Rudabánya, Hungary: evidence from dental meso-and micro-wear analyses of large vegetarian mammals // J. Hum. Evol. 2007. V. 53. № 4. P. 331–349.
Peigne S., Merceron G. Palaeoecology of cave bears as evidenced by dental wear analysis: a review of methods and recent findings // Historical Biology. 2019. V. 31. № 4. P. 448–460.
Ungar P.S., Teaford M.F. Preliminary examination of non-occlusal dental microwear in anthropoids: implications for the study of fossil primates // Am. J. Phys. Anthropol. 1996. V. 100. № 1. P. 101–113.
Perez-Perez A., Espurz V., Bermudez de Castro J.M., et al. Non-occlusal dental microwear variability in a sample of Middle and Late Pleistocene human populations from Europe and the Near East // J. Hum. Evol. 2003. V. 44. № 4. P. 497–513.
Romero A., Galbany J., De Juan J., et al. Brief communication: Short- and long-term in vivo human buccal-dental microwear turnover // Am. J. Phys. Anthropol. 2012. V. 148. № 3. P. 467–472.
Pappa S., Schreve D.C., Rivals F. The bear necessities: A new dental microwear database for the interpretation of palaeodiet in fossil Ursidae // Palaeogeography, Palaeoclimatology, Palaeoecology. 2019. V. 514. P. 168–188.
Pinto Llona A.C. Comparative dental microwear analysis of cave bears Ursus spelaeus Rosenmuller, 1794 and brown bears Ursus arctos Linnaeus, 1758. // Sci. Annals. Geol. School. 2006. V. 98. P. 103–108.
Donohue S.L., DeSantis L.R.G., Schubert B.W., et al. Was the Giant Short-Faced Bear a Hyper-Scavenger? A New Approach to the Dietary Study of Ursids Using Dental Microwear Textures // PLOS One. 2013. V. 8. № 10. P. e77531.
Jones D.B., Desantis L.R.G. Dietary ecology of ungulates from the La Brea tar pits in southern California: a multi-proxy approach // Palaeogeography, Palaeoclimatology, Palaeoecology. 2017. V. 466. P. 110–127.
Bocherens H. Isotopic insights on cave bear palaeodiet // Historical Biology. 2019. V. 31. № 4. P. 410–421.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни