

Доклады Российской академии наук. Науки о жизни, 2022, T. 503, № 1, стр. 127-132
Сезонная динамика и закономерности поворотной активности участков ствола деревьев сосны обыкновенной
С. А. Шавнин 1, *, Д. Ю. Голиков 1, А. А. Монтиле 1, **
1 Федеральное государственное бюджетное учреждение науки Ботанический сад Уральского отделения Российской академии наук
Екатеринбург, Россия
* E-mail: sash@botgard.uran.ru
** E-mail: org17@mail.ru
Поступила в редакцию 10.11.2021
После доработки 27.11.2021
Принята к публикации 27.11.2021
- EDN: UFPGXG
- DOI: 10.31857/S2686738922020196
Аннотация
С целью углубления представлений о природе явления закручивания стволов деревьев и его роли в процессах роста и морфогенеза, в течение двух лет исследовали сезонные динамики и взаимосвязи характеристик углов поворота участков внешних слоев заболони у стволов деревьев сосны обыкновенной (Pinus sylvestris L.) в естественном насаждении. Повороты существуют у всех деревьев в течение всего вегетационного периода. Их характеристики не зависят от экспозиции и отличаются у разных участков ствола. Величины углов и направления поворотов смежных участков ствола положительно коррелируют. Доминируют направления поворота против часовой стрелки, в результате чего суммарный угол поворота может увеличиваться с течением времени. Направления и величины сил, вызывающих повороты внешних участков заболони ствола дерева, объясняются, по-видимому, градиентами механических напряжений, возникающих между соседними частями ксилемы в результате неоднородности их увлажнения.
У большого числа видов древесных растений, как в течение суток, так и более длительного времени, наблюдаются изменения диаметра ствола [1, 2]. Этот эффект связан с процессами радиального роста и набухания-сжатия тканей, сопровождающими вертикальный транспорт воды, минеральных веществ и ассимилятов по ксилеме [1] и флоэме [3, 4]. Следует отметить, что восходящие потоки распределены по ксилеме в сечении ствола неравномерно [5, 6]. Например, у деревьев сосны обыкновенной наблюдается их спиральное движение [7]. Оно объясняется упорядоченным в пространстве расположением ветвей, задающим в ходе транспирации формирование в стволе градиентов водного потенциала – основного фактора, определяющего распределение влаги в растениях [8–10]. При этом изменение оводненности разных участков тканей приводит к их набуханию-сжатию и образованию соответствующих механических напряжений в древесине [11]. В связи с неравномерным распределением в объеме ствола участков с различной оводненностью [12], градиенты действующих сил не полностью совпадают с его вертикальной осью симметрии и могут приводить к тангенциальным смещениям в многослойной структуре сечения ствола [13].
В ходе исследований динамических процессов, участвующих в регуляции роста и морфогенеза дерева, ранее было описано явление поворотов ствола у деревьев сосны (Pinus sylvestris L.) и ели (Picea obovata Ldb.) и предложена качественная модель связи поворота ствола с процессами камбиального роста [14], объясняющая закономерный характер явления и образование косослоя древесины и закрученности стволов деревьев. Однако в модели отсутствует указание на природу сил, определяющих направление, углы и длительность формирования косослоя.
В связи с необходимостью выяснения причин наблюдаемого явления, цели настоящей работы включали: 1) определить величины и направления поворота ствола (участков внешних слоев заболони) в течение вегетационного периода; 2) оценить характер и степень взаимосвязи этих характеристик поворотной активности у разных сторон ствола. Для достижения целей были проведены: а) исследования сезонной динамики поворотной активности стволов деревьев сосны обыкновенной, произрастающих в насаждении естественного происхождения; б) анализ закономерностей сезонных изменений направлений и величин углов поворота участков стволов разной экспозиции; в) разработка дополнений к предложенной ранее качественной модели закручивания ствола дерева.
Объектом исследований являлись деревья сосны обыкновенной, произрастающие на территории дендрария Ботанического сада УрО РАН (г. Екатеринбург, РФ), относящейся к южно-таежной подзоне Среднеуральской низкогорной провинции Уральской горнолесной области [15]. При обследовании древостоев использовали метод пробных площадей (ПП). На ПП, заложенной в соответствии с ОСТ 56-69-83, определяли базовые таксационные показатели. Насаждения имеют естественное происхождение, тип леса – С.ртр., состав – 10С, ед.Б, VII класс возраста, III класс бонитета, полнота 0.8, средний диаметр 39.7 см, средняя высота 25.4 м. Общее количество деревьев на ПП составляет 481, площадь ПП 2.1 га.
Для проведения исследований были отобраны 36 средних по диаметрам и высотам деревьев. Величины и направления поворота определяли у участков внешних слоев заболони стволов отдельных деревьев с четырех разных экспозиций (Ю-В, Ю-З, С-В, С-З) на протяжении 2017–2018 гг. Угол поворота определяли 5–6 раз за период “весна – осень” по величине хорды окружности, описываемой рычагом измерительного устройства в горизонтальной плоскости (Шавнин и др., 2018). Общее количество определений в каждый момент времени составляло 144. С целью выявления наиболее общих закономерностей анализировали величины суммарных углов поворотов стволов за отдельные периоды измерений, которые определяли усреднением всех измерений (независимо от номера дерева и экспозиции) в данный момент времени с учетом знака (направления поворота), либо по модулю.
Для анализа рядов сезонной динамики поворотов ствола применяли стандартные методы многомерной статистики. Расчеты проводились в программах Statistica 8.0 (StatSoft Inc., 2007), Excel (Microsoft, 2003) и “Классан” [16] .
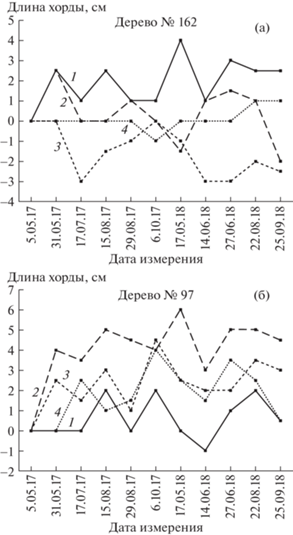
Ввиду большого объема данных, результаты измерений углов поворотов стволов у отдельных деревьев рассмотрены на примере двух типичных экземпляров. В 2017 г. у дерева № 162 (рис. 1а) вращение в мае происходило в сторону “+” (против часовой стрелки) у Ю-З и С-З экспозиций и отсутствовало – у Ю-В и С-В. В период 05.05.2017 г.–17.07.2017 г. направление поворота изменилось на противоположное у трех экспозиций и отсутствовало – у Ю-В. В дальнейшем углы поворота у разных экспозиций изменялись по направлению и величине также не согласованно или частично согласованно на протяжении всех двух лет исследований. Следует отметить, что у данного дерева на протяжении двух лет наблюдаются как “+”, так и “–” – повороты. У дерева № 97 (рис. 1б) у экспозиций С-З, С-В и Ю-В доминируют “+” – повороты, а у Ю-З доминирование не наблюдается.
Рис. 1.
Сезонные динамики (траектории) величин углов поворота (длин хорд окружностей, описываемых рычагом измерительного устройства) участков стволов разных экспозиций деревьев № 162 (а) и № 97 (б) в 2017–2018 гг. 1–4: Ю-З, С-З, С-В, Ю-В экспозиции соответственно; положительные изменения соответствуют повороту против часовой стрелки, отрицательные – по часовой стрелке.
Проведенный для всех деревьев корреляционный анализ сезонных динамик углов поворота смежных сторон ствола между собой показал, что их взаимосвязи достоверны только в половине случаев. Величины коэффициентов корреляции варьируют в широком диапазоне (0.125–0.918, p < < 0.1) и есть единичные случаи нулевых и отрицательных корреляций. У несмежных экспозиций ствола величины углов поворота, как правило, не взаимосвязаны. Величины и направления поворотов каждой стороны дерева изменяются в течение всего периода весна – осень.
Ввиду отсутствия наблюдаемой у большинства деревьев взаимосвязи между характеристиками вращения разных сторон ствола, годовые сезонные динамики по каждой экспозиции (траектории) кластеризовали независимо от экспозиции у всех деревьев с помощью евклидовой метрики (метод Уорда). Анализ данных 2017 г. (рис. 2) позволяет установить четыре типа динамик, которые наблюдаются у разных деревьев с различной частотой. У одного дерева встречается 4 типа траекторий, у 21 дерева – 3 типа, у 12–2 типа и у 2–1 тип. Аналогичный анализ, проведенный для данных 2018 г., подтвердил выявленные закономерности, свидетельствующие о неслучайном характере участвующих в формировании поворота процессов.
Рис. 2.
Дендрограмма сходства 144 траекторий поворотов участков стволов четырех экспозиций 36 деревьев в 2017 г. I–IV – выделенные типы траекторий.

Далее была рассмотрена сезонная динамика суммарного угла поворота ствола в течение двух лет (рис. 3), расчет которой проводили с учетом направления вращения для выборки, включающей все деревья. Статистически значимое вращение стволов наблюдается уже в мае в 2017 г. В период 31.05.2017 г. – 15.06.2017 г. происходит увеличение поворотной активности с последующей ее относительной стабилизацией. В 2018 г. активация вращения произошла позже (во второй половине июня), а стабилизация – в сентябре. Следует отметить, что в течение апреля – начале мая 2018 г. [17] суммы средних за декаду эффективных температур были на 20–30°С ниже, чем в 2017 г.
Рис. 3.
Сезонные динамики величин средних суммарных углов поворота участков стволов четырех экспозиций 36 деревьев в 2017–2018 гг. Приведены длины хорд окружностей, описываемых рычагом измерительного устройства при повороте; 1, 2 – величины рассчитаны с учетом и без учета направления поворота соответственно; указаны стандартные ошибки среднего.

Суммарный угол поворота ствола, рассчитанный без учета направлений величин его отдельных составляющих (рис. 3), возрастает уже в мае 2017 г. В дальнейшем его величина стабилизируется, дополнительно увеличивается в середине августа, после чего вращение стволов приостанавливается. В мае 2018 г. наблюдалось небольшое, а в августе – сентябре – достоверное увеличение суммарного угла поворота. Отличие двух описанных динамик в мае 2017 г. объясняется значительным количеством обратных по направлению поворотов в этот период. Проведенный двухэтапный анализ позволяет заключить, что поворотная активность деревьев сосны начинается в мае – июне, что соответствует периоду активного камбиального и апикального роста, и сохраняется до августа – сентября.
Регрессионный анализ величин углов поворота всех изученных деревьев в пределах одной экспозиции (табл. 1) показал, что в течение 2017–2018 гг. существуют положительные парные корреляции между значениями, полученными в интервале времени до двух месяцев. Эта закономерность указывает на то, что в пределах одной экспозиции вызывающие вращение силы могут изменяться по модулю, однако направление их результирующей сохраняется. Согласно данным 2017 г., в период измерений май – июль степень взаимосвязи меньше (коэффициент детерминации R2 = 0.350), чем в последующие три (R2 = 0.712, 0.648 и 0.670 соответственно). Более слабая корреляция углов поворота в этот период свидетельствует о наличии весной и в начале лета дополнительных факторов, снижающих упорядоченность во времени локализаций и величин возникающих в древесине механических напряжений. Одним из них, по-видимому, является отличие в 2017 г. сроков активации физиологических процессов у разных деревьев в древостое. По сравнению с 2017 г., в 2018 г. корреляция углов поворота за смежные по времени периоды в мае – июне выше, и сохраняется в течение всего вегетационного сезона (R2 = 0.576–0.690).
Таблица 1.
Результаты регрессионного анализа величин углов поворотов участков ствола (36 деревьев, 4 экспозиции) за два смежных по времени срока. Приведены параметры линейных регрессий и их коэффициенты детерминации; x, y – величины углов поворота в предшествующий и последующий моменты времени измерений соответственно
| Даты измерений поворотов | Параметры регрессии | ||
|---|---|---|---|
| R2 | Уравнение | ||
| 31.05.2017 | 17.07.2017 | 0.350 | y = 0.579x + 0.448 |
| 17.07.2017 | 15.08.2017 | 0.712 | y = 0.856x + 0.991 |
| 15.08.2017 | 29.08.2017 | 0.648 | y = 0.842x + 0.049 |
| 29.08.2017 | 06.10.2017 | 0.670 | y = 0.867x + 0.121 |
| 17.05.2018 | 14.06.2018 | 0.576 | y = 0.770x + 0.228 |
| 14.06.2018 | 27.06.2018 | 0.624 | y = 0.764x + 0.734 |
| 27.06.2018 | 22.08.2018 | 0.690 | y = 0.877x + 0.388 |
| 22.08.2018 | 25.09.2018 | 0.660 | y = 0.799x + 0.518 |
Результаты анализа полученных данных указывают на то, что повороты внешних слоев заболони вызывают силы, возникающие в результате действия внутренних механических напряжений в древесине ствола. Последние являются, по-видимому, следствием разной оводненности смежных участков заболони, обусловленной неравномерным распределением в объеме ствола вертикальных и/или спиральных транспортных потоков воды. На существование напряжений, связанных с неравномерным распределением влаги в поперечном сечении ствола, указывают следующие закономерности и факты. Первое – установлено наличие взаимосвязи между характеристиками поворотов участков ствола смежных экспозиций. Оно связано, по-видимому, с несимметричной относительно центральной оси локализацией восходящих потоков влаги [18]. О пространственной упорядоченности этих потоков свидетельствует явление спирального движения воды в стволе молодых деревьев сосны обыкновенной [7]. Следует отметить, что наличие спиральных структур в древесине установлено у большого числа видов деревьев, включая сосну обыкновенную [19, 20]. Второе – поворотная активность участков ствола наблюдается в течение всего периода вегетации, что указывает на важную роль в ее формировании погодно-климатических факторов. Третье – установлены наличие связи между характеристиками поворотов участков ствола одной экспозиции в течение 1–2 мес и существование в сезонной динамике фактов изменения направления поворота на противоположное. Эти результаты свидетельствуют о том, что механические напряжения в заболони возникают на определенное время и могут изменяться как по величине, так и по направлению.
Таким образом, повороты внешних участков заболони на разных сторонах ствола существуют у всех деревьев сосны обыкновенной в течение всего вегетационного периода. Они возникают в результате смещений внешних слоев заболони, которые вызывают поворот исследуемого участка ствола вокруг вертикальной оси в горизонтальной плоскости в одном из двух тангенциальных направлений.
Величины и направления углов поворота разных участков ствола отличаются и не зависят непосредственно от экспозиции. Величины углов поворотов участков ствола с одинаковой экспозицией максимально коррелируют в течение смежных периодов времени (одного – двух месяцев). Существует взаимосвязь направлений поворотов у участков ствола смежных экспозиций. Доминируют направления поворота против часовой стрелки, в результате чего суммарный угол поворота может увеличиваться с течением времени. При этом на протяжении вегетационного периода наблюдаются изменения как величин углов, так и направлений поворота отдельных участков ствола. Установлено наличие четырех типов сезонных динамик поворотов участков ствола, указывающее на наличие упорядочивающего поворотную активность фактора.
Направления и величины сил, вызывающих повороты внешних участков заболони ствола дерева, объясняются, по-видимому, механическими напряжениями, возникающими между соседними частями ксилемы в результате неоднородности их увлажнения.
Список литературы
Sevanto S., Vesala T., Perämäki M., et al. Time lags for xylem and stem diameter variations in Scots pine // Plant, Cell Environ. 2002. V. 25, Iss. 8. P. 1071–1077.
Chan T., Berninger F., Kolari P., et al. Linking stem growth respiration to the seasonal course of stem growth and GPP of Scots pine // Tree Physiol. 2018. V. 38 (9). P. 1356–1370.
Sevanto S., Nikinmaa E., Riikonen A., et al. Linking xylem diameter variations with sap flow measurements // Plant Soil. 2008. V. 305, Iss. 1–2. P. 77–90.
Knoblauch M., Peters W. S. Münch, morphology, microfluidics – our structural problem with the phloem // Plant, Cell Environ. 2010. V. 33 (9). P. 1439–1452.
Matthes U., Kelly P.E., Ryan C.E., et al. The formation and possible ecological function of stem strips in Thuja occidentalis // Int. J. Plant Sci. 2002. V. 163 (6). P. 949–958.
Nadezdina N. Integration of water transport pathways in a maple tree: responces of sap flow to branch severing // Ann. For. Sci. 2010. V. 67. P. 107.
Кайбияйнен Л.К., Хари П. Сбалансированность системы водного транспорта у сосны обыкновенной. I. Пути движения влаги в ксилеме // Лесоведение. 1985. № 5. С. 23–28.
Sevanto S., Hölttä T., Holbrook N.M. Effects of the hydraulic coupling between xylem and phloem on diurnal phloem diameter variation // Plant, Cell Environ. 2011. V. 34 (4). P. 690–703.
Oberhuber W., Kofler W., Schuster R., et al. Environmental effects on stem water deficit in co-occurring conifers exposed to soil dryness // Int. J. Biometeorol. 2015. V. 59 (4). P. 417–426.
Venturas M.D., Sperry J.S., Hacke U.G. Plant xylem hydraulics: What we understand, current research, and future challenges // J. Integr. Plant Biol. 2017. V. 59, Iss. 6. P. 356–389.
Swaef T.D., De Schepper V., Vandegehuchte M.W., et al. Stem diameter variations as a versatile research tool in ecophysiology // Tree Physiol. 2015. V. 35 (10), Iss. 10. P. 1047–1061.
Pfautsch S., Aspinwall M.J, Drake J.E, et al. Traits and trade-offs in whole-tree hydraulic architecture along the vertical axis of Eucalyptus grandis // Ann. Bot. 2018. V. 121 (1). P. 129–141.
Zimmermann M.H. Xylem structure and the ascent of sap. Springer-Verlag, Berlin Heidelberg, 1983. 146 p.
Шавнин С.А., Овчинников И.С., Голиков Д.Ю., и др. Явление поворота ствола в процессе роста у древесных растений (на примере Pinus sylvestris L. и Picea obovata Ldb.) // Сиб. экол. журн. 2018. Т. 25, № 1. С. 89–97.
Колесников Б.П., Зубарева Р.С., Смолоногов Е.П. Лесорастительные условия и типы лесов Свердловской области: Практическое руководство. Свердловск: ИЭРиЖ, 1973. 176 с.
Свид. 2017610856 Российская Федерация. Свидетельство о государственной регистрации программ для ЭВМ. Программа решения задачи классификации на основе анализа наборов кластерных конфигурация и средств визуальной обработки данных / А.А. Монтиле, А.И. Монтиле, С.А. Шавнин; заявитель и правообладатель ФГБУН Ботанический сад УрО РАН (RU). – Заявка № 2016662912; заявл. 23.11.2016; опубл. 18.01.2017, Реестр программ для ЭВМ. – 1 с.
Интернет-ресурс метеорологической информации. 2019. URL: http://www.pogodaiklimat.ru
Cermak J., Kucera J. Scaling up transpiration data between trees, stands and watersheds // Silva Carelica. 1990. V. 15. P. 101–120.
Harris J.M. Spiral Grain and Wave Phenomena in Wood Formation. Springer-Verlag, 1989. 214 p.
Richter Ch. Wood characteristics: description, causes, prevention, impact on use and technological adaptation. Springer International Publishing Switzerland, 2015. 222 p.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни